
Proliferation and Differentiation of Intestinal Caco-2 Cells Are Maintained in Culture with Human Platelet Lysate Instead of Fetal Calf Serum



Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
- Panexin Basic Serum Replacement with Defined Components (P04-96090, PAN Biotech, Aidenbach, Germany); “basic”
- Panexin NTA Serum Substitute with Defined Components for Adherent Cells (P04-95750, PAN Biotech, Aidenbach, Germany); “NTA”
- PL solution research grade (PLS-100.01PL BioScience GmbH, Aachen, Germany); “hPL”
- PL solution-FD (fibrinogen derived) research grade (PLS-FD-500.01, PL BioScience GmbH, Aachen, Germany); “FD”
- PL solution-XF (fibrinogen derived, xenogen-free) research grade (PLS-XF-500.01, PL BioScience GmbH, Aachen, Germany); “XF”
2.2. Viability Assay
2.3. Gene Expression Analysis: Two-Step RT-qPCR
2.4. Protein Expression Analysis
2.4.1. Extraction of Total Protein
2.4.2. Preparation of Brush Border Membranes (BBM)
2.4.3. Preparation of Lipid Rafts (LR)
2.4.4. Western Blot Analysis
2.5. Statistics
3. Results
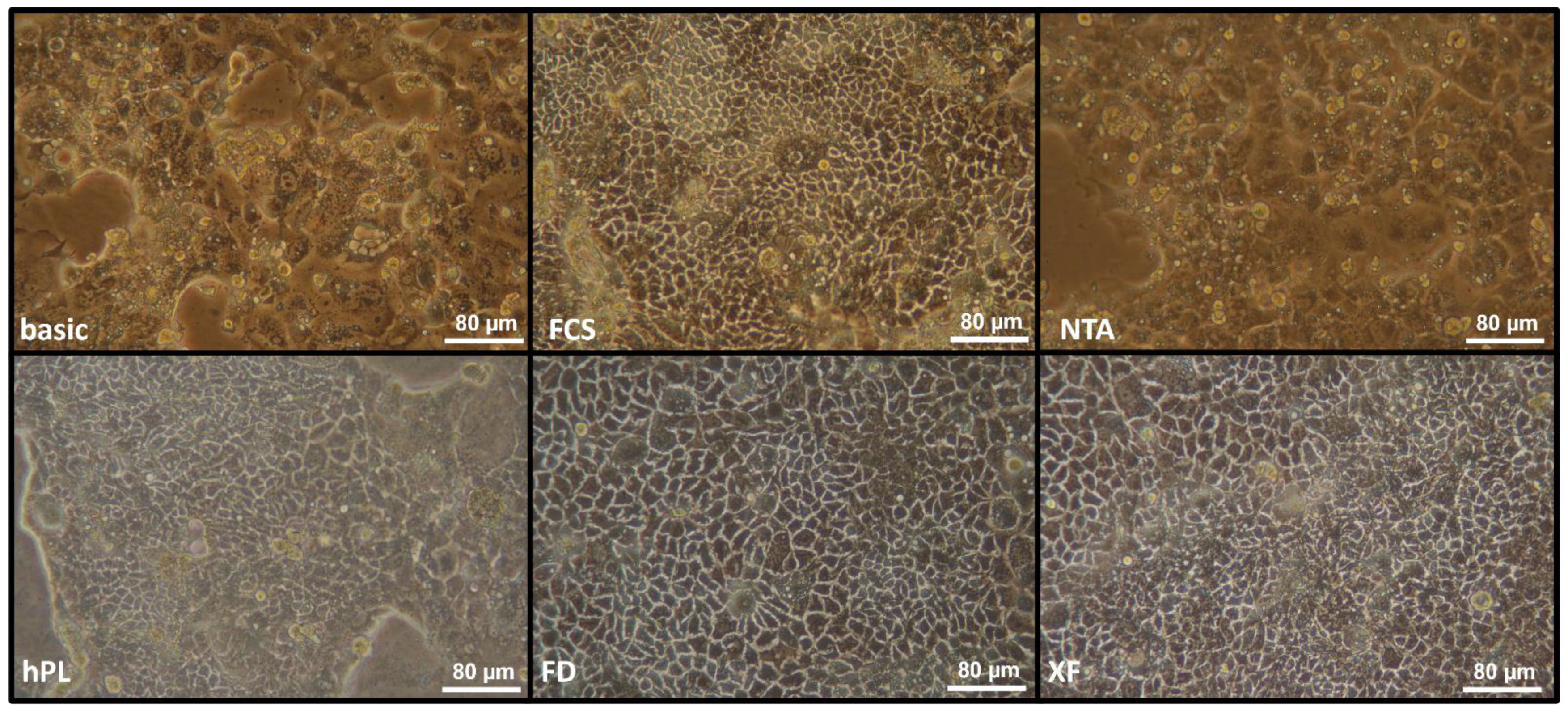
3.1. Morphology
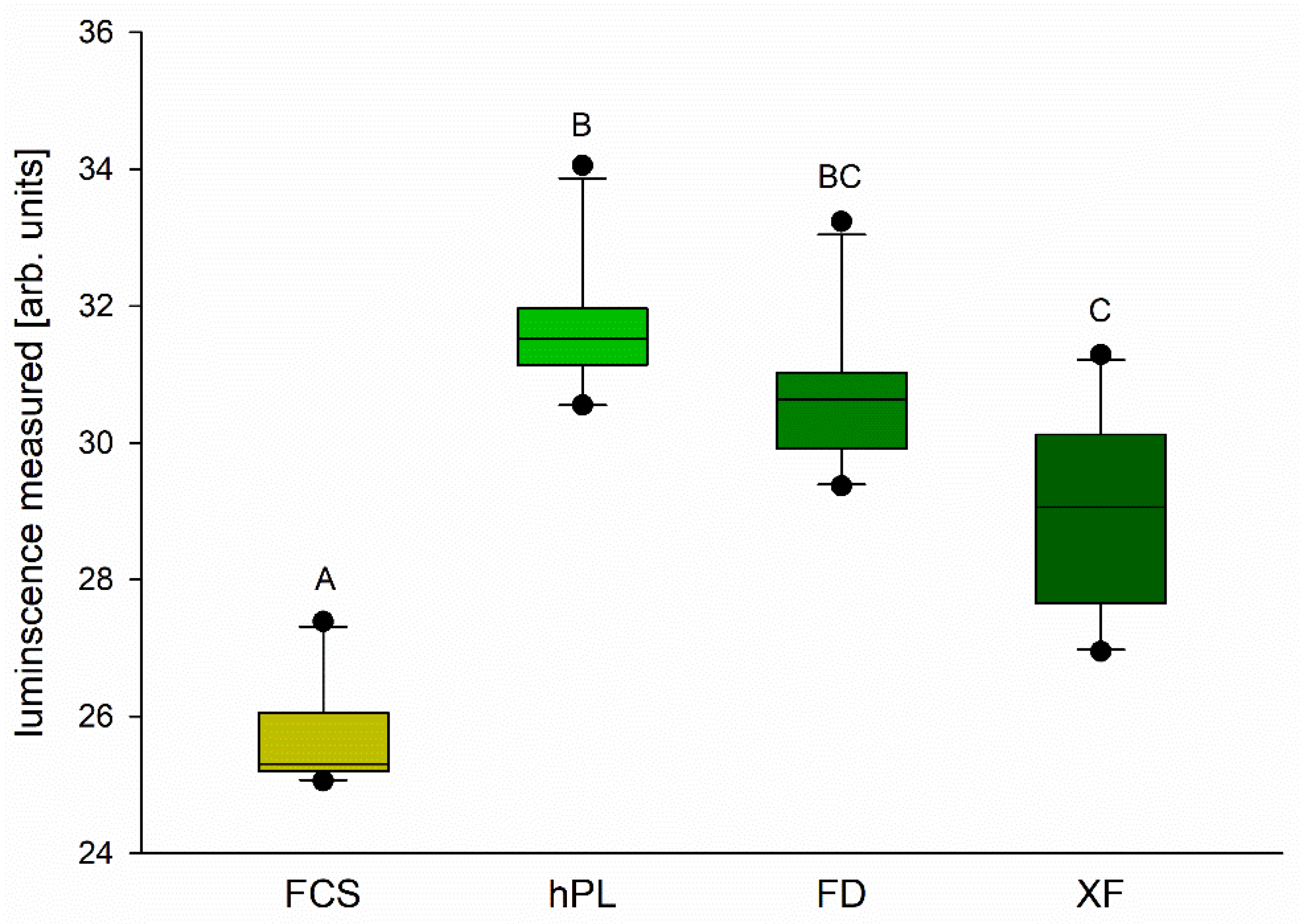
3.2. Cell Viability
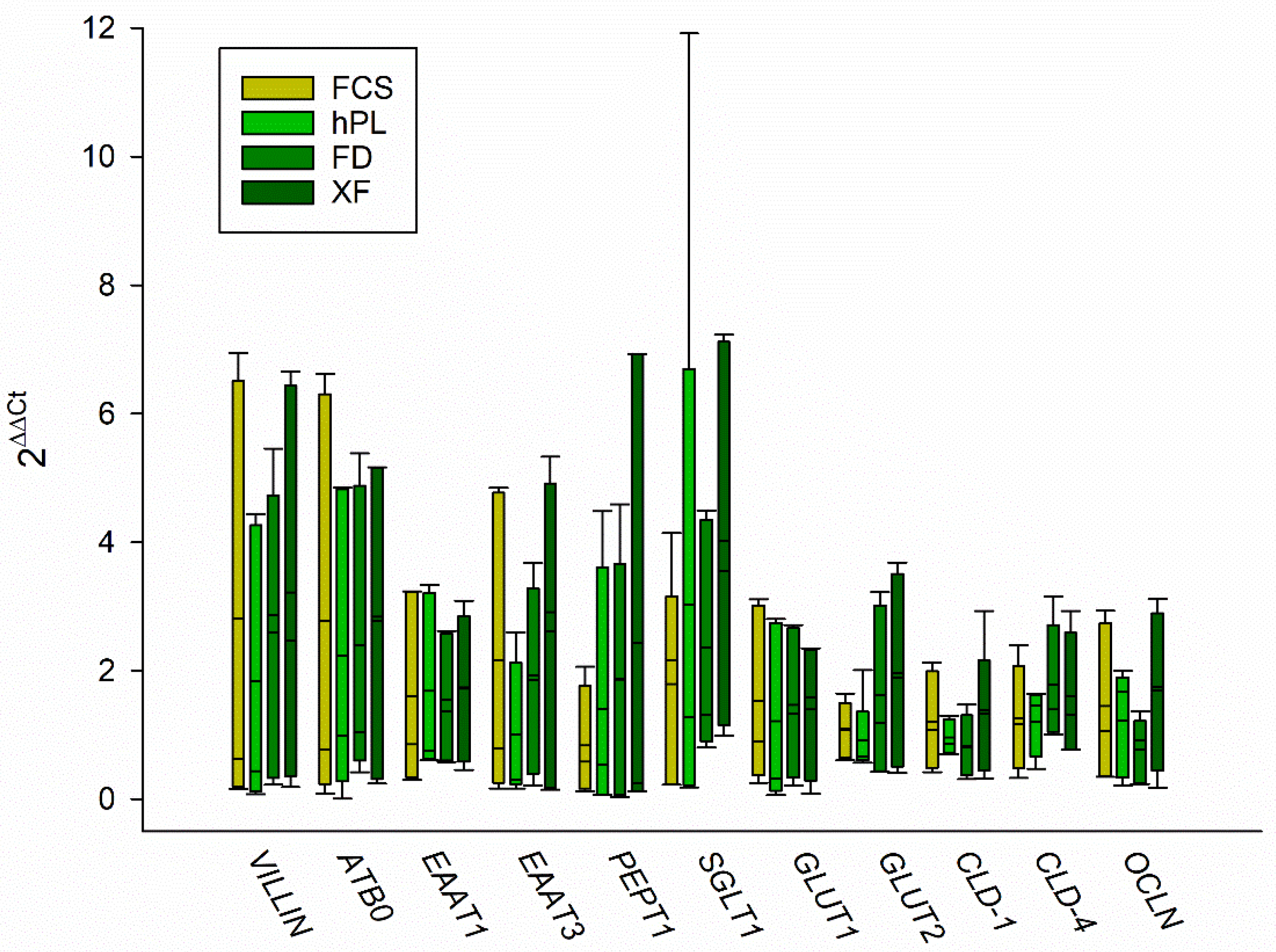
3.3. Gene Expression
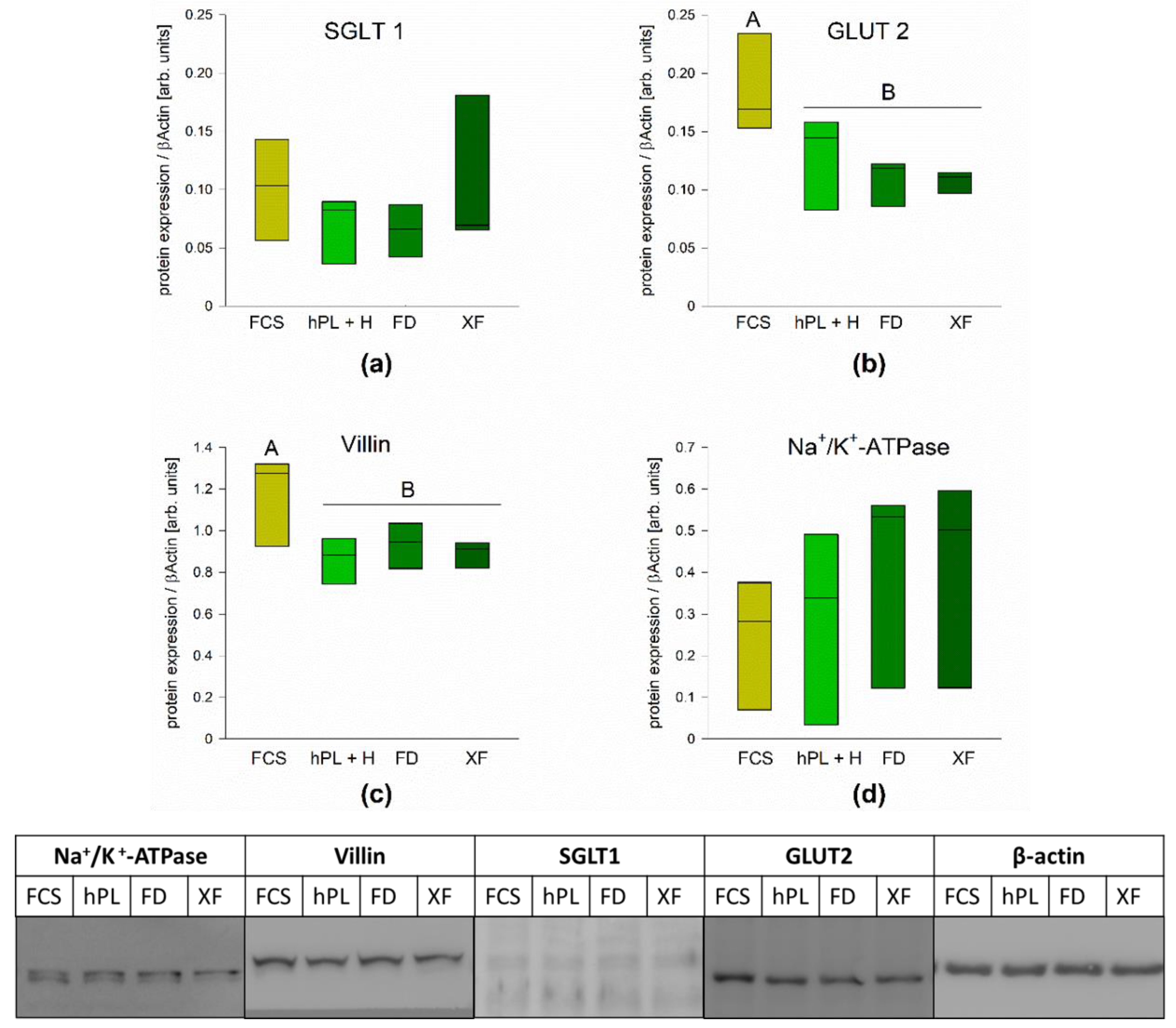
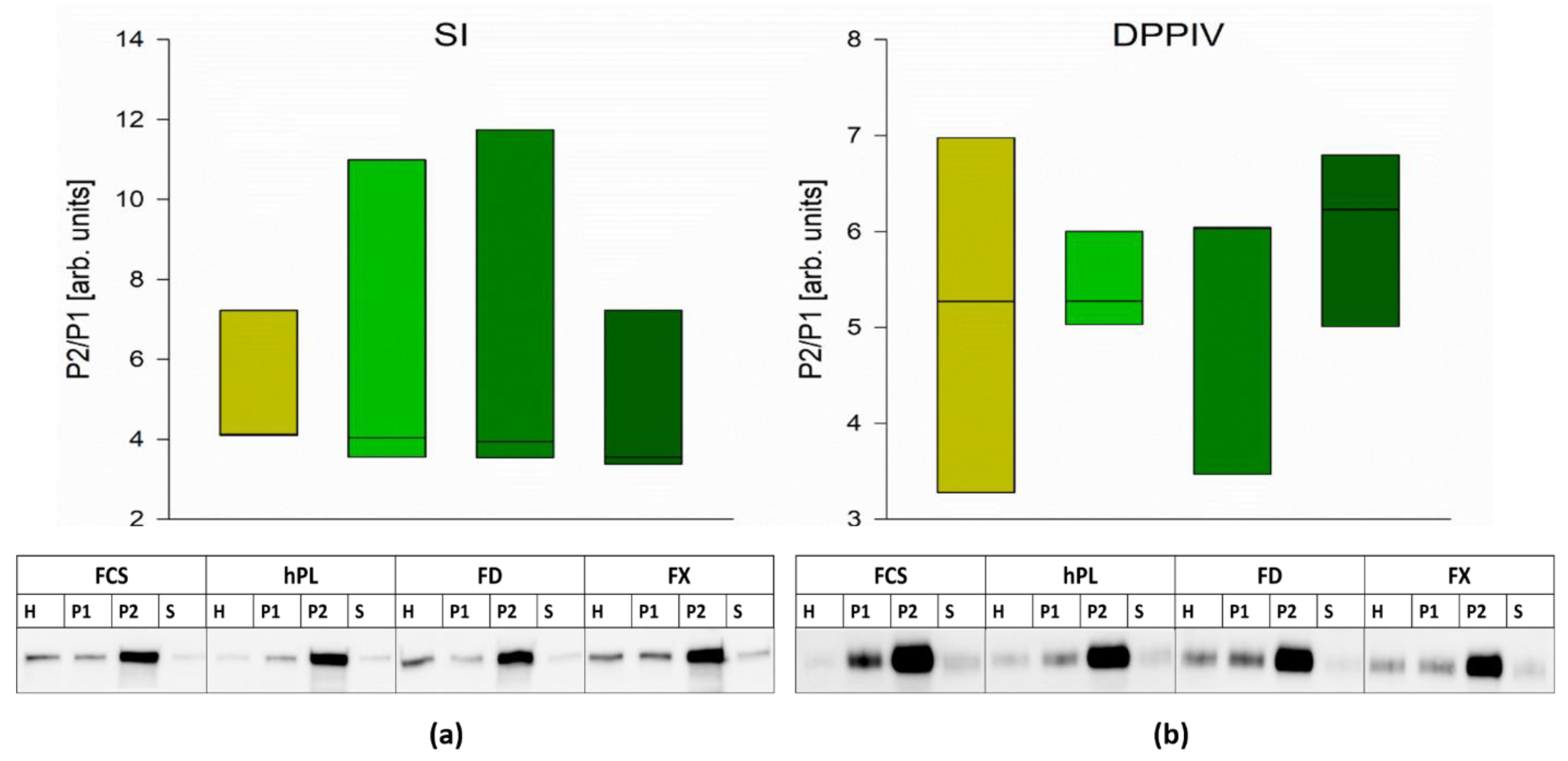
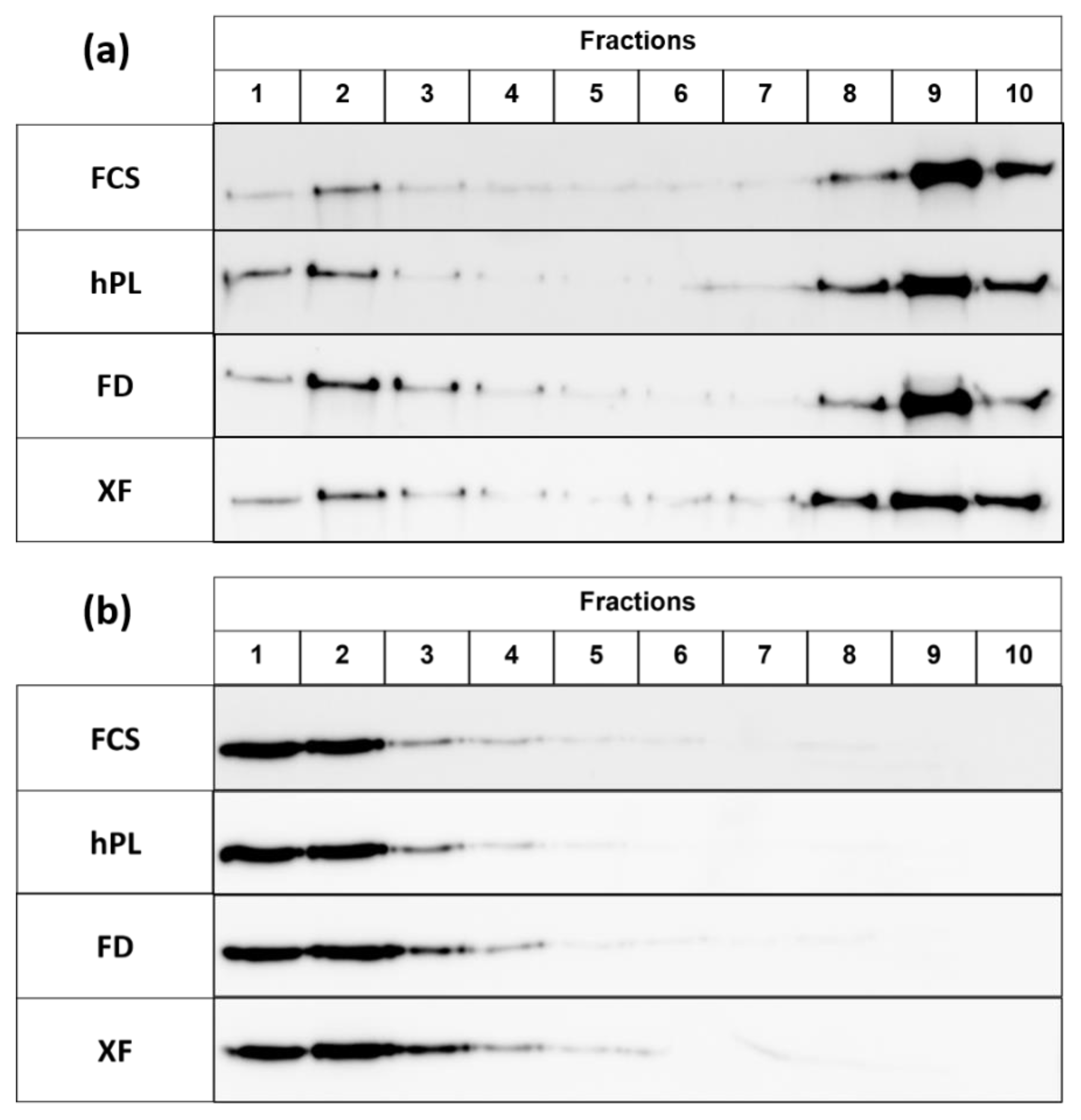
3.4. Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van der Valk, J.; Brunner, D.; De Smet, K.; Svenningsen, Å.F.; Honegger, P.; Knudsen, L.E.; Lindl, T.; Noraberg, J.; Price, A.; Scarino, M.L.; et al. Optimization of chemically defined cell culture media—Replacing fetal bovine serum in mammalian in vitro methods. Toxicol. Vitr. 2010, 24, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Jochems, C.E.A.; van der Valk, J.B.; Stafleu, F.R.; Baumans, V. The Use of Fetal Bovine Serum: Ethical or Scientific Problem? Altern. Lab. Anim. 2002, 30, 219–227. [Google Scholar] [CrossRef] [PubMed]
