
Gfi1 Loss Protects against Two Models of Induced Diabetes

 , , ,
, , ,  , ,
, , 
Abstract
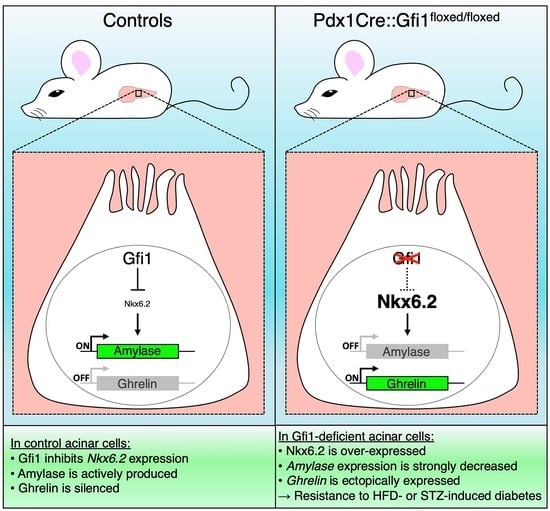
:
1. Introduction
2. Materials and Methods
2.1. Generation of Gf1Cre::RosaLac and Pdx1Cre::Gfi1cko Mouse Lines
2.2. Animal Manipulations
2.3. Gene Expression Analyses
2.4. Immunohistochemistry
2.5. ELISA Immunoassay
2.6. X-Gal Staining
2.7. RNAscope
2.8. Oligosaccharides Analyses in Fecal Samples
2.9. Quantification and Data Analysis
3. Results
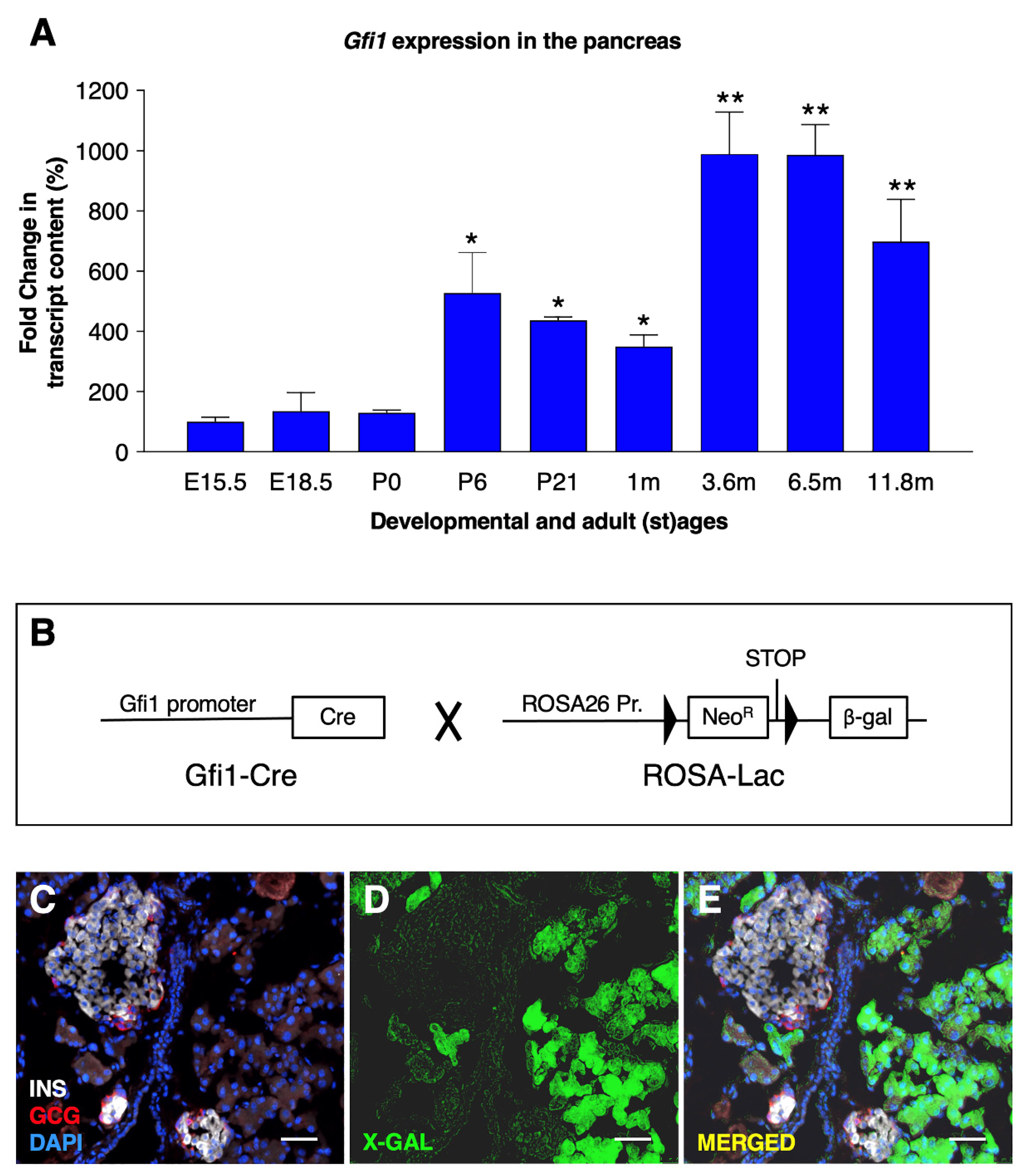
3.1. Gfi1 Expression in Murine Pancreas
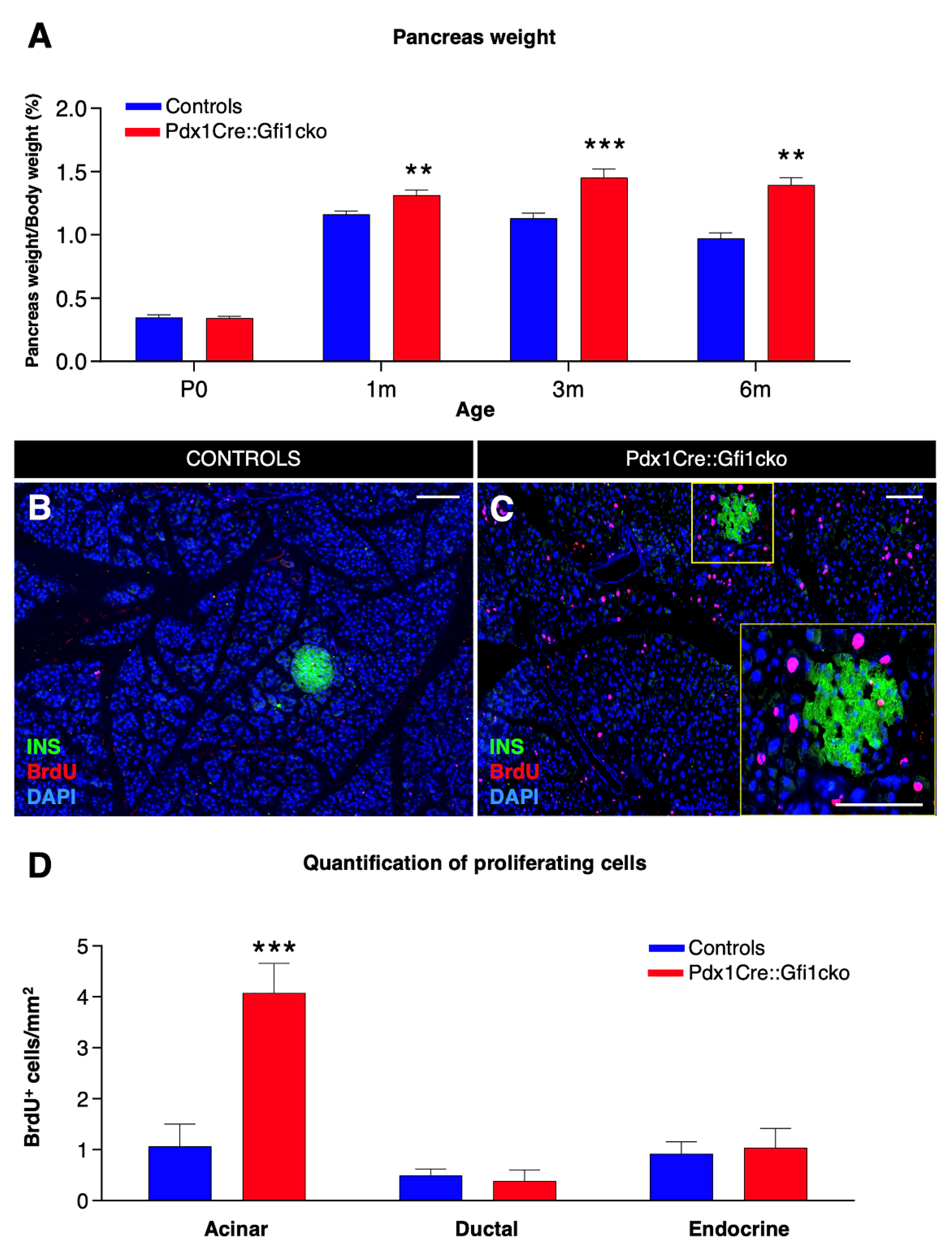
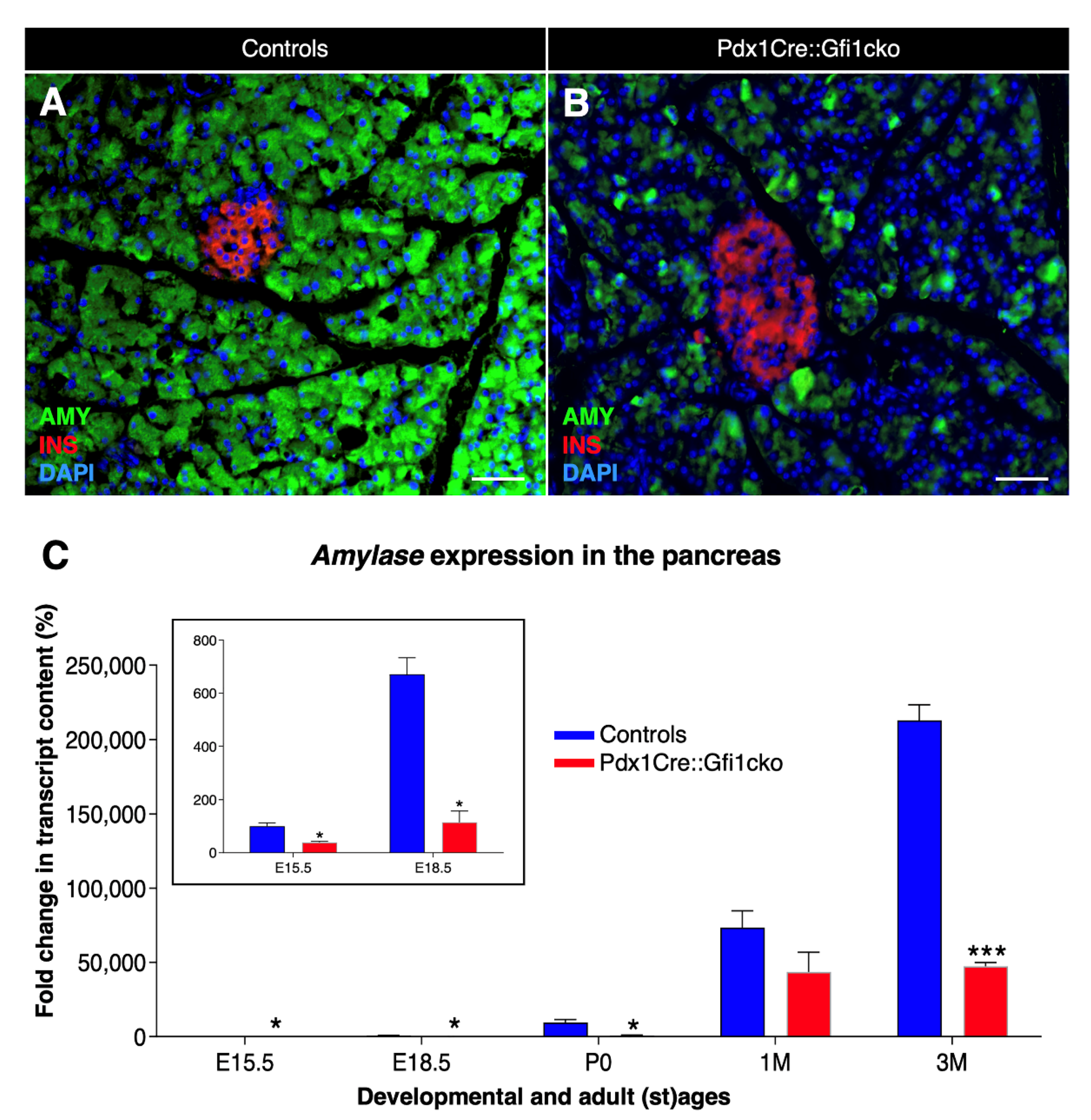
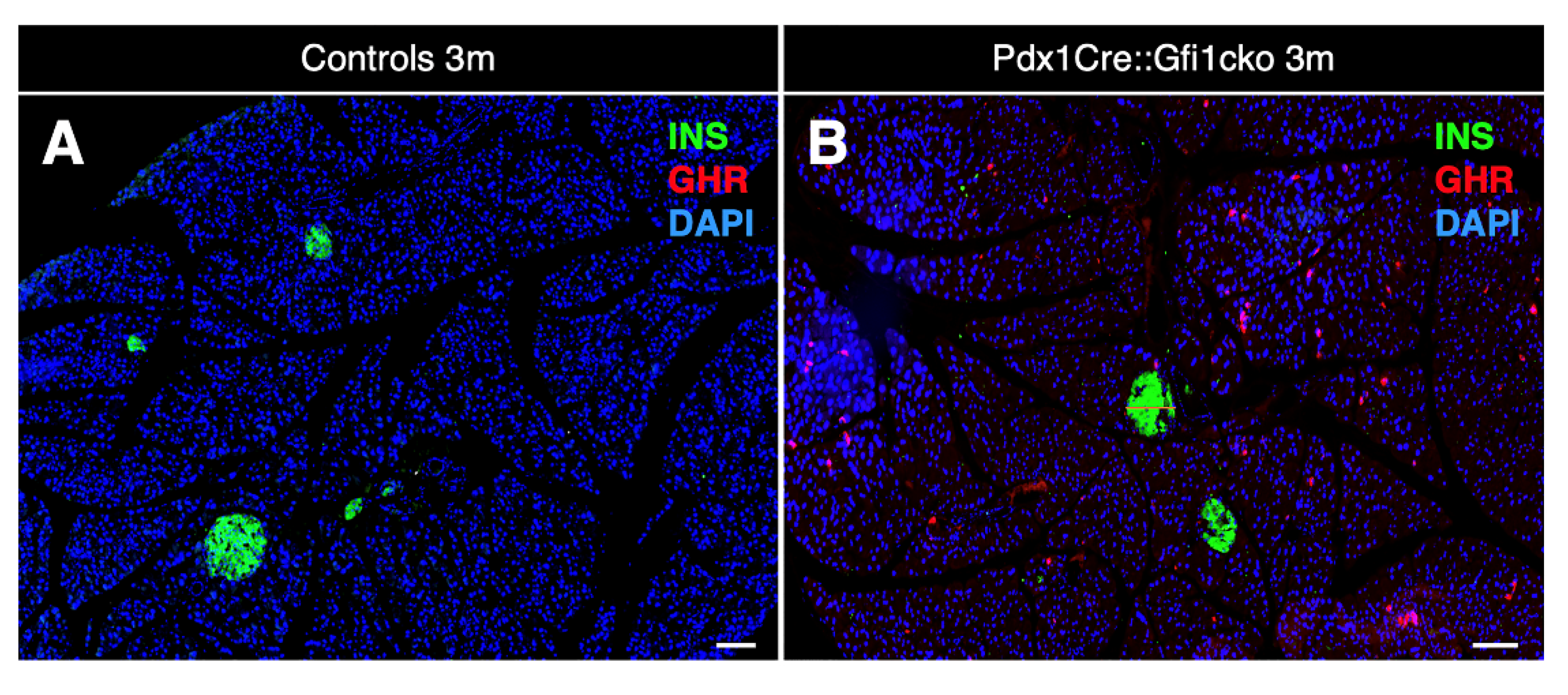
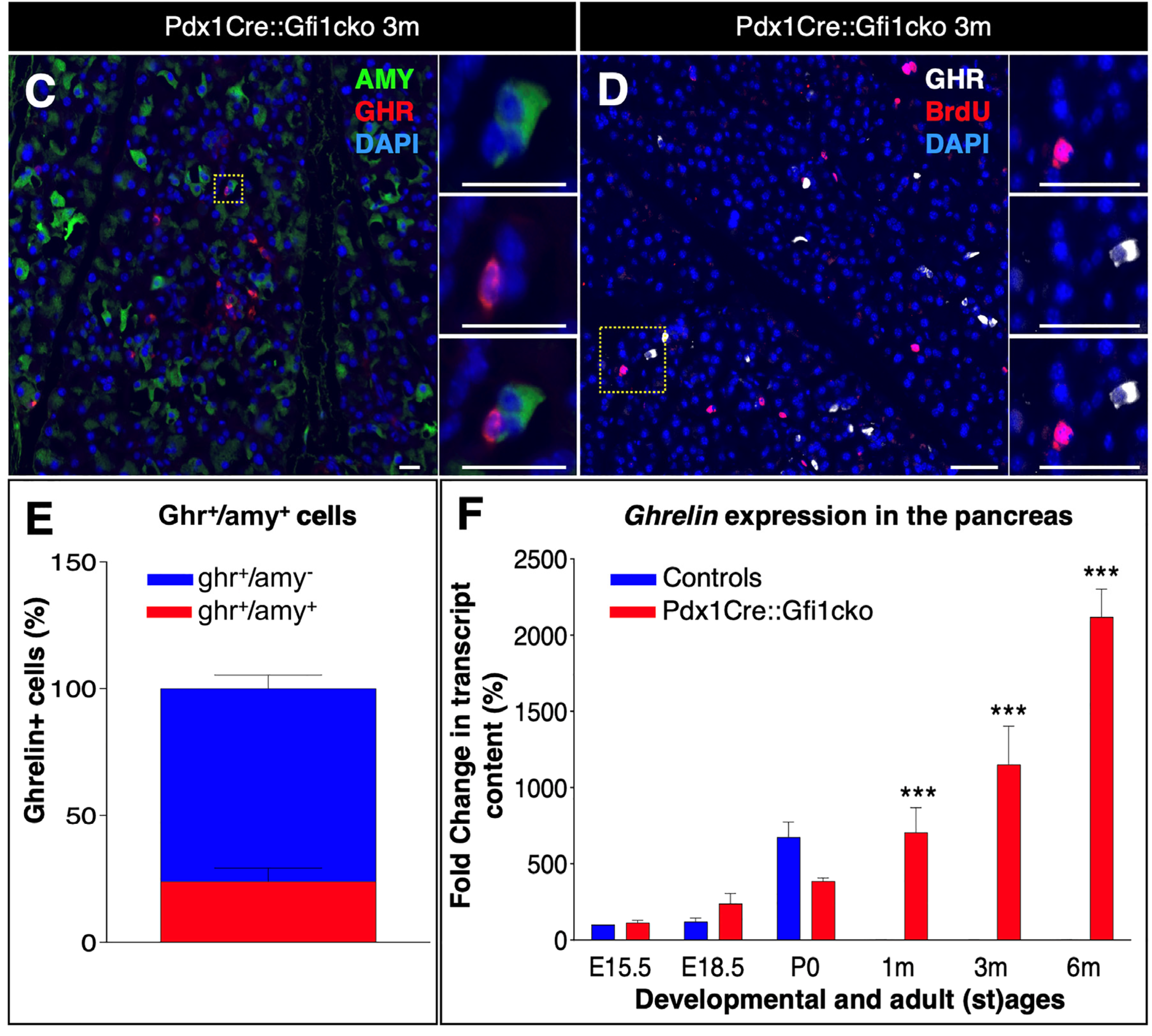
3.2. Loss of Gfi1 Affects the Maturation of Acinar Cells
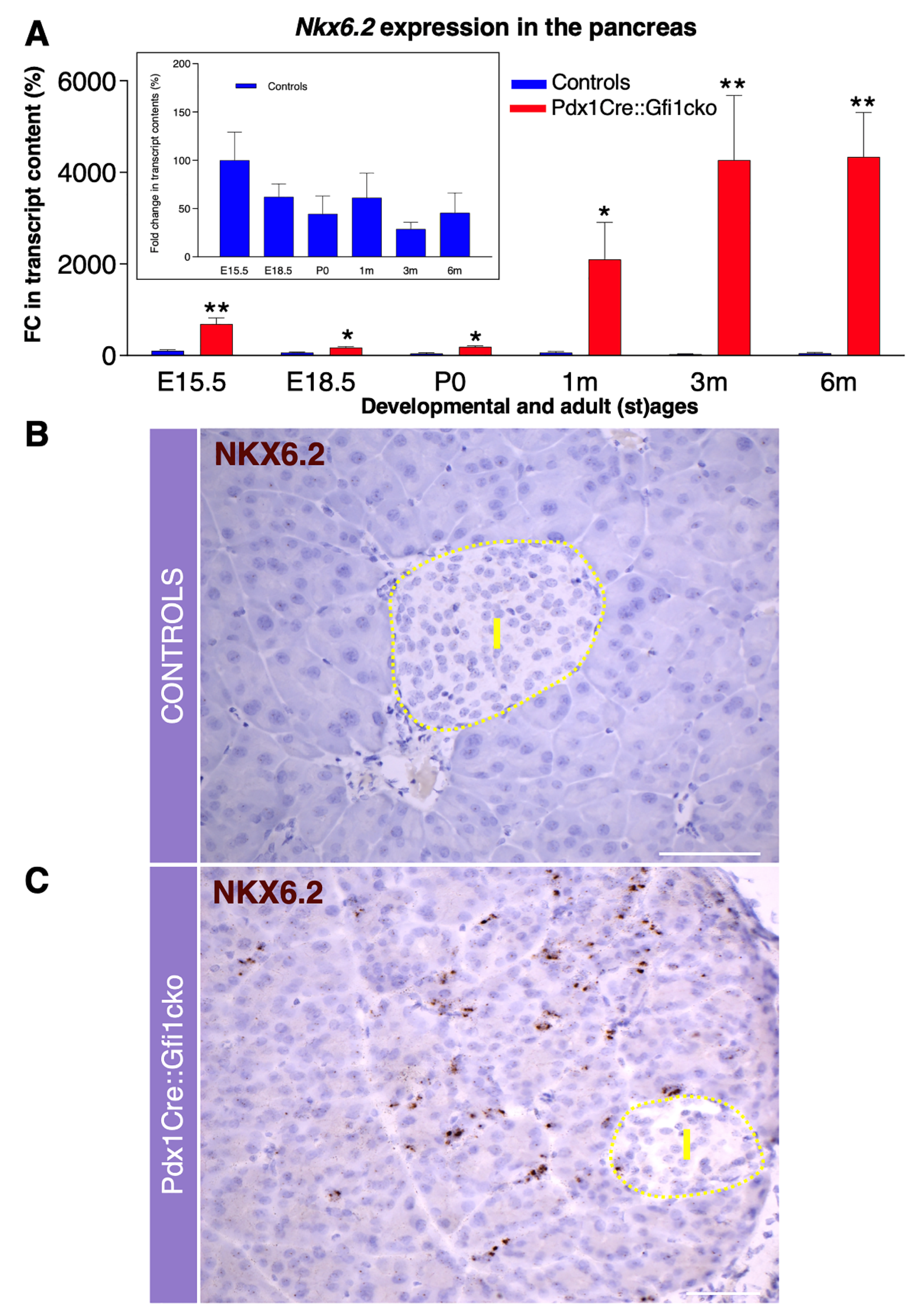
3.3. Gfi1 Mutants Display an Aberrant Overexpression of Nkx6.2
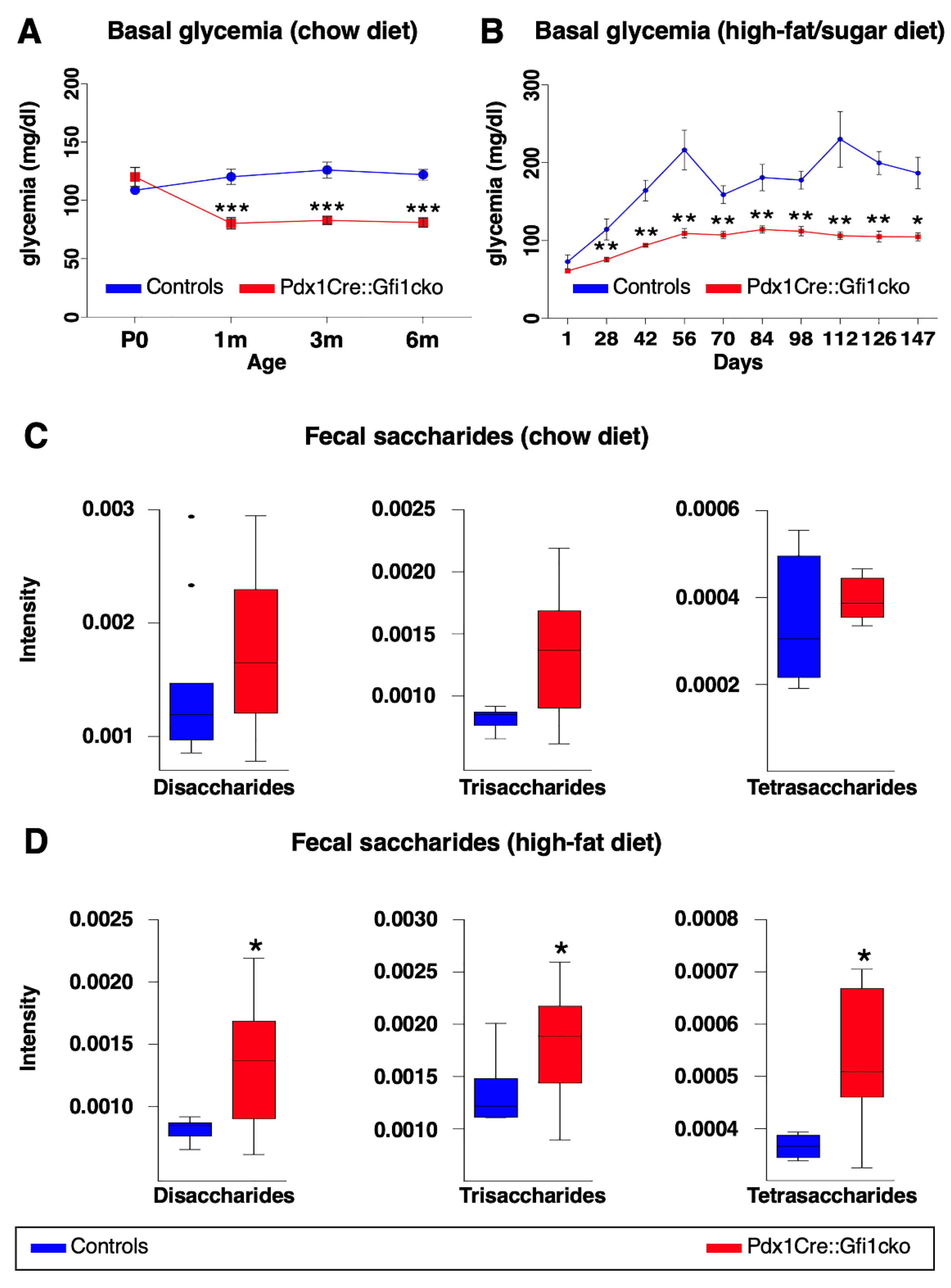
3.4. Pdx1Cre::Gfi1cko Mice Appear Resistant to Diet-Induced Hyperglycemia
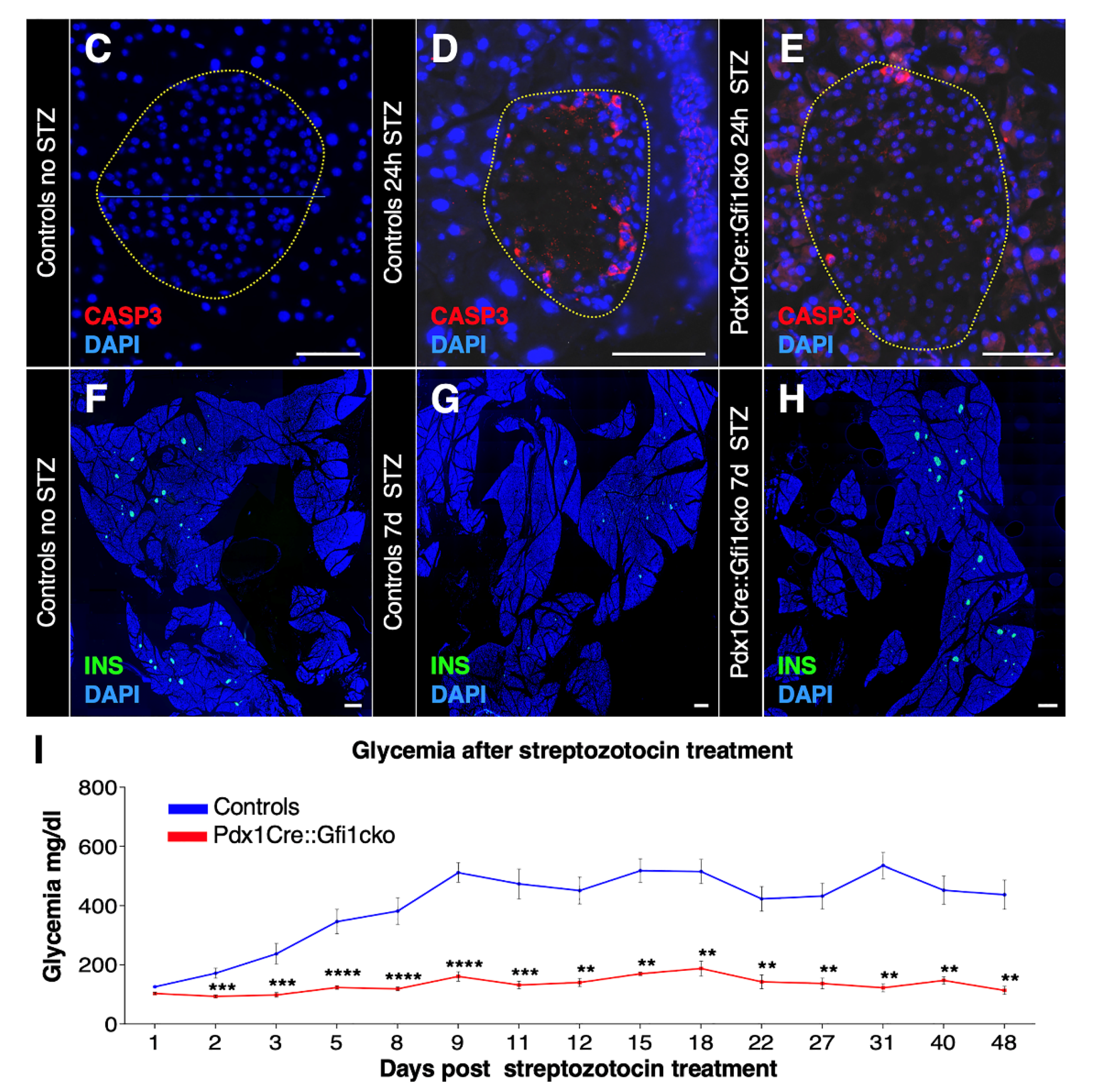
3.5. Loss of Gfi1 Protects Mice against STZ-Induced Diabetes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitcomb, D.C.; Lowe, M.E. Human pancreatic digestive enzymes. Dig. Dis. Sci. 2007, 52, 1–17. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, S.A.; Mukherjee, S. Physiology, Pancreas; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Roder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [PubMed]
- Rindi, G.; Necchi, V.; Savio, A.; Torsello, A.; Zoli, M.; Locatelli, V.; Raimondo, F.; Cocchi, D.; Solcia, E. Characterisation of gastric ghrelin cells in man and other mammals: Studies in adult and fetal tissues. Histochem. Cell Biol. 2002, 117, 511–519. [Google Scholar] [CrossRef]
- Andralojc, K.M.; Mercalli, A.; Nowak, K.W.; Albarello, L.; Calcagno, R.; Luzi, L.; Bonifacio, E.; Doglioni, C.; Piemonti, L. Ghrelin-producing epsilon cells in the developing and adult human pancreas. Diabetologia 2009, 52, 486–493. [Google Scholar] [CrossRef]
