
Harnessing CD8+CD28− Regulatory T Cells as a Tool to Treat Autoimmune Disease


{kind=link}
Abstract
:1. Introduction
2. Origin of CD8+ Tregs
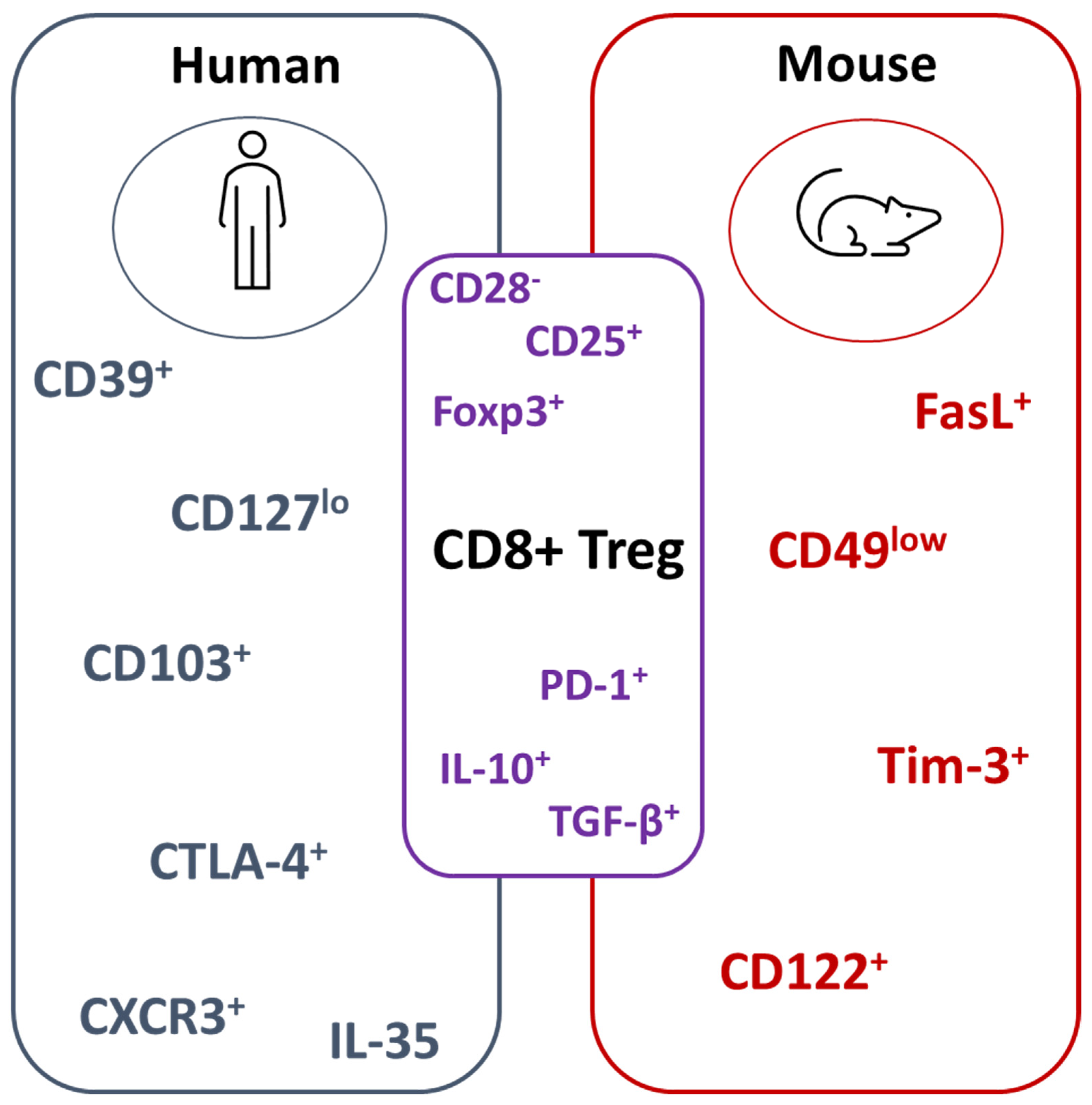
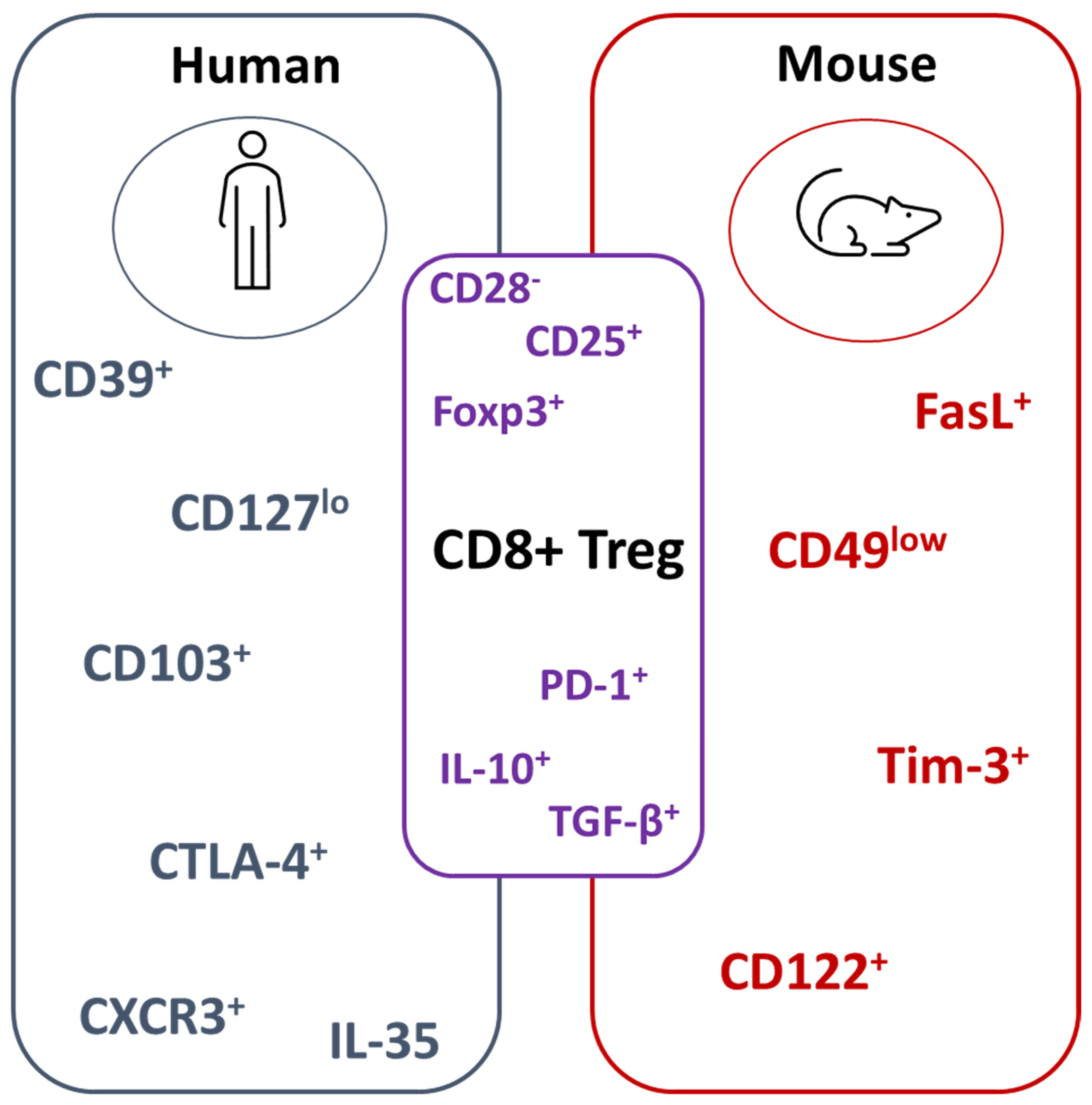
3. CD8+ Treg Populations in Mice and Humans
4. Mechanisms of Suppression Mediated by CD8+ Tregs
5. CD8+CD28− Tregs
6. CD8+CD28− Tregs in Autoimmunity
6.1. Multiple Sclerosis
6.2. Rheumatoid Arthritis (RA)
6.3. Systemic Lupus Erythematosus (SLE)
6.4. Diabetes
6.5. Systemic Sclerosis (SSc)
7. CD8+CD28− Tregs in Gastrointestinal Disease
8. Viral Infections
9. Transplantation
10. Cancer
11. Adoptive Cell Therapy
12. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Cantor, H.; Shen, F.W.; Boyse, E.A. Separation of helper T cells from suppressor T cells expressing different Ly components. II. Activation by antigen: After immunization, antigen-specific suppressor and helper activities are mediated by distinct T-cell subclasses. J. Exp. Med. 1976, 143, 1391. [Google Scholar] [CrossRef] [Green Version]
- Jandinski, J.; Cantor, H.; Tadakuma, T.; Peavy, D.L.; Pierce, C.W. Separation of helper T cells from suppressor T cells expressing different Ly components. I. Polyclonal activation: Suppressor and helper activities are inherent properties of distinct T-cell subclasses. J. Exp. Med. 1976, 143, 1382–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Khattri, R.; Cox, T.; Yasayko, S.A.; Ramsdell, F. An essential role for Scurfin in CD4+CD25+ T regulatory cells. Nat. Immunol. 2003, 4, 337–342. [Google Scholar] [CrossRef]
- Bennett, C.L.; Christie, J.; Ramsdell, F.; Brunkow, M.E.; Ferguson, P.J.; Whitesell, L.; Kelly, T.E.; Saulsbury, F.T.; Chance, P.F.; Ochs, H.D. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat. Genet. 2001, 27, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Powell, B.R.; Buist, N.R.; Stenzel, P. An X-linked syndrome of diarrhea, polyendocrinopathy, and fatal infection in infancy. J. Pediatr. 1982, 100, 731–737. [Google Scholar] [CrossRef]
- Mahic, M.; Henjum, K.; Yaqub, S.; Bjornbeth, B.A.; Torgersen, K.M.; Tasken, K.; Aandahl, E.M. Generation of highly suppressive adaptive CD8(+)CD25(+)FOXP3(+) regulatory T cells by continuous antigen stimulation. Eur. J. Immunol. 2008, 38, 640–646. [Google Scholar] [CrossRef]
- Filaci, G.; Fravega, M.; Negrini, S.; Procopio, F.; Fenoglio, D.; Rizzi, M.; Brenci, S.; Contini, P.; Olive, D.; Ghio, M.; et al. Nonantigen specific CD8+ T suppressor lymphocytes originate from CD8+CD28− T cells and inhibit both T-cell proliferation and CTL function. Hum. Immunol. 2004, 65, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Poggi, A.; Zocchi, M.R. Role of bone marrow stromal cells in the generation of human CD8+ regulatory T cells. Hum. Immunol. 2008, 69, 755–759. [Google Scholar] [CrossRef]
- Niederlova, V.; Tsyklauri, O.; Chadimova, T.; Stepanek, O. CD8(+) Tregs revisited: A heterogeneous population with different phenotypes and properties. Eur. J. Immunol. 2021, 51, 512–530. [Google Scholar] [CrossRef]
