
Ischemia-Reperfusion Injury in a Simulated Lung Transplant Setting Differentially Regulates Transcriptomic Profiles between Human Lung Endothelial and Epithelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Reagents
2.2. Western Blotting
2.3. Flow Cytometry
2.4. Simulated IR Model for Lung Transplantation
2.5. Gene Expression
2.6. Gene Set Enrichment Analysis (GSEA)
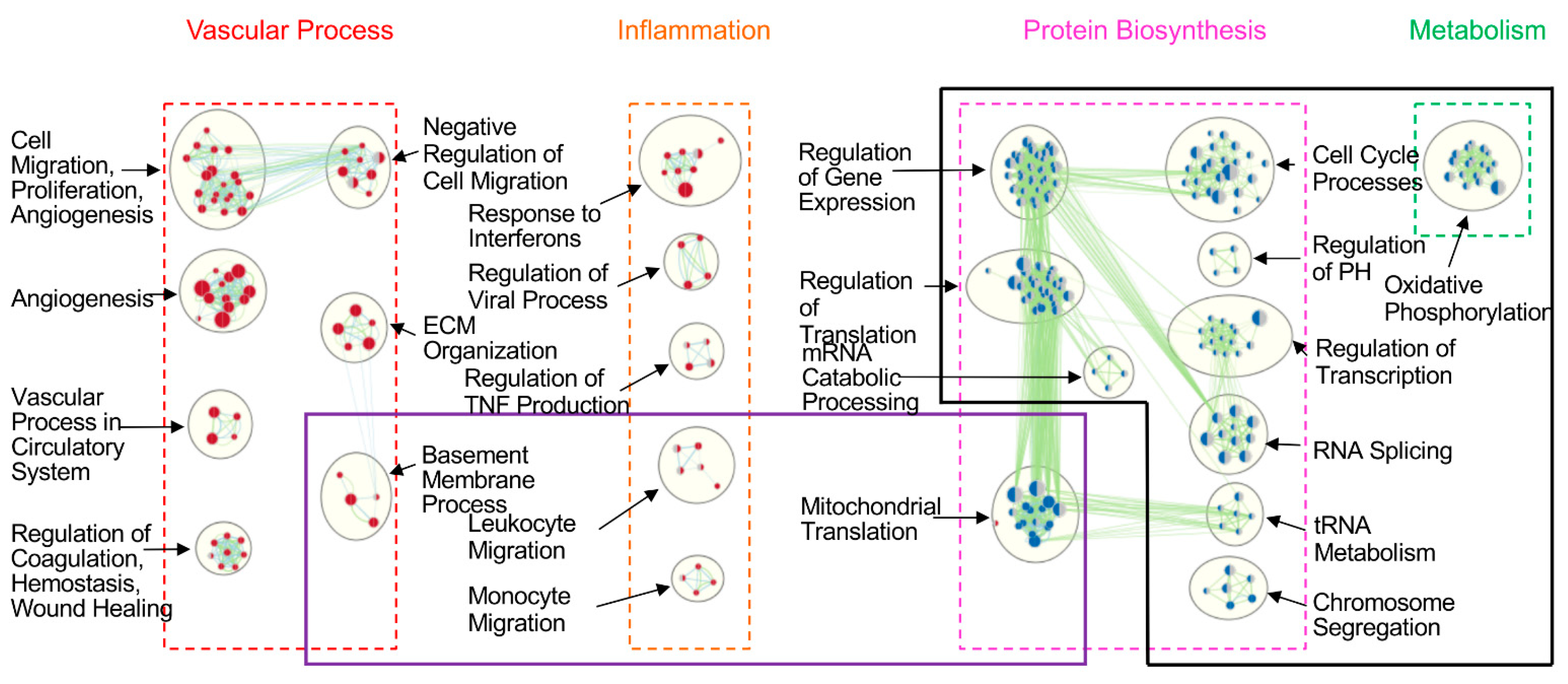
3. Results
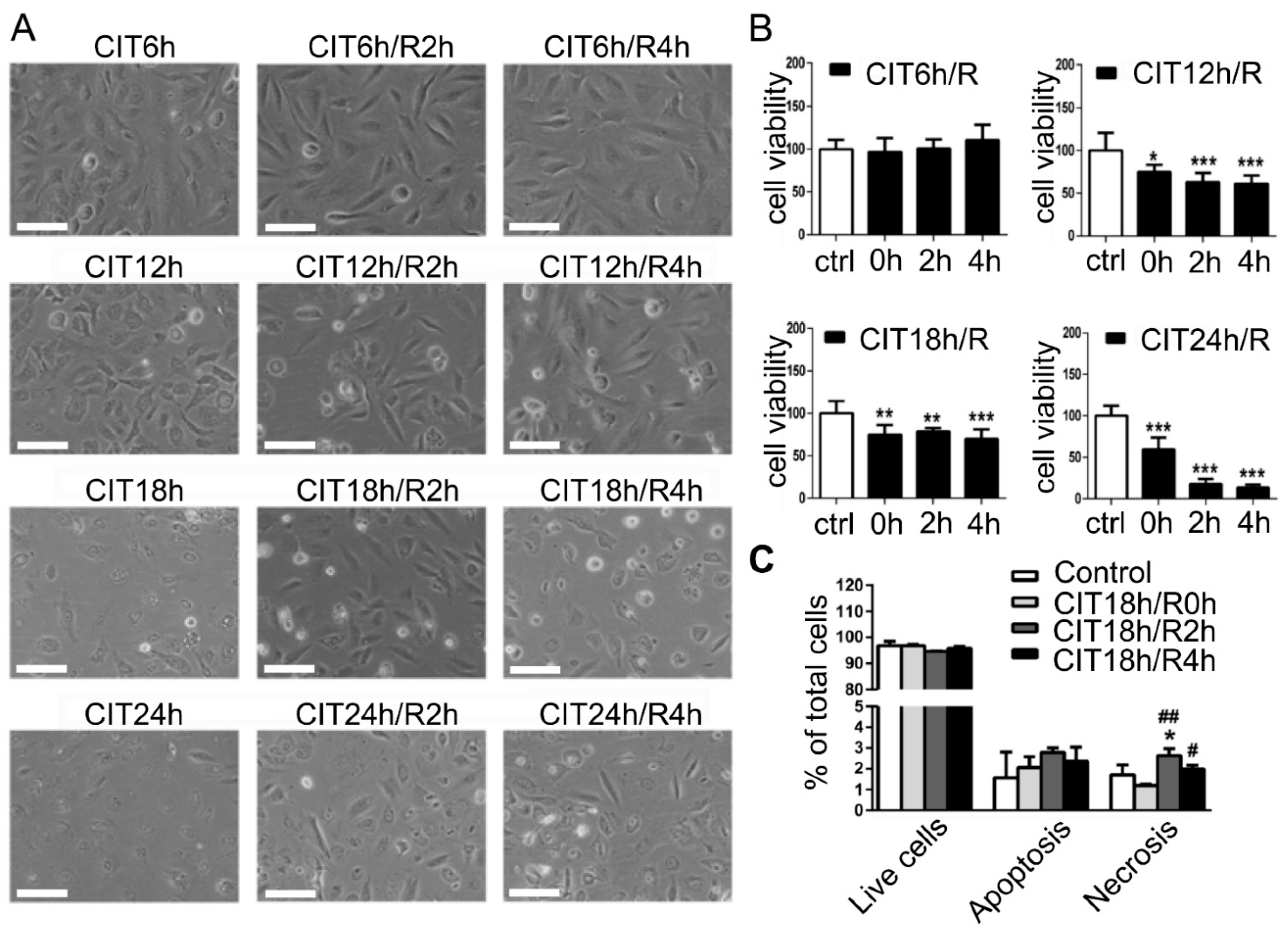
3.1. Cold Preservation and Warm Reperfusion Reduced Viability of HPMEC and BEAS-2B Cells
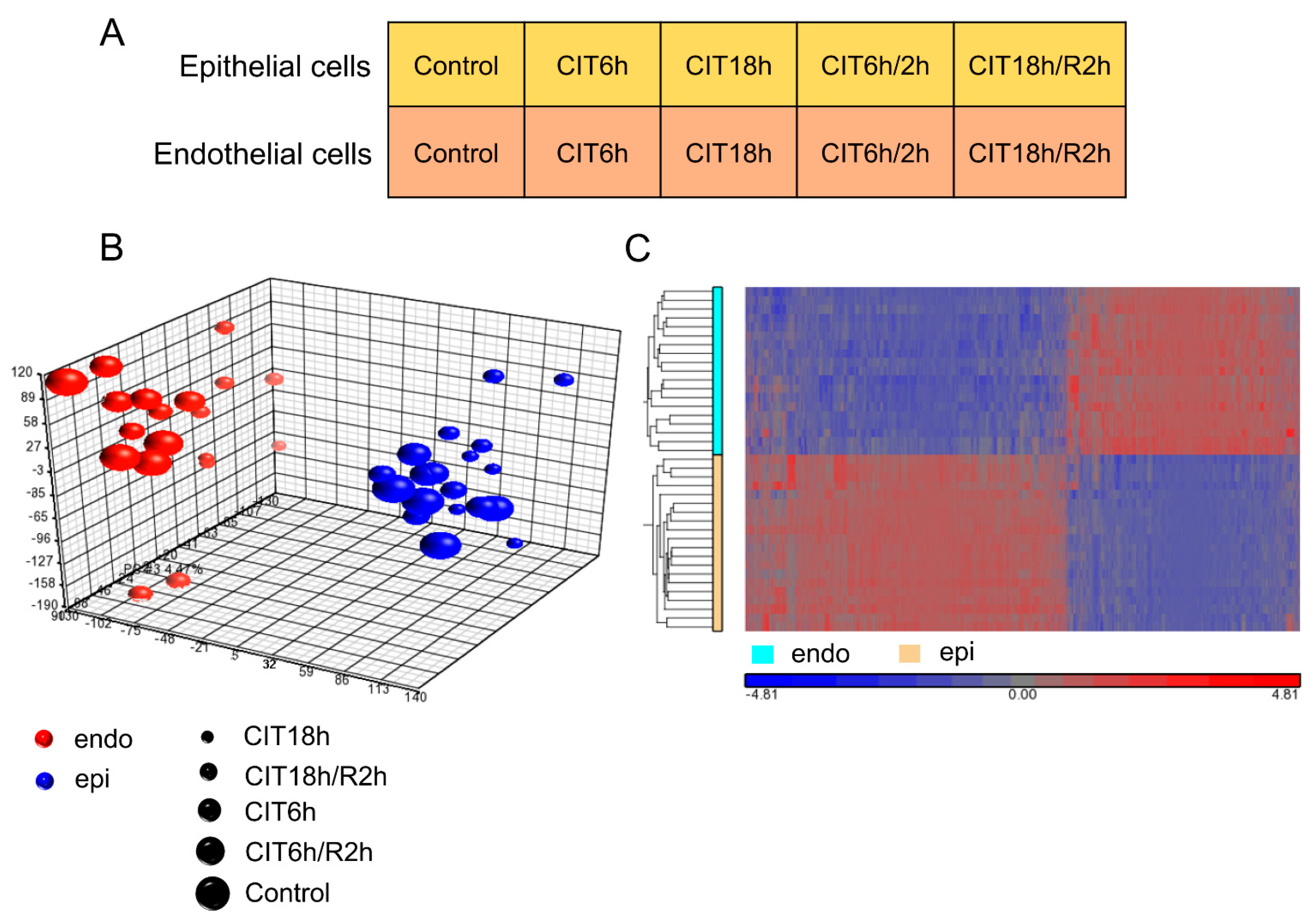
3.2. HPMEC and BEAS-2B Cells Showed Significantly Different Transcriptomic Gene Profiles
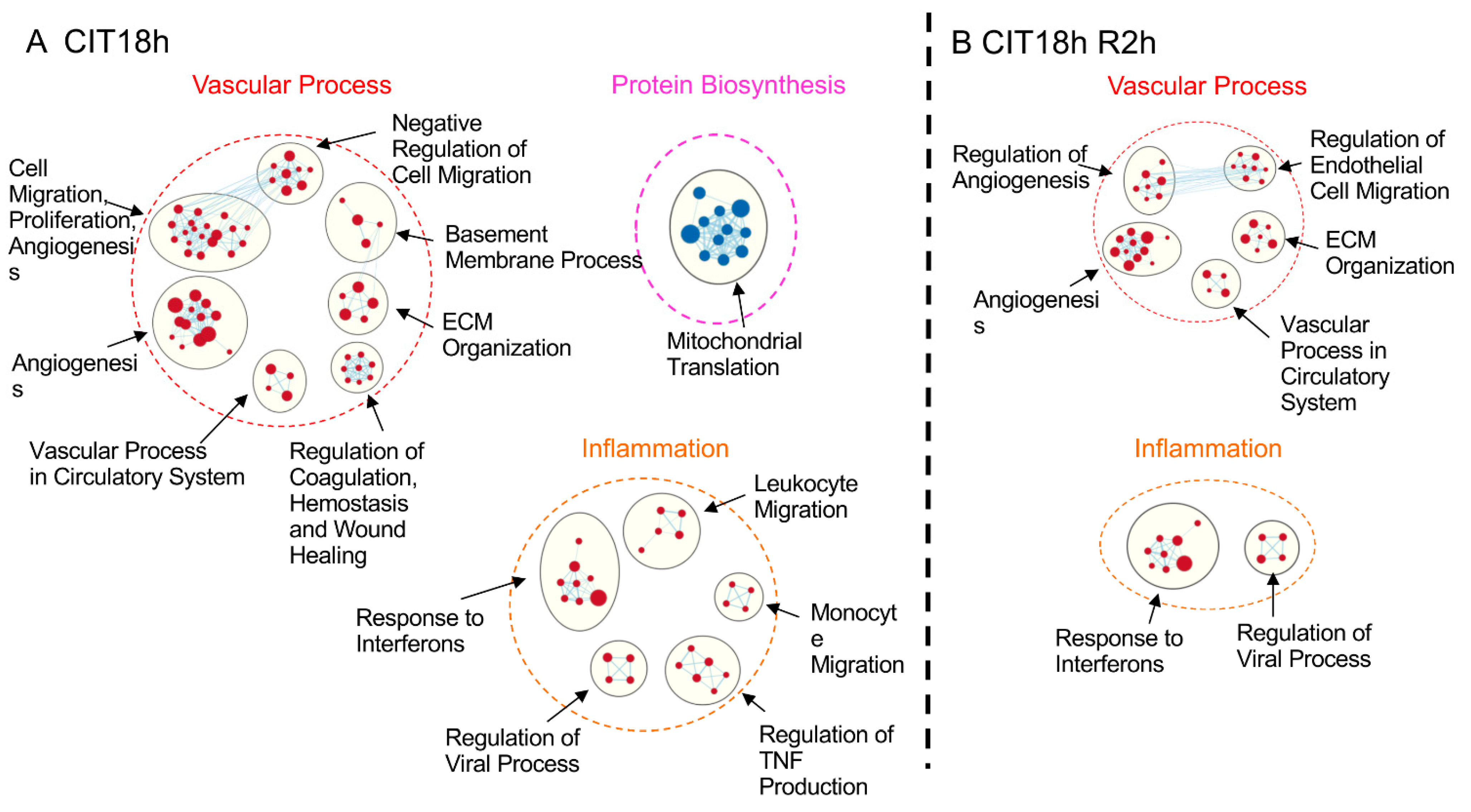
3.3. IR Differentially Affected Gene Expression in Human Pulmonary Endothelial and Epithelial Cells in a CIT Time-Dependent Manner
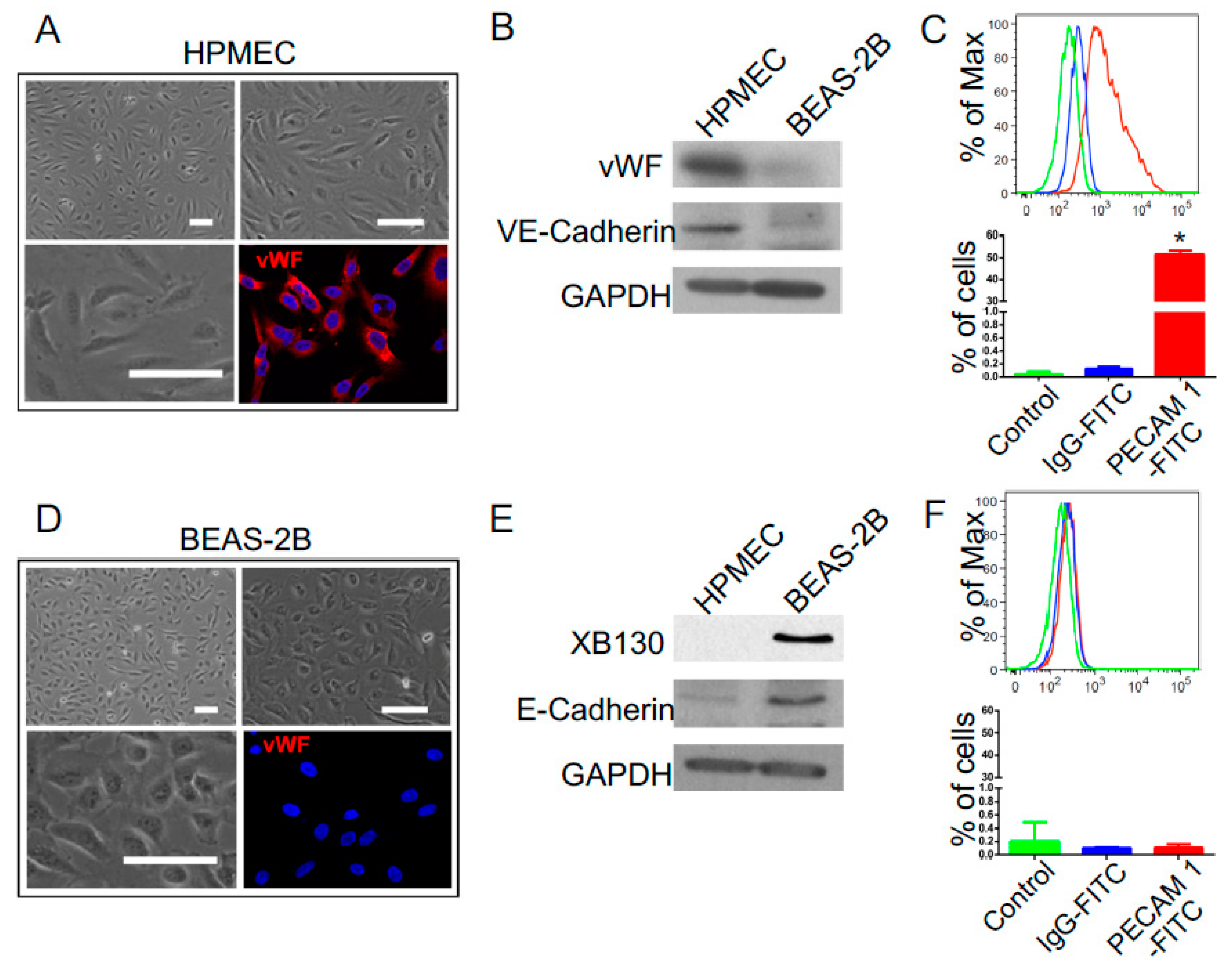
3.4. IR-Induced Loss of Phenotypic Gene Expression Characteristics of Human Lung Endothelial and Epithelial Cells
3.5. Prolonged CIT Dramatically Changed the Cell-Type Specific Signatures
4. Discussion
4.1. IR-Induced Cell Death and Gene Expression in Human Lung Endothelial and Epithelial Cells
4.2. The Phenotypic Comparison between Human Lung Endothelial and Epithelial Cells
4.3. IRI Reduced Phenotypic Characteristics of Human Lung Cells