- van der Valk, J.; Mellor, D.; Brands, R.; Fischer, R.; Gruber, F.; Gstraunthaler, G.; Hellebrekers, L.; Hyllner, J.; Jonker, F.; Prieto, P.; et al. The humane collection of fetal bovine serum and possibilities for serum-free cell and tissue culture. Toxicol. Vitr. 2004, 18, 1–12. [Google Scholar] [CrossRef]
- Gstraunthaler, G. Alternatives to the use of fetal bovine serum: Serum-free cell culture. ALTEX 2003, 20, 275–281. [Google Scholar]
- Bauman, E.; Granja, P.; Barrias, C. Fetal bovine serum-free culture of endothelial progenitor cells-progress and challenges. J. Tissue Eng. Regen. Med. 2018, 12, 1567–1578. [Google Scholar] [CrossRef]
- Burnouf, T.; Strunk, D.; Koh, M.B.; Schallmoser, K. Human platelet lysate: Replacing fetal bovine serum as a gold standard for human cell propagation? Biomaterials 2016, 76, 371–387. [Google Scholar] [CrossRef]
- Mirabet, V.; Solves, P.; Miñana, M.D.; Encabo, A.; Carbonell-Uberos, F.; Blanquer, A.; Roig, R. Human platelet lysate enhances the proliferative activity of cultured human fibroblast-like cells from different tissues. Cell Tissue Bank. 2007, 9, 1–10. [Google Scholar] [CrossRef]
- Hemeda, H.; Giebel, B.; Wagner, W. Evaluation of human platelet lysate versus fetal bovine serum for culture of mesenchymal stromal cells. Cytotherapy 2014, 16, 170–180. [Google Scholar] [CrossRef]
- Johansson, L.; Klinth, J.; Holmqvist, O.; Ohlson, S. Platelet lysate: A replacement for fetal bovine serum in animal cell culture? Cytotechnology 2003, 42, 67–74. [Google Scholar] [CrossRef]