- Shao, L.; Jacobs, A.R.; Johnson, V.V.; Mayer, L. Activation of CD8+ regulatory T cells by human placental trophoblasts. J. Immunol. 2005, 174, 7539–7547. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Futagami, Y.; Horie, S.; Mochizuki, M. Transforming growth factor beta-producing Foxp3(+)CD8(+)CD25(+) T cells induced by iris pigment epithelial cells display regulatory phenotype and acquire regulatory functions. Exp. Eye Res. 2007, 85, 626–636. [Google Scholar] [CrossRef]
- Fagnoni, F.F.; Vescovini, R.; Mazzola, M.; Bologna, G.; Nigro, E.; Lavagetto, G.; Franceschi, C.; Passeri, M.; Sansoni, P. Expansion of cytotoxic CD8+ CD28− T cells in healthy ageing people, including centenarians. Immunology 1996, 88, 501–507. [Google Scholar] [CrossRef]
- Parish, S.T.; Wu, J.E.; Effros, R.B. Modulation of T lymphocyte replicative senescence via TNF-{alpha} inhibition: Role of caspase-3. J. Immunol. 2009, 182, 4237–4243. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.; Wan, N.; Zhang, S.; Moore, Y.; Wan, F.; Dai, Z. Cutting edge: Programmed death-1 defines CD8+CD122+ T cells as regulatory versus memory T cells. J. Immunol. 2010, 185, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Rifa’i, M.; Shi, Z.; Zhang, S.Y.; Lee, Y.H.; Shiku, H.; Isobe, K.; Suzuki, H. CD8+CD122+ regulatory T cells recognize activated T cells via conventional MHC class I-alphabetaTCR interaction and become IL-10-producing active regulatory cells. Int. Immunol. 2008, 20, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Izawa, A.; Yamaura, K.; Albin, M.J.; Jurewicz, M.; Tanaka, K.; Clarkson, M.R.; Ueno, T.; Habicht, A.; Freeman, G.J.; Yagita, H.; et al. A novel alloantigen-specific CD8+PD1+ regulatory T cell induced by ICOS-B7h blockade in vivo. J. Immunol. 2007, 179, 786–796. [Google Scholar] [CrossRef]
- Villarreal, D.O.; Allegrezza, M.J.; Smith, M.A.; Chin, D.; Luistro, L.L.; Snyder, L.A. Targeting of CD122 enhances antitumor immunity by altering the tumor immune environment. Oncotarget 2017, 8, 109151–109160. [Google Scholar] [CrossRef] [Green Version]
- Motegi, A.; Kinoshita, M.; Inatsu, A.; Habu, Y.; Saitoh, D.; Seki, S. IL-15-induced CD8+CD122+ T cells increase antibacterial and anti-tumor immune responses: Implications for immune function in aged mice. J. Leukoc. Biol. 2008, 84, 1047–1056. [Google Scholar] [CrossRef]
- Suzuki, H.; Shi, Z.; Okuno, Y.; Isobe, K. Are CD8+CD122+ cells regulatory T cells or memory T cells? Hum. Immunol. 2008, 69, 751–754. [Google Scholar] [CrossRef]
- Ellis, S.D.; McGovern, J.L.; van Maurik, A.; Howe, D.; Ehrenstein, M.R.; Notley, C.A. Induced CD8+Foxp3+ Treg cells in rheumatoid arthritis are modulated by p38 phosphorylation and monocytes expressing membrane tumor necrosis factor alpha and CD86. Arthritis Rheumatol. 2014, 66, 2694–2705. [Google Scholar] [CrossRef]
- Zhang, L.; Bertucci, A.M.; Ramsey-Goldman, R.; Burt, R.K.; Datta, S.K. Regulatory T cell (Treg) subsets return in patients with refractory lupus following stem cell transplantation, and TGF-beta-producing CD8+ Treg cells are associated with immunological remission of lupus. J. Immunol. 2009, 183, 6346–6358. [Google Scholar] [CrossRef] [Green Version]