- Granata, R.; Settanni, F.; Biancone, L.; Trovato, L.; Nano, R.; Bertuzzi, F.; Destefanis, S.; Annunziata, M.; Martinetti, M.; Catapano, F.; et al. Acylated and unacylated ghrelin promote proliferation and inhibit apoptosis of pancreatic beta-cells and human islets: Involvement of 3′,5′-cyclic adenosine monophosphate/protein kinase A, extracellular signal-regulated kinase 1/2, and phosphatidyl inositol 3-Kinase/Akt signaling. Endocrinology 2007, 148, 512–529. [Google Scholar]
- Katsarou, A.; Gudbjornsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, A. Type 1 diabetes mellitus. Nat. Rev. Dis. Primers 2017, 3, 17016. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- McCarthy, M.I. Genomics, type 2 diabetes, and obesity. N. Engl. J. Med. 2010, 363, 2339–2350. [Google Scholar] [CrossRef]
- Bonnefond, A.; Froguel, P.; Vaxillaire, M. The emerging genetics of type 2 diabetes. Trends Mol. Med. 2010, 16, 407–416. [Google Scholar] [CrossRef]
- Burlison, J.S.; Long, Q.; Fujitani, Y.; Wright, C.V.; Magnuson, M.A. Pdx-1 and Ptf1a concurrently determine fate specification of pancreatic multipotent progenitor cells. Dev. Biol. 2008, 316, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Guz, Y.; Montminy, M.R.; Stein, R.; Leonard, J.; Gamer, L.W.; Wright, C.V.; Teitelman, G. Expression of murine STF-1, a putative insulin gene transcription factor, in beta cells of pancreas, duodenal epithelium and pancreatic exocrine and endocrine progenitors during ontogeny. Development 1995, 121, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.C.; Wright, C. Pancreas organogenesis: From bud to plexus to gland. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2011, 240, 530–565. [Google Scholar] [CrossRef]
- Shih, H.P.; Wang, A.; Sander, M. Pancreas organogenesis: From lineage determination to morphogenesis. Annu. Rev. Cell Dev. Biol. 2013, 29, 81–105. [Google Scholar] [CrossRef]
- Schaffer, A.E.; Freude, K.K.; Nelson, S.B.; Sander, M. Nkx6 transcription factors and Ptf1a function as antagonistic lineage determinants in multipotent pancreatic progenitors. Dev. Cell 2010, 18, 1022–1029. [Google Scholar] [CrossRef]