- Pons, M.; Nagel, G.; Zeyn, Y.; Beyer, M.; Laguna, T.; Brachetti, T.; Sellmer, A.; Mahboobi, S.; Conradi, R.; Butter, F.; et al. Human platelet lysate as validated replacement for animal serum to assess chemosensitivity_suppl. ALTEX 2018, 36, 277–288. [Google Scholar] [CrossRef]
- Rauch, C. Alternatives to the use of fetal bovine serum: Human platelet lysates as a serum substitute in cell culture media. ALTEX 2011, 28, 305–316. [Google Scholar] [CrossRef]
- Tylek, T.; Schilling, T.; Schlegelmilch, K.; Ries, M.; Rudert, M.; Jakob, F.; Groll, J. Platelet lysate outperforms FCS and human serum for co-culture of primary human macrophages and hMSCs. Sci. Rep. 2019, 9, 3533. [Google Scholar] [CrossRef]
- Fazzina, R.; Iudicone, P.; Mariotti, A.; Fioravanti, D.; Procoli, A.; Cicchetti, E.; Scambia, G.; Bonanno, G.; Pierelli, L. Culture of human cell lines by a pathogen-inactivated human platelet lysate. Cytotechnology 2015, 68, 1185–1195. [Google Scholar] [CrossRef][Green Version]
- Ferruzza, S. Serum-reduced and serum-free media for differentiation of Caco-2 cells. ALTEX 2013, 30, 159–168. [Google Scholar] [CrossRef][Green Version]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Van Breemen, R.B.; Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin. Drug Metab. Toxicol. 2005, 1, 175–185. [Google Scholar] [CrossRef]
- Shah, P.; Jogani, V.; Bagchi, T.; Misra, A. Role of Caco-2 Cell Monolayers in Prediction of Intestinal Drug Absorption. Biotechnol. Prog. 2006, 22, 186–198. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Schmitz, J.; Preiser, H.; Maestracci, D.; Ghosh, B.; Cerda, J.; Crane, R. Purification of the human intestinal brush border membrane. Biochim. Biophys. Acta (BBA) Biomembr. 1973, 323, 98–112. [Google Scholar] [CrossRef]