- Popescu, I.; Macedo, C.; Abu-Elmagd, K.; Shapiro, R.; Hua, Y.; Thomson, A.W.; Morelli, A.E.; Storkus, W.J.; Metes, D. EBV-specific CD8+ T cell reactivation in transplant patients results in expansion of CD8+ type-1 regulatory T cells. Am. J. Transpl. 2007, 7, 1215–1223. [Google Scholar] [CrossRef]
- Kiniwa, Y.; Miyahara, Y.; Wang, H.Y.; Peng, W.; Peng, G.; Wheeler, T.M.; Thompson, T.C.; Old, L.J.; Wang, R.F. CD8+ Foxp3+ regulatory T cells mediate immunosuppression in prostate cancer. Clin. Cancer Res. 2007, 13, 6947–6958. [Google Scholar] [CrossRef] [Green Version]
- Avivi, I.; Stroopinsky, D.; Rowe, J.M.; Katz, T. A subset of CD8+ T cells acquiring selective suppressive properties may play a role in GvHD management. Transpl. Immunol. 2013, 28, 57–61. [Google Scholar] [CrossRef]
- Sharabi, A.; Mozes, E. The suppression of murine lupus by a tolerogenic peptide involves foxp3-expressing CD8 cells that are required for the optimal induction and function of foxp3-expressing CD4 cells. J. Immunol. 2008, 181, 3243–3251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, R.J.; Lineburg, K.E.; Kuns, R.D.; Wilson, Y.A.; Raffelt, N.C.; Olver, S.D.; Varelias, A.; Alexander, K.A.; Teal, B.E.; Sparwasser, T.; et al. Identification and expansion of highly suppressive CD8(+)FoxP3(+) regulatory T cells after experimental allogeneic bone marrow transplantation. Blood 2012, 119, 5898–5908. [Google Scholar] [CrossRef] [Green Version]
- Zimmerer, J.M.; Ringwald, B.A.; Elzein, S.M.; Avila, C.L.; Warren, R.T.; Abdel-Rasoul, M.; Bumgardner, G.L. Antibody-suppressor CD8+ T Cells Require CXCR5. Transplantation 2019, 103, 1809–1820. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.X.; Wu, G.; Zou, Y.; Feng, Y.L.; Liu, H.B.; Feng, J.S.; Chi, H.G.; Lv, R.X.; Zheng, X.B. Balance of CD8+ CD28+/CD8+ CD28− T lymphocytes is vital for patients with ulcerative colitis. Dig. Dis. Sci. 2013, 58, 88–96. [Google Scholar] [CrossRef]
- Tulunay, A.; Yavuz, S.; Direskeneli, H.; Eksioglu-Demiralp, E. CD8+CD28−, suppressive T cells in systemic lupus erythematosus. Lupus 2008, 17, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Mikulkova, Z.; Praksova, P.; Stourac, P.; Bednarik, J.; Strajtova, L.; Pacasova, R.; Belobradkova, J.; Dite, P.; Michalek, J. Numerical defects in CD8+CD28− T-suppressor lymphocyte population in patients with type 1 diabetes mellitus and multiple sclerosis. Cell Immunol. 2010, 262, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Ceeraz, S.; Hall, C.; Choy, E.H.; Spencer, J.; Corrigall, V.M. Defective CD8+CD28+ regulatory T cell suppressor function in rheumatoid arthritis is restored by tumour necrosis factor inhibitor therapy. Clin. Exp. Immunol. 2013, 174, 18–26. [Google Scholar] [CrossRef]