- Sarkar, S.A.; Lee, C.E.; Tipney, H.; Karimpour-Fard, A.; Dinella, J.D.; Juhl, K.; Walters, J.A.; Hutton, J.C.; Hunter, L.E. Synergizing genomic analysis with biological knowledge to identify and validate novel genes in pancreatic development. Pancreas 2012, 41, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Zweidler-McKay, P.A.; Grimes, H.L.; Flubacher, M.M.; Tsichlis, P.N. Gfi-1 encodes a nuclear zinc finger protein that binds DNA and functions as a transcriptional repressor. Mol. Cell. Biol. 1996, 16, 4024–4034. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Nyeng, P.; Xiao, F.; Dorantes, J.; Jensen, J. Growth Factor Independence-1 (Gfi1) Is Required for Pancreatic Acinar Unit Formation and Centroacinar Cell Differentiation. CMGH Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 233–247.e1. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Gan, J.; Xie, X.; Deng, M.; Feng, L.; Chen, X.; Gao, Z.; Gan, L. Gfi1-Cre knock-in mouse line: A tool for inner ear hair cell-specific gene deletion. Genesis 2010, 48, 400–406. [Google Scholar] [CrossRef]
- Soriano, P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat. Genet. 1999, 21, 70–71. [Google Scholar] [CrossRef]
- Gannon, M.; Herrera, P.L.; Wright, C.V. Mosaic Cre-mediated recombination in pancreas using the pdx-1 enhancer/promoter. Genesis 2000, 26, 143–144. [Google Scholar] [CrossRef]
- Zhu, J.; Jankovic, D.; Grinberg, A.; Guo, L.; Paul, W.E. Gfi-1 plays an important role in IL-2-mediated Th2 cell expansion. Proc. Natl. Acad. Sci. USA 2006, 103, 18214–18219. [Google Scholar] [CrossRef]
- Collombat, P.; Mansouri, A.; Hecksher-Sorensen, J.; Serup, P.; Krull, J.; Gradwohl, G.; Gruss, P. Opposing actions of Arx and Pax4 in endocrine pancreas development. Genes Dev. 2003, 17, 2591–2603. [Google Scholar] [CrossRef]