4.4. Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wong, A.; Zamel, R.; Yeung, J.; Bader, G.D.; Dos Santos, C.C.; Bai, X.; Wang, Y.; Keshavjee, S.; Liu, M. Potential Therapeutic Targets for Lung Repair during Human Ex Vivo Lung Perfusion. Eur. Respir. J. 2020, 55, 1902222. [Google Scholar] [CrossRef]
- Andrade, C.F.; Kaneda, H.; Der, S.; Tsang, M. Toll-like Receptor and Cytokine Gene Expression in the Early Phase of Human Lung Transplantation. J. Heart Lung Transplant. 2006, 25, 1317–1323. [Google Scholar] [CrossRef]
- Sakiyama, S.; Hamilton, J.; Han, B.; Jiao, Y.; Shen-Tu, G.; de Perrot, M.; Keshavjee, S.; Liu, M. Activation of Mitogen-Activated Protein Kinases during Human Lung Transplantation. J. Heart Lung Transplant. 2005, 24, 2079–2085. [Google Scholar] [CrossRef]
- Fischer, S.; Cassivi, S.D.; Xavier, A.M.; Cardella, J.A.; Cutz, E.; Edwards, V.; Liu, M.; Keshavjee, S. Cell Death in Human Lung Transplantation: Apoptosis Induction in Human Lungs During Ischemia and After Transplantation. Ann. Surg. 2000, 231, 424–431. [Google Scholar] [CrossRef]
- Huang, L.; Yang, C.; Liu, M. Intracellular Signal Transduction Pathways as Potential Drug Targets for Ischemia-Reperfusion Injury in Lung Transplantation. J. Thorac. Dis. 2018, 10, S3965–S3969. [Google Scholar] [CrossRef] [PubMed]