- Jiang, H.; Chess, L. The specific regulation of immune responses by CD8+ T cells restricted by the MHC class Ib molecule, Qa-1. Annu. Rev. Immunol. 2000, 18, 185–216. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Verbinnen, B.; Tang, X.; Lu, L.; Cantor, H. Inhibition of follicular T-helper cells by CD8(+) regulatory T cells is essential for self tolerance. Nature 2010, 467, 328–332. [Google Scholar] [CrossRef]
- Clement, M.; Guedj, K.; Andreata, F.; Morvan, M.; Bey, L.; Khallou-Laschet, J.; Gaston, A.T.; Delbosc, S.; Alsac, J.M.; Bruneval, P.; et al. Control of the T follicular helper-germinal center B-cell axis by CD8(+) regulatory T cells limits atherosclerosis and tertiary lymphoid organ development. Circulation 2015, 131, 560–570. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Y.; Zeng, Q.; Zeng, Y.Q.; Liang, C.L.; Qiu, F.; Nie, H.; Dai, Z. Suppression of allograft rejection by CD8+CD122+PD-1+ Tregs is dictated by their Fas ligand-initiated killing of effector T cells versus Fas-mediated own apoptosis. Oncotarget 2017, 8, 24187–24195. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, D.A.; Pan, S.; Ou, J.N.; Wang, J.; Chen, M.; Gray, J.D.; Zheng, S.G. Therapeutic polyclonal human CD8+ CD25+ Fox3+ TNFR2+ PD-L1+ regulatory cells induced ex-vivo. Clin. Immunol. 2013, 149, 450–463. [Google Scholar] [CrossRef] [Green Version]
- Taghavie-Moghadam, P.L.; Waseem, T.C.; Hattler, J.; Glenn, L.M.; Dobrian, A.D.; Kaplan, M.H.; Yang, Y.; Nurieva, R.; Nadler, J.L.; Galkina, E.V. STAT4 Regulates the CD8(+) Regulatory T Cell/T Follicular Helper Cell Axis and Promotes Atherogenesis in Insulin-Resistant Ldlr(-/-) Mice. J. Immunol. 2017, 199, 3453–3465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yu, M.; Zheng, Y.; Fu, G.; Xin, G.; Zhu, W.; Luo, L.; Burns, R.; Li, Q.Z.; Dent, A.L.; et al. CXCR5(+)PD-1(+) follicular helper CD8 T cells control B cell tolerance. Nat. Commun. 2019, 10, 4415. [Google Scholar] [CrossRef] [Green Version]
- Simone, R.; Zicca, A.; Saverino, D. The frequency of regulatory CD3+CD8+CD28− CD25+ T lymphocytes in human peripheral blood increases with age. J. Leukoc. Biol. 2008, 84, 1454–1461. [Google Scholar] [CrossRef] [Green Version]
- Filaci, G.; Fenoglio, D.; Fravega, M.; Ansaldo, G.; Borgonovo, G.; Traverso, P.; Villaggio, B.; Ferrera, A.; Kunkl, A.; Rizzi, M.; et al. CD8+ CD28− T regulatory lymphocytes inhibiting T cell proliferative and cytotoxic functions infiltrate human cancers. J. Immunol. 2007, 179, 4323–4334. [Google Scholar] [CrossRef] [Green Version]
- Colovai, A.I.; Mirza, M.; Vlad, G.; Wang, S.; Ho, E.; Cortesini, R.; Suciu-Foca, N. Regulatory CD8+CD28− T cells in heart transplant recipients. Hum. Immunol. 2003, 64, 31–37. [Google Scholar] [CrossRef]
- Baeten, D.; Louis, S.; Braud, C.; Braudeau, C.; Ballet, C.; Moizant, F.; Pallier, A.; Giral, M.; Brouard, S.; Soulillou, J.P. Phenotypically and functionally distinct CD8+ lymphocyte populations in long-term drug-free tolerance and chronic rejection in human kidney graft recipients. J. Am. Soc. Nephrol. 2006, 17, 294–304. [Google Scholar] [CrossRef] [Green Version]
- Ciubotariu, R.; Vasilescu, R.; Ho, E.; Cinti, P.; Cancedda, C.; Poli, L.; Late, M.; Liu, Z.; Berloco, P.; Cortesini, R.; et al. Detection of T suppressor cells in patients with organ allografts. Hum. Immunol. 2001, 62, 15–20. [Google Scholar] [CrossRef]