- Tziotis, D.; Hertkorn, N.; Schmitt-Kopplin, P. Kendrick-analogous network visualisation of ion cyclotron resonance Fourier transform mass spectra: Improved options for the assignment of elemental compositions and the classification of organic molecular complexity. Eur. J. Mass Spectrom. (Chichester) 2011, 17, 415–421. [Google Scholar] [CrossRef]
- Moritz, F.; Kaling, M.; Schnitzler, J.P.; Schmitt-Kopplin, P. Characterization of poplar metabotypes via mass difference enrichment analysis. Plant Cell Environ. 2017, 40, 1057–1073. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Druelle, N.; Vieira, A.; Shabro, A.; Courtney, M.; Mondin, M.; Rekima, S.; Napolitano, T.; Silvano, S.; Navarro-Sanz, S.; Hadzic, B.; et al. Ectopic expression of Pax4 in pancreatic delta cells results in beta-like cell neogenesis. J. Cell Biol. 2017, 216, 4299–4311. [Google Scholar] [CrossRef] [PubMed]
- Arnes, L.; Hill, J.T.; Gross, S.; Magnuson, M.A.; Sussel, L. Ghrelin expression in the mouse pancreas defines a unique multipotent progenitor population. PLoS ONE 2012, 7, e52026. [Google Scholar] [CrossRef] [PubMed]
- Henseleit, K.D.; Nelson, S.B.; Kuhlbrodt, K.; Hennings, J.C.; Ericson, J.; Sander, M. NKX6 transcription factor activity is required for alpha- and beta-cell development in the pancreas. Development 2005, 132, 3139–3149. [Google Scholar] [CrossRef]
- Shintani, M.; Ogawa, Y.; Ebihara, K.; Aizawa-Abe, M.; Miyanaga, F.; Takaya, K.; Hayashi, T.; Inoue, G.; Hosoda, K.; Kojima, M.; et al. Ghrelin, an endogenous growth hormone secretagogue, is a novel orexigenic peptide that antagonizes leptin action through the activation of hypothalamic neuropeptide Y/Y1 receptor pathway. Diabetes 2001, 50, 227–232. [Google Scholar] [CrossRef]
- Adamik, J.; Silbermann, R.; Marino, S.; Sun, Q.; Anderson, J.L.; Zhou, D.; Xie, X.Q.; Roodman, G.D.; Galson, D.L. XRK3F2 Inhibition of p62-ZZ Domain Signaling Rescues Myeloma-Induced GFI1-Driven Epigenetic Repression of the Runx2 Gene in Pre-osteoblasts to Overcome Differentiation Suppression. Front. Endocrinol. (Lausanne) 2018, 9, 344. [Google Scholar] [CrossRef]