- Travaglini, K.J.; Nabhan, A.N.; Penland, L.; Sinha, R.; Gillich, A.; Sit, R.V.; Chang, S.; Conley, S.D.; Mori, Y.; Seita, J.; et al. A Molecular Cell Atlas of the Human Lung from Single-Cell RNA Sequencing. Nature 2020, 587, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Cardella, J.A.; Keshavjee, S.; Mourgeon, E.; Cassivi, S.D.; Fischer, S.; Isowa, N.; Slutsky, A.; Liu, M. A Novel Cell Culture Model for Studying Ischemia-Reperfusion Injury in Lung Transplantation. J. Appl. Physiol. 2000, 89, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Zhao, J.; Zhang, Q.; Wang, Y.; Lee, D.; Bai, X.; Turrell, L.; Chen, M.; Gao, W.; Keshavjee, S.; et al. ΔV1-1 Reduces Pulmonary Ischemia Reperfusion-Induced Lung Injury by Inhibiting Necrosis and Mitochondrial Localization of PKCδ and P53. Am. J. Transplant. 2016, 16, 83–98. [Google Scholar] [CrossRef]
- Lee, D.; Zhao, J.; Yang, H.; Xu, S.; Kim, H.; Pacheco, S.; Keshavjee, S.; Liu, M. Effective Delivery of a Rationally Designed Intracellular Peptide Drug with Gold Nanoparticle–Peptide Hybrids. Nanoscale 2015, 7, 12356–12360. [Google Scholar] [CrossRef]
- Kim, H.; Zamel, R.; Bai, X.-H.; Lu, C.; Keshavjee, S.; Keshavjee, S.; Liu, M. Ischemia-Reperfusion Induces Death Receptor-Independent Necroptosis via Calpain-STAT3 Activation in a Lung Transplant Setting. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L595–L608. [Google Scholar] [CrossRef]