- Allez, M.; Brimnes, J.; Dotan, I.; Mayer, L. Expansion of CD8+ T cells with regulatory function after interaction with intestinal epithelial cells. Gastroenterology 2002, 123, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Ciubotariu, R.; Colovai, A.I.; Pennesi, G.; Liu, Z.; Smith, D.; Berlocco, P.; Cortesini, R.; Suciu-Foca, N. Specific suppression of human CD4+ Th cell responses to pig MHC antigens by CD8+CD28− regulatory T cells. J. Immunol. 1998, 161, 5193–5202. [Google Scholar]
- Jiang, S.; Tugulea, S.; Pennesi, G.; Liu, Z.; Mulder, A.; Lederman, S.; Harris, P.; Cortesini, R.; Suciu-Foca, N. Induction of MHC-class I restricted human suppressor T cells by peptide priming in vitro. Hum. Immunol. 1998, 59, 690–699. [Google Scholar] [CrossRef]
- Scotto, L.; Naiyer, A.J.; Galluzzo, S.; Rossi, P.; Manavalan, J.S.; Kim-Schulze, S.; Fang, J.; Favera, R.D.; Cortesini, R.; Suciu-Foca, N. Overlap between molecular markers expressed by naturally occurring CD4+CD25+ regulatory T cells and antigen specific CD4+CD25+ and CD8+CD28− T suppressor cells. Hum. Immunol. 2004, 65, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Z.; Jiang, S.; Cortesini, R.; Lederman, S.; Suciu-Foca, N. T suppressor lymphocytes inhibit NF-kappa B-mediated transcription of CD86 gene in APC. J. Immunol. 1999, 163, 6386–6392. [Google Scholar]
- Manavalan, J.S.; Kim-Schulze, S.; Scotto, L.; Naiyer, A.J.; Vlad, G.; Colombo, P.C.; Marboe, C.; Mancini, D.; Cortesini, R.; Suciu-Foca, N. Alloantigen specific CD8+CD28− Foxp3+ T suppressor cells induce ILT3+ ILT4+ tolerogenic endothelial cells, inhibiting alloreactivity. Int. Immunol. 2004, 16, 1055–1068. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zitzner, J.R.; Houlihan, J.; Herrera, N.; Xu, L.; Miller, J.; Mathew, J.M.; Tambur, A.R.; Luo, X. Common gamma chain cytokines promote rapid in vitro expansion of allo-specific human CD8+ suppressor T cells. PLoS ONE 2011, 6, e28948. [Google Scholar] [CrossRef]
- Najafian, N.; Chitnis, T.; Salama, A.D.; Zhu, B.; Benou, C.; Yuan, X.; Clarkson, M.R.; Sayegh, M.H.; Khoury, S.J. Regulatory functions of CD8+CD28− T cells in an autoimmune disease model. J. Clin. Invest. 2003, 112, 1037–1048. [Google Scholar] [CrossRef]
- Yang, N.; Li, Z.; Jiao, Z.; Gu, P.; Zhou, Y.; Lu, L.; Chou, K.Y. A Trichosanthin-derived peptide suppresses type 1 immune responses by TLR2-dependent activation of CD8(+)CD28(−) Tregs. Clin. Immunol. 2014, 153, 277–287. [Google Scholar] [CrossRef]
- Sun, D.; Qin, Y.; Chluba, J.; Epplen, J.T.; Wekerle, H. Suppression of experimentally induced autoimmune encephalomyelitis by cytolytic T-T cell interactions. Nature 1988, 332, 843–845. [Google Scholar] [CrossRef]
- Zhang, J.; Medaer, R.; Stinissen, P.; Hafler, D.; Raus, J. MHC-restricted depletion of human myelin basic protein-reactive T cells by T cell vaccination. Science 1993, 261, 1451–1454. [Google Scholar] [CrossRef]