- Prado, C.L.; Pugh-Bernard, A.E.; Elghazi, L.; Sosa-Pineda, B.; Sussel, L. Ghrelin cells replace insulin-producing beta cells in two mouse models of pancreas development. Proc. Natl. Acad. Sci. USA 2004, 101, 2924–2929. [Google Scholar] [CrossRef] [PubMed]
- van der Meer, L.T.; Jansen, J.H.; van der Reijden, B.A. Gfi1 and Gfi1b: Key regulators of hematopoiesis. Leukemia 2010, 24, 1834–1843. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Elghazi, L.; Martin, S.; Martins, I.; Srinivasan, R.S.; Geng, X.; Sleeman, M.; Collombat, P.; Houghton, J.; Sosa-Pineda, B. Ghrelin is a novel target of Pax4 in endocrine progenitors of the pancreas and duodenum. Dev. Dyn. An. Off. Publ. Am. Assoc. Anat. 2008, 237, 51–61. [Google Scholar]
- Warzecha, Z.; Ceranowicz, P.; Dembinski, A.; Cieszkowski, J.; Kusnierz-Cabala, B.; Tomaszewska, R.; Kuwahara, A.; Kato, I. Therapeutic effect of ghrelin in the course of cerulein-induced acute pancreatitis in rats. J. Physiol. Pharm. 2010, 61, 419–427. [Google Scholar]
- Zhang, Y.; Ying, B.; Shi, L.; Fan, H.; Yang, D.; Xu, D.; Wei, Y.; Hu, X.; Zhang, Y.; Zhang, X. Ghrelin inhibit cell apoptosis in pancreatic β cell line HIT-T15 via mitogen-activated protein kinase/phosphoinositide 3-kinase pathways. Toxicology 2007, 237, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, L.; Zhao, B.; Jiao, A.; Li, X.; Sun, N.; Zhang, J. Ghrelin Protects against Dexamethasone-Induced INS-1 Cell Apoptosis via ERK and p38MAPK Signaling. Int. J. Endocrinol. 2016, 2016, 4513051. [Google Scholar] [CrossRef]
- Irako, T.; Akamizu, T.; Hosoda, H.; Iwakura, H.; Ariyasu, H.; Tojo, K.; Tajima, N.; Kangawa, K. Ghrelin prevents development of diabetes at adult age in streptozotocin-treated newborn rats. Diabetologia 2006, 49, 1264–1273. [Google Scholar] [CrossRef]
- Navarro, J.A.; Decara, J.; Medina-Vera, D.; Tovar, R.; Suarez, J.; Pavon, J.; Serrano, A.; Vida, M.; Gutierrez-Adan, A.; Sanjuan, C.; et al. D-Pinitol from Ceratonia siliqua Is an Orally Active Natural Inositol That Reduces Pancreas Insulin Secretion and Increases Circulating Ghrelin Levels in Wistar Rats. Nutrients 2020, 12, 2030. [Google Scholar] [CrossRef]