- Kanou, T.; Ohsumi, A.; Kim, H.; Chen, M.; Bai, X.; Guan, Z.; Hwang, D.; Cypel, M.; Keshavjee, S.; Liu, M. Inhibition of Regulated Necrosis Attenuates Receptor-Interacting Protein Kinase 1–Mediated Ischemia-Reperfusion Injury after Lung Transplantation. J. Heart Lung Transplant. 2018, 37, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhao, J.; Kim, H.; Xu, S.; Chen, M.; Bai, X.; Toba, H.; Cho, H.-R.R.; Zhang, H.; Keshavjeel, S.; et al. A1-Antitrypsin Inhibits Ischemia Reperfusion-Induced Lung Injury by Reducing Inflammatory Response and Cell Death. J. Heart Lung Transplant. 2014, 33, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Iskender, I.; Sakamoto, J.; Nakajima, D.; Lin, H.; Chen, M.; Kim, H.; Guan, Z.; Del Sorbo, L.; Hwang, D.; Waddell, T.K.; et al. Human A1-Antitrypsin Improves Early Post-Transplant Lung Function: Pre-Clinical Studies in a Pig Lung Transplant Model. J. Heart Lung Transplant. 2016, 35, 913–921. [Google Scholar] [CrossRef]
- Gao, X.; Wei, L.; Mingyao, L. Ex Vivo Lung Perfusion: Scientific Research and Clinical Application. Pract. J. Organ Transplant. 2017, 5, 177–187. [Google Scholar]