- Jiang, H.; Kashleva, H.; Xu, L.X.; Forman, J.; Flaherty, L.; Pernis, B.; Braunstein, N.S.; Chess, L. T cell vaccination induces T cell receptor Vbeta-specific Qa-1-restricted regulatory CD8(+) T cells. Proc. Natl. Acad. Sci. USA 1998, 95, 4533–4537. [Google Scholar] [CrossRef] [Green Version]
- Davila, E.; Kang, Y.M.; Park, Y.W.; Sawai, H.; He, X.; Pryshchep, S.; Goronzy, J.J.; Weyand, C.M. Cell-based immunotherapy with suppressor CD8+ T cells in rheumatoid arthritis. J. Immunol. 2005, 174, 7292–7301. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.; Davies, R.; Williams, A.; Jones, G.; Choy, E.H.S. CD28(−) Cells Are Increased in Early Rheumatoid Arthritis and Are Linked With Cytomegalovirus Status. Front. Med. 2020, 7, 129. [Google Scholar] [CrossRef]
- Zizzo, G.; De Santis, M.; Bosello, S.; Tolusso, B.; Alivernini, S.; De Luca, G.; Gremese, E.; Ferraccioli, G. Measuring the T-cell down-regulation of TCR-zeta, ZAP-70 and CD28 in arthritis patients: An old tool for new biomarkers. Eur. J. Immunol. 2019, 49, 2195–2203. [Google Scholar] [CrossRef]
- Hahn, B.H.; Singh, R.P.; La Cava, A.; Ebling, F.M. Tolerogenic treatment of lupus mice with consensus peptide induces Foxp3-expressing, apoptosis-resistant, TGFbeta-secreting CD8+ T cell suppressors. J. Immunol. 2005, 175, 7728–7737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; La Cava, A.; Wong, M.; Ebling, F.; Hahn, B.H. CD8+ T cell-mediated suppression of autoimmunity in a murine lupus model of peptide-induced immune tolerance depends on Foxp3 expression. J. Immunol. 2007, 178, 7649–7657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; La Cava, A.; Hahn, B.H. pConsensus peptide induces tolerogenic CD8+ T cells in lupus-prone (NZB x NZW)F1 mice by differentially regulating Foxp3 and PD1 molecules. J. Immunol. 2008, 180, 2069–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabinska, M.; Krajewska, M.; Koscielska-Kasprzak, K.; Klinger, M. CD3(+)CD8(+)CD28(−) T Lymphocytes in Patients with Lupus Nephritis. J. Immunol. Res. 2016, 2016, 1058165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisikirska, B.; Colgan, J.; Luban, J.; Bluestone, J.A.; Herold, K.C. TCR stimulation with modified anti-CD3 mAb expands CD8+ T cell population and induces CD8+CD25+ Tregs. J. Clin. Invest. 2005, 115, 2904–2913. [Google Scholar] [CrossRef] [Green Version]
- Shimokawa, C.; Kato, T.; Takeuchi, T.; Ohshima, N.; Furuki, T.; Ohtsu, Y.; Suzue, K.; Imai, T.; Obi, S.; Olia, A.; et al. CD8(+) regulatory T cells are critical in prevention of autoimmune-mediated diabetes. Nat. Commun. 2020, 11, 1922. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Larregina, A.T.; Domsic, R.T.; Stolz, D.B.; Medsger, T.A., Jr.; Lafyatis, R.; Fuschiotti, P. Skin-Resident Effector Memory CD8(+)CD28(−) T Cells Exhibit a Profibrotic Phenotype in Patients with Systemic Sclerosis. J. Investig. Dermatol. 2017, 137, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, D.; Battaglia, F.; Parodi, A.; Stringara, S.; Negrini, S.; Panico, N.; Rizzi, M.; Kalli, F.; Conteduca, G.; Ghio, M.; et al. Alteration of Th17 and Treg cell subpopulations co-exist in patients affected with systemic sclerosis. Clin. Immunol. 2011, 139, 249–257. [Google Scholar] [CrossRef]