- Sterchi, E.E.; Woodley, J.F. Peptide hydrolases of the human small intestinal mucosa: Identification of six distinct enzymes in the brush border membrane. Clin. Chim. Acta 1980, 102, 57–65. [Google Scholar] [CrossRef]
- Shimada, Y.; Inomata, M.; Suzuki, H.; Hayashi, M.; Waheed, A.A.; Ohno-Iwashita, Y. Separation of a cholesterol-enriched microdomain involved in T-cell signal transduction. FEBS J. 2005, 272, 5454–5463. [Google Scholar] [CrossRef]
- Naim, H.Y.; E Sterchi, E.; Lentze, M.J. Biosynthesis of the human sucrase-isomaltase complex. Differential O-glycosylation of the sucrase subunit correlates with its position within the enzyme complex. J. Biol. Chem. 1988, 263, 7242–7253. [Google Scholar] [CrossRef]
- Hauri, H.P.; Sterchi, E.E.; Bienz, D.; Fransen, J.; Marxer, A. Expression and intracellular transport of microvillus membrane hydrolases in human intestinal epithelial cells. J. Cell Biol. 1985, 101, 838–851. [Google Scholar] [CrossRef]
- Van Der Valk, J. Fetal bovine serum (FBS): Past—Present—Future. ALTEX 2018, 35, 99–118. [Google Scholar] [CrossRef]
- Fernandez-Rebollo, E.; Mentrup, B.; Ebert, R.; Franzen, J.; Abagnale, G.; Sieben, T.; Ostrowska, A.; Hoffmann, P.; Roux, P.-F.; Rath, B.; et al. Human Platelet Lysate versus Fetal Calf Serum: These Supplements do not Select for Different Mesenchymal Stromal Cells. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Hemeda, H.; Kalz, J.; Walenda, G.; Lohmann, M.; Wagner, W. Heparin concentration is critical for cell culture with human platelet lysate. Cytotherapy 2013, 15, 1174–1181. [Google Scholar] [CrossRef]