- Lin, H.; Chen, M.; Tian, F.; Tikkanen, J.; Ding, L.; Andrew Cheung, H.Y.; Nakajima, D.; Wang, Z.; Mariscal, A.; Hwang, D.; et al. α 1-Anti-Trypsin Improves Function of Porcine Donor Lungs during Ex-Vivo Lung Perfusion. J. Heart Lung Transplant. 2018, 37, 656–666. [Google Scholar] [CrossRef]
- Mariscal, A.; Nykanen, A.; Tikkanen, J.; Ali, A.; Soltanieh, S.; Duong, A.; Galasso, M.; Juvet, S.; Martinu, T.; Cypel, M.; et al. Alpha 1 Antitrypsin Treatment during Human Ex Vivo Lung Perfusion Improves Lung Function by Protecting Lung Endothelium. J. Heart Lung Transplant. 2020, 39, S71–S72. [Google Scholar] [CrossRef]
- Berger, M.; Liu, M.; Uknis, M.E.; Koulmanda, M. Alpha-1-Antitrypsin in Cell and Organ Transplantation. Am. J. Transplant. 2018, 18, 1589–1595. [Google Scholar] [CrossRef]
- Casiraghi, M.; Tatreau, J.R.; Abano, J.B.; Blackwell, J.W.; Watson, L.; Burridge, K.; Randell, S.H.; Egan, T.M. In Vitro Modeling of Nonhypoxic Cold Ischemia-Reperfusion Simulating Lung Transplantation. J. Thorac. Cardiovasc. Surg. 2009, 138, 760–767. [Google Scholar] [CrossRef]
- Krump-Konvalinkova, V.; Bittinger, F.; Unger, R.E.; Peters, K.; Lehr, H.-A.; Kirkpatrick, C.J. Generation of Human Pulmonary Microvascular Endothelial Cell Lines. Lab. Investig. 