- Menager-Marcq, I.; Pomie, C.; Romagnoli, P.; van Meerwijk, J.P. CD8+CD28− regulatory T lymphocytes prevent experimental inflammatory bowel disease in mice. Gastroenterology 2006, 131, 1775–1785. [Google Scholar] [CrossRef] [Green Version]
- Pomie, C.; Vicente, R.; Vuddamalay, Y.; Lundgren, B.A.; van der Hoek, M.; Enault, G.; Kagan, J.; Fazilleau, N.; Scott, H.S.; Romagnoli, P.; et al. Autoimmune regulator (AIRE)-deficient CD8+CD28low regulatory T lymphocytes fail to control experimental colitis. Proc. Natl. Acad. Sci. USA 2011, 108, 12437–12442. [Google Scholar] [CrossRef] [Green Version]
- Kawakita, A.; Shirasaki, H.; Yasutomi, M.; Tokuriki, S.; Mayumi, M.; Naiki, H.; Ohshima, Y. Immunotherapy with oligomannose-coated liposomes ameliorates allergic symptoms in a murine food allergy model. Allergy 2012, 67, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Gu, H.; Lin, Q.; Xing, T.; Chen, M.; Zhong, T.; Wu, G.; Feng, Y.; Liu, H.; Gao, Y.; et al. Disequilibrium in the CD8(+)CD28(+)/CD8(+)CD28(−) T Lymphocyte Balance Is Related to Prognosis in Rats with Trinitrobenzenesulfonic Acid-Induced Colitis. Dig. Dis. Sci. 2017, 62, 639–651. [Google Scholar] [CrossRef]
- Dai, S.X.; Gu, H.X.; Lin, Q.Y.; Wu, Y.K.; Wang, X.Y.; Huang, S.Z.; Xing, T.S.; Chen, M.H.; Zhang, Q.F.; Zheng, Z.W.; et al. Decreased CD8+CD28+/CD8+CD28− T cell ratio can sensitively predict poor outcome for patients with complicated Crohn disease. Medicine 2017, 96, e7247. [Google Scholar] [CrossRef]
- Brimnes, J.; Allez, M.; Dotan, I.; Shao, L.; Nakazawa, A.; Mayer, L. Defects in CD8+ regulatory T cells in the lamina propria of patients with inflammatory bowel disease. J. Immunol. 2005, 174, 5814–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toy, L.S.; Yio, X.Y.; Lin, A.; Honig, S.; Mayer, L. Defective expression of gp180, a novel CD8 ligand on intestinal epithelial cells, in inflammatory bowel disease. J. Clin. Investig. 1997, 100, 2062–2071. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.H.; Xu, P.; Yu, X.; Song, H.F.; Chen, H.; Zhang, X.G.; Wu, M.Y.; Wang, X.F. Discrepant Clinical Significance of CD28(+)CD8(−) and CD4(+)CD25(high) Regulatory T Cells During the Progression of Hepatitis B Virus Infection. Viral Immunol. 2018, 31, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Saukkonen, J.J.; Kornfeld, H.; Berman, J.S. Expansion of a CD8+CD28− cell population in the blood and lung of HIV-positive patients. J. Acquir. Immune Defic. Syndr. 1993, 6, 1194–1204. [Google Scholar]
- Klatt, T.; Ouyang, Q.; Flad, T.; Koetter, I.; Buhring, H.J.; Kalbacher, H.; Pawelec, G.; Muller, C.A. Expansion of peripheral CD8+ CD28− T cells in response to Epstein-Barr virus in patients with rheumatoid arthritis. J. Rheumatol. 2005, 32, 239–251. [Google Scholar]