- Chanclon, B.; Martinez-Fuentes, A.J.; Gracia-Navarro, F. Role of SST, CORT and ghrelin and its receptors at the endocrine pancreas. Front. Endocrinol. (Lausanne) 2012, 3, 114. [Google Scholar]
- Kaiser, J.; Krippeit-Drews, P.; Drews, G. Acyl-Ghrelin Influences Pancreatic beta-Cell Function by Interference with KATP Channels. Diabetes 2021, 70, 423–435. [Google Scholar] [CrossRef]
- DiGruccio, M.R.; Mawla, A.M.; Donaldson, C.J.; Noguchi, G.M.; Vaughan, J.; Cowing-Zitron, C.; van der Meulen, T.; Huising, M.O. Comprehensive alpha, beta and delta cell transcriptomes reveal that ghrelin selectively activates delta cells and promotes somatostatin release from pancreatic islets. Mol. Metab. 2016, 5, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, A.E.; Svendsen, B.; Lam, B.Y.; Yeo, G.S.; Holst, J.J.; Reimann, F.; Gribble, F.M. Transcriptomic profiling of pancreatic alpha, beta and delta cell populations identifies delta cells as a principal target for ghrelin in mouse islets. Diabetologia 2016, 59, 2156–2165. [Google Scholar] [CrossRef] [PubMed]
- Dezaki, K.; Kakei, M.; Yada, T. Ghrelin uses Gαi2 and activates voltage-dependent K+ channels to attenuate glucose-induced Ca2+ signaling and insulin release in islet β-cells: Novel signal transduction of ghrelin. Diabetes 2007, 56, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Huang, L.; Huang, Z.; Feng, D.; Clark, R.J.; Chen, C. LEAP-2: An Emerging Endogenous Ghrelin Receptor Antagonist in the Pathophysiology of Obesity. Front. Endocrinol. (Lausanne) 2021, 12, 717544. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resources | Source | Identifier |
|---|---|---|
| Antibodies | ||
| Guinea pig polyclonal anti-insulin | Dako, Hovedstaden, Denmark | Ref: A0564 |
| Rat monoclonal anti-BrdU | Abcam, Cambridge, UK | Ref: ab6326 |
| Rabbit polyclonal anti-pancreatic amylase | ThermoFisher, Waltham, MA, USA | Ref: PA5-25330 |
| Mouse monoclonal anti-ghrelin | Merck, Readington Township, NJ, USA | Ref: MAB10404 |