2001, 81, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P.; Beazer-barclay, Y.D.; Antonellis, K.J.; Speed, T.P. Exploration, Normalization, and Summaries of High Density Oligonucleotide Array Probe Level Data. Biostatistics 2018, 4, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, B.S.; Irizarry, R.A. A Framework for Oligonucleotide Microarray Preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway Enrichment Analysis and Visualization of Omics Data Using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]
- Wu, Q.; Nadesalingam, J.; Moodley, S.; Bai, X.; Liu, M. XB130 Translocation to Microfilamentous Structures Mediates NNK-Induced Migration of Human Bronchial Epithelial Cells. Oncotarget 2015, 6, 18050–18065. [Google Scholar] [CrossRef][Green Version]
- Moodley, S.; Bai, X.H.; Kapus, A.; Yang, B.; Liu, M. XB130/Tks5 Scaffold Protein Interaction Regulates Src-Mediated Cell Proliferation and Survival. Mol. Biol. Cell 2015, 26, 4492–4502. [Google Scholar] [CrossRef]
- Moodley, S.; Derouet, M.; Bai, X.H.; Xu, F.; Kapus, A.; Yang, B.B.; Liu, M. Stimulus-Dependent Dissociation between XB130 and Tks5 Scaffold Proteins Promotes Airway Epithelial Cell Migration. Oncotarget 2016, 7, 76437–76452. [Google Scholar] [CrossRef]
- Fischer, S.; Maclean, A.A.; Liu, M.; Cardella, J.A.; Slutsky, A.S.; Suga, M.; Moreira, J.F.; Keshavjee, S. Dynamic Changes in Apoptotic and Necrotic Cell Death Correlate with Severity of Ischemia-Reperfusion Injury in Lung Transplantation. Am. J. Respir. Crit. Care Med. 2000, 162, 1932–1939. [Google Scholar] [CrossRef]