- Granito, A.; Muratori, L.; Lalanne, C.; Quarneti, C.; Ferri, S.; Guidi, M.; Lenzi, M.; Muratori, P. Hepatocellular carcinoma in viral and autoimmune liver diseases: Role of CD4+ CD25+ Foxp3+ regulatory T cells in the immune microenvironment. World J. Gastroenterol. 2021, 27, 2994–3009. [Google Scholar] [CrossRef]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: New mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Ren, J.; Zhou, X.; Wang, X.; Song, G.; Hobeika, A.; Yuan, Y.; Lyerly, H.K. Circulating CD8(+)CD28(−) suppressor T cells tied to poorer prognosis among metastatic breast cancer patients receiving adoptive T-cell therapy: A cohort study. Cytotherapy 2018, 20, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Parodi, A.; Battaglia, F.; Kalli, F.; Ferrera, F.; Conteduca, G.; Tardito, S.; Stringara, S.; Ivaldi, F.; Negrini, S.; Borgonovo, G.; et al. CD39 is highly involved in mediating the suppression activity of tumor-infiltrating CD8+ T regulatory lymphocytes. Cancer Immunol. Immunother. 2013, 62, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Robert, C. A decade of immune-checkpoint inhibitors in cancer therapy. Nat. Commun. 2020, 11, 3801. [Google Scholar] [CrossRef]
- Saleh, R.; Elkord, E. FoxP3(+) T regulatory cells in cancer: Prognostic biomarkers and therapeutic targets. Cancer Lett. 2020, 490, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, R.; DelaRosa, O.; Alonso, C.; Ostos, B.; Espejo, J.; Pena, J.; Solana, R. Increased expression of NK cell markers on T lymphocytes in aging and chronic activation of the immune system reflects the accumulation of effector/senescent T cells. Mech. Ageing Dev. 2000, 121, 77–88. [Google Scholar] [CrossRef]
- Labalette, M.; Leteurtre, E.; Thumerelle, C.; Grutzmacher, C.; Tourvieille, B.; Dessaint, J.P. Peripheral human CD8(+)CD28(+)T lymphocytes give rise to CD28(−)progeny, but IL-4 prevents loss of CD28 expression. Int. Immunol. 1999, 11, 1327–1336. [Google Scholar] [CrossRef] [Green Version]
- Bezie, S.; Charreau, B.; Vimond, N.; Lasselin, J.; Gerard, N.; Nerriere-Daguin, V.; Bellier-Waast, F.; Duteille, F.; Anegon, I.; Guillonneau, C. Human CD8+ Tregs expressing a MHC-specific CAR display enhanced suppression of human skin rejection and GVHD in NSG mice. Blood Adv. 2019, 3, 3522–3538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceeraz, S.; Thompson, C.R.; Beatson, R.; Choy, E.H. Harnessing CD8+CD28− Regulatory T Cells as a Tool to Treat Autoimmune Disease. Cells 2021, 10, 2973. https://doi.org/10.3390/cells10112973
Ceeraz S, Thompson CR, Beatson R, Choy EH. Harnessing CD8+CD28− Regulatory T Cells as a Tool to Treat Autoimmune Disease. Cells. 2021; 10(11):2973. https://doi.org/10.3390/cells10112973
Chicago/Turabian StyleCeeraz, Sabrina, Charlotte R. Thompson, Richard Beatson, and Ernest H. Choy. 2021. "Harnessing CD8+CD28− Regulatory T Cells as a Tool to Treat Autoimmune Disease" Cells 10, no. 11: 2973. https://doi.org/10.3390/cells10112973
APA StyleCeeraz, S., Thompson, C. R., Beatson, R., & Choy, E. H. (2021). Harnessing CD8+CD28− Regulatory T Cells as a Tool to Treat Autoimmune Disease. Cells, 10(11), 2973. https://doi.org/10.3390/cells10112973