- Thieme, D.; Reuland, L.; Lindl, T.; Kruse, F.; Fuchsluger, T. Optimized human platelet lysate as novel basis for a serum-, xeno-, and additive-free corneal endothelial cell and tissue culture. J. Tissue Eng. Regen. Med. 2017, 12, 557–564. [Google Scholar] [CrossRef]
- Nepal, N.; Arthur, S.; Sundaram, U. Unique Regulation of Na-K-ATPase during Growth and Maturation of Intestinal Epithelial Cells. Cells 2019, 8, 593. [Google Scholar] [CrossRef]
- Ranaldi, G.; Consalvo, R.; Sambuy, Y.; Scarino, M.L. Permeability characteristics of parental and clonal human intestinal Caco-2 cell lines differentiated in serum-supplemented and serum-free media. Toxicol. Vitr. 2003, 17, 761–767. [Google Scholar] [CrossRef]
- Zweibaum, A.; Triadou, N.; Kedinger, M.; Augeron, C.; Robine-Léon, S.; Pinto, M.; Rousset, M.; Haffen, K. Sucrase-isomaltase: A marker of foetal and malignant epithelial cells of the human colon. Int. J. Cancer 1983, 32, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, G.; Heine, M.; Rohwedder, A.; Naim, H.Y. Impact of glycosylation and detergent-resistant membranes on the function of intestinal sucrase-isomaltase. Biol. Chem. 2009, 390, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Toutounji, M.; Wanes, D.; El-Harakeh, M.; El-Sabban, M.; Rizk, S.; Naim, H.Y. Dextran Sodium Sulfate-Induced Impairment of Protein Trafficking and Alterations in Membrane Composition in Intestinal Caco-2 Cell Line. Int. J. Mol. Sci. 2020, 21, 2726. [Google Scholar] [CrossRef] [PubMed]
- Alfalah, M.; Jacob, R.; Preuss, U.; Zimmer, K.-P.; Naim, H.; Naim, H.Y. O-linked glycans mediate apical sorting of human intestinal sucrase-isomaltase through association with lipid rafts. Curr. Biol. 1999, 9, 593–S2. [Google Scholar] [CrossRef]
- Hashimoto, K.; Shimizu, M. Epithelial properties of human intestinal Caco-2 cells cultured in a serum-free medium. Cytotechnology 1993, 13, 175–184. [Google Scholar] [CrossRef]