| Rabbit polyclonal active caspase-3 | R&D Systems, Minneapolis, MN, USA | Ref: AF835 |
| Rabbit monoclonal anti-proglucagon | Cell Signaling, Danvers, MA, USA | Ref: 8233 |
| Rabbit polyclonal anti-somatostatin | Dako, Hovedstaden, Denmark | Ref: A0566 |
| Rabbit polyclonal anti-pancreatic polypeptide | Millipore, Burlington, MA, USA | Ref: AB939 |
| Goat anti-guinea pig IgG (H+L) Alexa Fluor 488 conjugated | ThermoFisher, Waltham, MA, USA | Ref: A-11073 |
| Goat anti-guinea pig IgG (H+L) Alexa Fluor 594 conjugated | ThermoFisher, Waltham, MA, USA | Ref: A-11076 |
| Goat anti-rat IgG (H+L) Alexa Fluor 594 conjugated | ThermoFisher, Waltham, MA, USA | Ref: A-11007 |
| Donkey anti-Rabbit IgG (H+L) Alexa Fluor 488 conjugated | ThermoFisher, Waltham, MA, USA | Ref: A-21206 |
| Goat anti-Mouse IgG (H+L) Alexa Fluor 594 conjugated | ThermoFisher, Waltham, MA, USA | Ref: A-11005 |
| Donkey anti-Rabbit IgG (H+L) Alexa Fluor 594 conjugated | ThermoFisher, Waltham, MA, USA | Ref: A-21207 |
| Experimental Models: Organisms/Strains | ||
| Pdx1-Cre | [21] | N/A |
| Gfi1cko | [22] | N/A |
| Gfi1-Cre | [19] | N/A |
| ROSA26-lox-Stop-lox-β-Gal | [20] | N/A |
| Chemicals, Peptides, and Recombinant Proteins | ||
| d-(+)-glucose | Sigma-Aldrich, St. Louis, MO, USA | Ref: G7528-1KG |
| Insulin | Novo Nordisk, Bagsvaerd Denmark | N/A |
| Streptozocin | Sigma-Aldrich, St. Louis, MO, USA | Ref: S0130 |
| RNAlater | Invitrogen, Waltham, MA, USA | Ref: AM7021 |
| Collagenase | Sigma-Aldrich, St. Louis, MO, USA | Ref: C7657 |
| Histopaque-1077 | Sigma-Aldrich, St. Louis, MO, USA | Ref: 10771 |
| Histopaque-1119 | Sigma-Aldrich, St. Louis, MO, USA | Ref: 11191 |
| Antigenfix | Microm Microtech, Brignais, France | Ref: F/P0016 |
| Antigen Unmasking Solution | CliniSciences, Nanterre, France | Ref: H-3300 |
| Tissue freezing medium | Leica, Wetzlar, Germany | Ref: 14020108926 |
| Amylase from porcine pancreas | Sigma-Aldrich, St. Louis, MO, USA | Ref: A6255 |
| Critical Commercial Assays | ||
| RNeasy Mini Kit | Qiagen, Hilden, Grmany | Ref: 74104 |