- Novick, R.J.; Gehman, K.E.; Ali, I.S.; Lee, J. Lung Preservation: The Importance of Endothelial and Alveolar Type II Cell Integrity. Ann. Thorac. Surg. 1996, 62, 302–314. [Google Scholar] [CrossRef]
- Gebb, S.; Stevens, T. On Lung Endothelial Cell Heterogeneity. Microvasc. Res. 2004, 68, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, L.; Wagner, E.M. Angiogenesis in the Lung. J. Physiol. 2019, 597, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Summer, R. Cellular Metabolism in Lung Health and Disease. Annu. Rev. Physiol. 2019, 81, 403–428. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Bernard, A. Lung Epithelium–Specific Proteins. Am. J. Respir. Crit. Care Med. 1999, 159, 646–678. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Tanswell, A.K.; Post, M. Mechanical Force-Induced Signal Transduction in Lung Cells. Am. J. Physiol. Cell. Mol. Physiol. 1999, 277, L667–L683. [Google Scholar] [CrossRef] [PubMed]
- Huh, D.; Matthews, B.D.; Mammoto, A.; Montoya-Zavala, M.; Hsin, H.Y.; Ingber, D.E. Reconstituting Organ-Level Lung Functions on a Chip. Science 2010, 328, 1662–1668. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saren, G.; Wong, A.; Lu, Y.