- Warrier, A.; Gunosewoyo, H.; Crowe, A. Efflux transporters and tight junction expression changes in human gastrointestinal cell lines cultured in defined medium vs serum supplemented medium. Life Sci. 2018, 207, 138–144. [Google Scholar] [CrossRef]
- Aldén, A.; Gonzalez, L.; Persson, A.; Christensson, K.; Holmqvist, O.; Ohlson, S. Porcine platelet lysate as a supplement for animal cell culture. Cytotechnology 2007, 55, 3–8. [Google Scholar] [CrossRef][Green Version]
- Hagen, A.; Lehmann, H.; Aurich, S.; Bauer, N.; Melzer, M.; Moellerberndt, J.; Patané, V.; Schnabel, C.L.; Burk, J. Scalable Production of Equine Platelet Lysate for Multipotent Mesenchymal Stromal Cell Culture. Front. Bioeng. Biotechnol. 2021, 8, 613621. [Google Scholar] [CrossRef]
- Abu-Ameerh, M.A.; Jafar, H.; Hasan, M.H.; Al Bdour, M.; Msallam, M.; Ababneh, O.H.; Alhattab, D.M.; Al-Kurdi, B.; Awidi, A.; Awidi, A.S. Platelet lysate promotes re-epithelialization of persistent epithelial defects: A pilot study. Int. Ophthalmol. 2019, 39, 1483–1490. [Google Scholar] [CrossRef]
- Henschler, R.; Gabriel, C.; Schallmoser, K.; Burnouf, T.; Koh, M.B. Human platelet lysate current standards and future developments. Transfusion 2019, 59, 1407–1413. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Bank Accession Number | Primer Sequence (5′–3′) | Annealing Temperature [°C] | Amplicon Length [bp] |
|---|---|---|---|---|
| HPRT1 | NM_000194.3 | F: ATGGACAGGACTGAACGTCTT R: TGTAATCCAGCAGGTCAGCA | 57 | 118 |
| PPIA | NM_021130.5 | F: GCCGAGGAAAACCGTGTACT R: CTGCAAACAGCTCAAAGGAGAC | 59 | 106 |
| SGLT1 | NM_000343.4 | F: AAGACCACCGCGGTCAC R: AAACATAGCCCACAGTCCGA | 57 | 119 |