| Unacylated Ghrelin (mouse, rat) Express Enzyme Immunoassay kit | Bertin Bioreagent, Frankfurt am Main Germany | Ref: A05118 |
| Acylated Ghrelin (mouse, rat) Express Enzyme Immunoassay kit | Bertin Bioreagent, Frankfurt am Main Germany | Ref: A05117 |
| Pierce™ BCA Protein Assay Kit | ThermoFisher, Waltham, MA, USA | Ref: 23225 |
| RNAscope 2.5 HD Reagent Kit-BROWN | Advanced Cell Diagnostics, Newark, CA, USA | Ref: 322370 |
| Software and Algorithms | ||
| Prism software (Graphpad) | GraphPad Software, Inc. 7 | N/A |
| FIJI | imageJ-win64 | N/A |
| ZEISS Axiovision | Carl Zeiss AG Axiovision Rel. 4.8 | N/A |
| Others | ||
| ONETOUCH Vita glucometer | Life Scan, Milpitas, CA, USA | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napolitano, T.; Avolio, F.; Silvano, S.; Forcisi, S.; Pfeifer, A.; Vieira, A.; Navarro-Sanz, S.; Friano, M.E.; Ayachi, C.; Garrido-Utrilla, A.; et al. Gfi1 Loss Protects against Two Models of Induced Diabetes. Cells 2021, 10, 2805. https://doi.org/10.3390/cells10112805
Napolitano T, Avolio F, Silvano S, Forcisi S, Pfeifer A, Vieira A, Navarro-Sanz S, Friano ME, Ayachi C, Garrido-Utrilla A, et al. Gfi1 Loss Protects against Two Models of Induced Diabetes. Cells. 2021; 10(11):2805. https://doi.org/10.3390/cells10112805
Chicago/Turabian StyleNapolitano, Tiziana, Fabio Avolio, Serena Silvano, Sara Forcisi, Anja Pfeifer, Andhira Vieira, Sergi Navarro-Sanz, Marika Elsa Friano, Chaïma Ayachi, Anna Garrido-Utrilla, and et al. 2021. "Gfi1 Loss Protects against Two Models of Induced Diabetes" Cells 10, no. 11: 2805. https://doi.org/10.3390/cells10112805
APA StyleNapolitano, T., Avolio, F., Silvano, S., Forcisi, S., Pfeifer, A., Vieira, A., Navarro-Sanz, S., Friano, M. E., Ayachi, C., Garrido-Utrilla, A., Atlija, J., Hadzic, B., Becam, J., Sousa-De-Veiga, A., Plaisant, M. D., Balaji, S., Pisani, D. F., Mondin, M., Schmitt-Kopplin, P., ... Collombat, P. (2021). Gfi1 Loss Protects against Two Models of Induced Diabetes. Cells, 10(11), 2805. https://doi.org/10.3390/cells10112805