-B.; Baciu, C.; Zhou, W.; Zamel, R.; Soltanieh, S.; Sugihara, J.; Liu, M. Ischemia-Reperfusion Injury in a Simulated Lung Transplant Setting Differentially Regulates Transcriptomic Profiles between Human Lung Endothelial and Epithelial Cells. Cells 2021, 10, 2713. https://doi.org/10.3390/cells10102713
Saren G, Wong A, Lu Y-B, Baciu C, Zhou W, Zamel R, Soltanieh S, Sugihara J, Liu M. Ischemia-Reperfusion Injury in a Simulated Lung Transplant Setting Differentially Regulates Transcriptomic Profiles between Human Lung Endothelial and Epithelial Cells. Cells. 2021; 10(10):2713. https://doi.org/10.3390/cells10102713
Chicago/Turabian StyleSaren, Gaowa, Aaron Wong, Yun-Bi Lu, Cristina Baciu, Wenyong Zhou, Ricardo Zamel, Sahar Soltanieh, Junichi Sugihara, and Mingyao Liu. 2021. "Ischemia-Reperfusion Injury in a Simulated Lung Transplant Setting Differentially Regulates Transcriptomic Profiles between Human Lung Endothelial and Epithelial Cells" Cells 10, no. 10: 2713. https://doi.org/10.3390/cells10102713
APA StyleSaren, G., Wong, A., Lu, Y.-B., Baciu, C., Zhou, W., Zamel, R., Soltanieh, S., Sugihara, J., & Liu, M. (2021). Ischemia-Reperfusion Injury in a Simulated Lung Transplant Setting Differentially Regulates Transcriptomic Profiles between Human Lung Endothelial and Epithelial Cells. Cells, 10(10), 2713. https://doi.org/10.3390/cells10102713