| GLUT1 | NM_006516.4 | F: GAACTCTTCAGCCAGGGTCC | 60 | 113 |
| R: ACCACACAGTTGCTCCACAT | ||||
| GLUT2 | NM_000340.2 | F: CAATGCACCTCAACAGGTAATAA R: AGATTGTGGGCAGTTCATCTGT | 57 | 119 |
| PEPT1 | NG_017032.1 | F: GGTCCCACTGCTGCTTTTTG | 60 | 117 |
| R: GCAGAGCAAGGGGTTGTACT | ||||
| EAAT1 | NM_001166696.3 | F: CCCCTTACAAAATCAGAAAAGTTGT | 60 | 101 |
| R: GGAGGGTCTCTTCTTTGCACT | ||||
| EAAT3 | NM_004170.6 | F: GGTGGTGCTAGGCATTACCA | 60 | 118 |
| R: TGATGAGTTTCAGCATCCGCA | ||||
| ATB0 | NM_005628.3 | F: GTGTCCTCACTCTGGCCATC | 60 | 98 |
| R: TACAGGACCGGTCGACTAGC | ||||
| VILLIN | NM_007127.3 | F: ACCGAGGGGATGTTTTCCTC R: GTCATGCCCCTGAGTCTCTC | 60 | 93 |
| CLD 1 | NM_021101.5 | F: CCAGTCAATGCCAGGTACGA | 60 | 93 |
| R: ACAGCAAAGTAGGGCACCTC | ||||
| CLD 4 | NM_001305.5 | F: TCTCCTCTGTTCCGGGTAGG | 60 | 90 |
| R: CAAGGCCTCAGCCATACTCC | ||||
| OCLN | NM_002538.4 | F: GGTCTAGGACGCAGCAGATTG | 60 | 112 |
| R: TGGACTTTCAAGAGGCCTGG |
| Target | Primary Antibody | Manufacturer/Catalog Number | Dilution | Secondary Antibody | Manufacturer/ Catalog Number | Dilution |
|---|---|---|---|---|---|---|
| β-ACTIN | Mouse-anti-β-Actin | Santa Cruz Biotechnology/sc-47778 | 1:1000 | Goat-anti-mouse HRP | Invitrogen/A16072 | 1:5000 |
| DPPIV | Mouse-anti-Dipeptidyl peptidase (HBB 3/775/42) | Provided by Dres. E.E. Sterchi and H.-P. Hauri. University of Bern and University of Basel, Switzerland [23,24] | 1:5000 | Goat-anti-mouse HRP | Invitrogen/A16072 | 1:5000 |
| GLUT2 | Rabbit-anti-GLLUT2 | Thermo Fisher Scientific/PA5-77459 | 1:200 | Donkey-anti-rabbit HRP | Santa Cruz Biotechnology/sc-2077 | 1:10,000 |
| Na+/K+-ATPase | Mouse-anti-Na+/K+-ATPase | Enzo Life Sciences AG 804-082 | 1:100 | Goat-anti-mouse HRP | Invitrogen/A16072 | 1:5000 |
| SGLT1 | Rabbit-anti-SGLT1 | Antibodies online/ABIN364451 | 1:1000 | Donkey-anti-rabbit HRP | Santa Cruz Biotechnology/sc-2077 | 1:10,000 |
| SI | Mouse-anti-sucrase–isomaltase (HBB3/705/60) | Provided by Drs. E.E. Sterchi and H.-P. Hauri. University of Bern and University of Basel, Switzerland [23,24] | 1:5000 | Goat-anti-mouse HRP | Invitrogen/A16072 | 1:5000 |
| VILLIN | Rabbit-anti-Villin | Thermo Fisher Scientific/PA5-78222 | 1:500 | Donkey-anti-rabbit HRP | Santa Cruz Biotechnology/sc-2077 | 1:10,000 |
| FLOTILLIN-2 | Mouse-anti-flotillin-2 B-6 | Santa Cruz Biotechnology/sc-28320 | 1:5000 | Goat-anti-mouse HRP | Santa Cruz Biotechnology, Inc., Texas, US | 1:5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanes, D.; Naim, H.Y.; Dengler, F. Proliferation and Differentiation of Intestinal Caco-2 Cells Are Maintained in Culture with Human Platelet Lysate Instead of Fetal Calf Serum. Cells 2021, 10, 3038. https://doi.org/10.3390/cells10113038
Wanes D, Naim HY, Dengler F. Proliferation and Differentiation of Intestinal Caco-2 Cells Are Maintained in Culture with Human Platelet Lysate Instead of Fetal Calf Serum. Cells. 2021; 10(11):3038. https://doi.org/10.3390/cells10113038
Chicago/Turabian StyleWanes, Dalanda, Hassan Y. Naim, and Franziska Dengler. 2021. "Proliferation and Differentiation of Intestinal Caco-2 Cells Are Maintained in Culture with Human Platelet Lysate Instead of Fetal Calf Serum" Cells 10, no. 11: 3038. https://doi.org/10.3390/cells10113038
APA StyleWanes, D., Naim, H. Y., & Dengler, F. (2021). Proliferation and Differentiation of Intestinal Caco-2 Cells Are Maintained in Culture with Human Platelet Lysate Instead of Fetal Calf Serum. Cells, 10(11), 3038. https://doi.org/10.3390/cells10113038