Kinetics of DNA Repair in Vicia faba Meristem Regeneration Following Replication Stress

 ,
,  ,
, 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
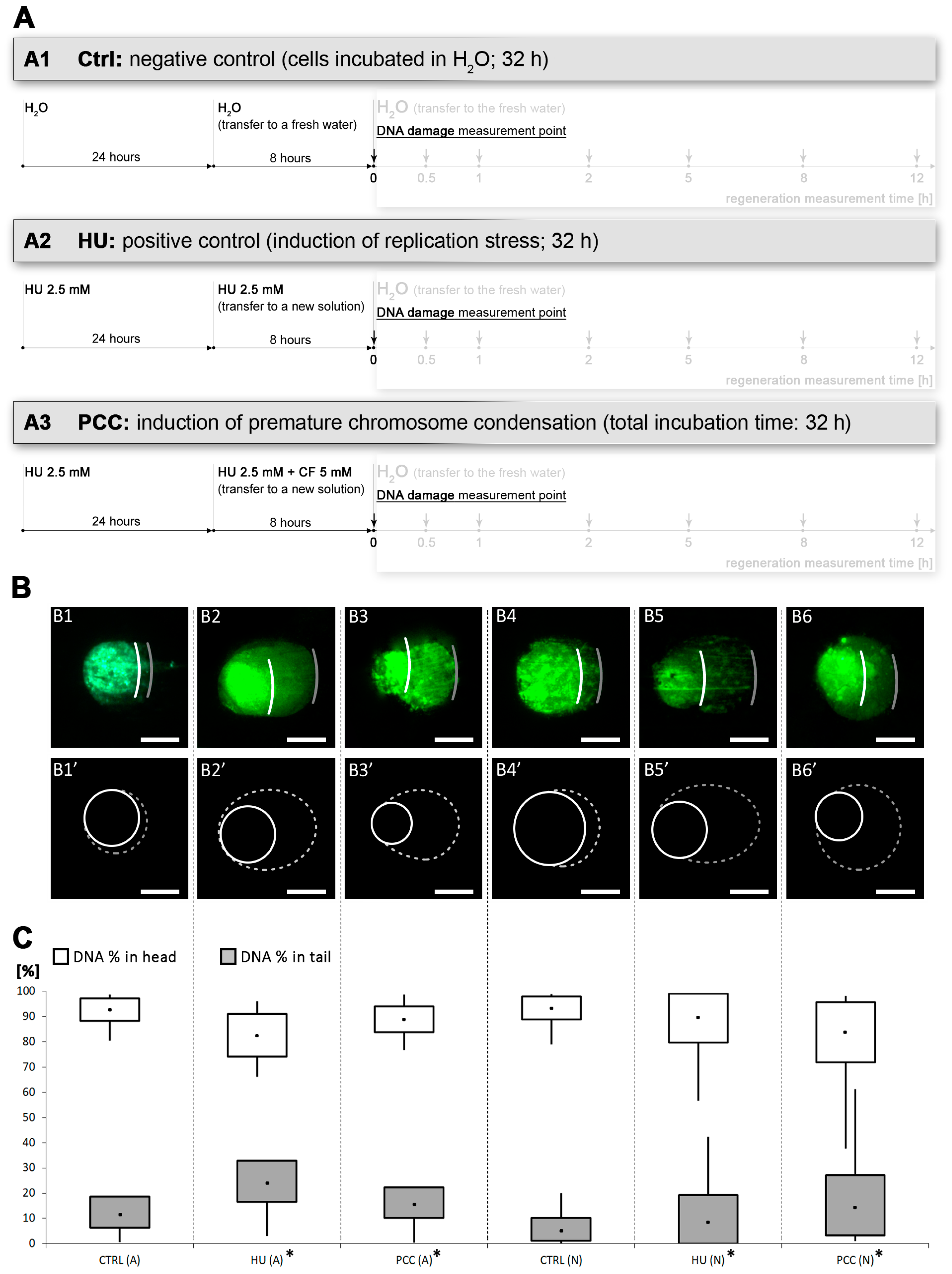
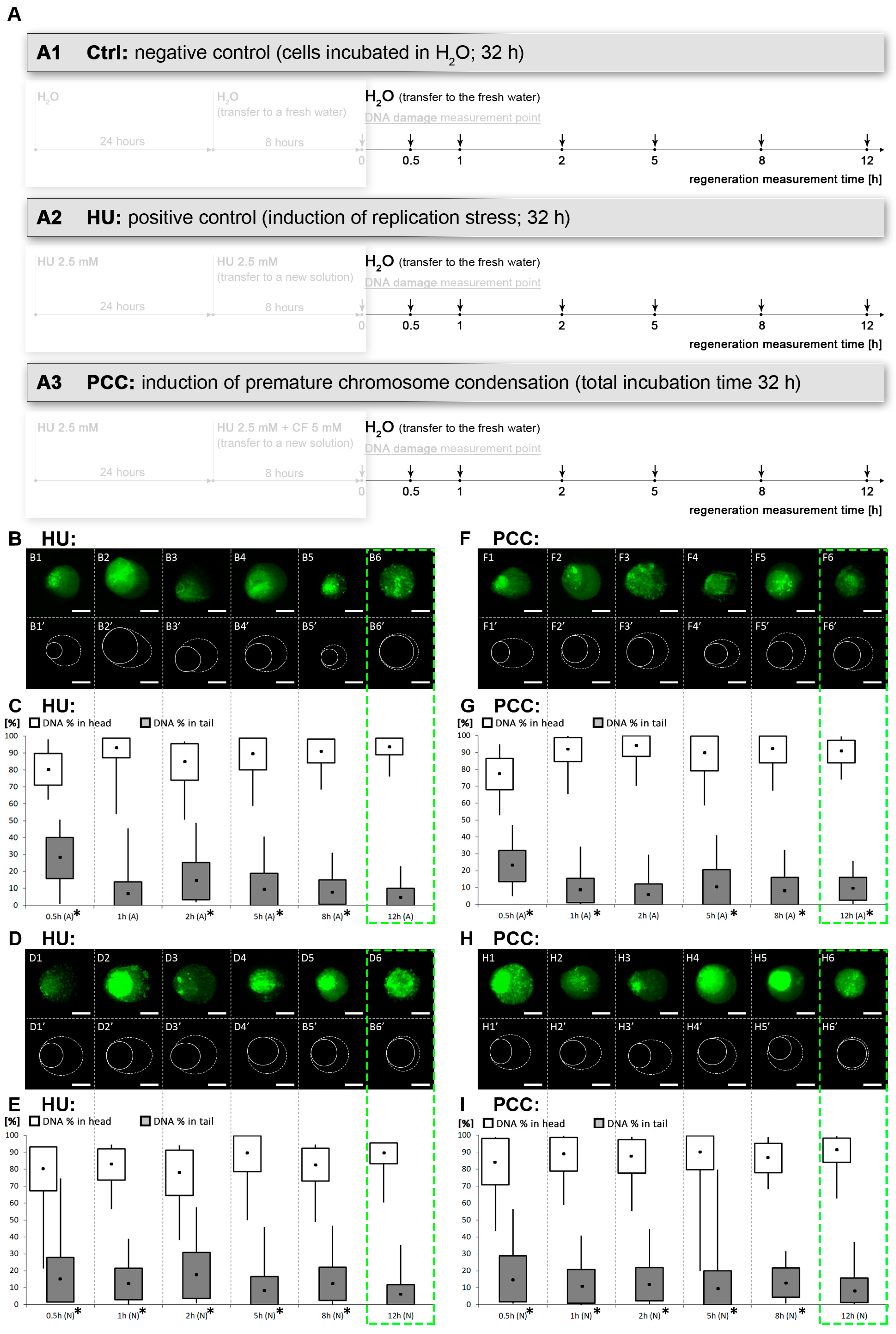
2.1. Growth Conditions of Plant Material, Induction of Replication Stress, PCC and Regeneration Period
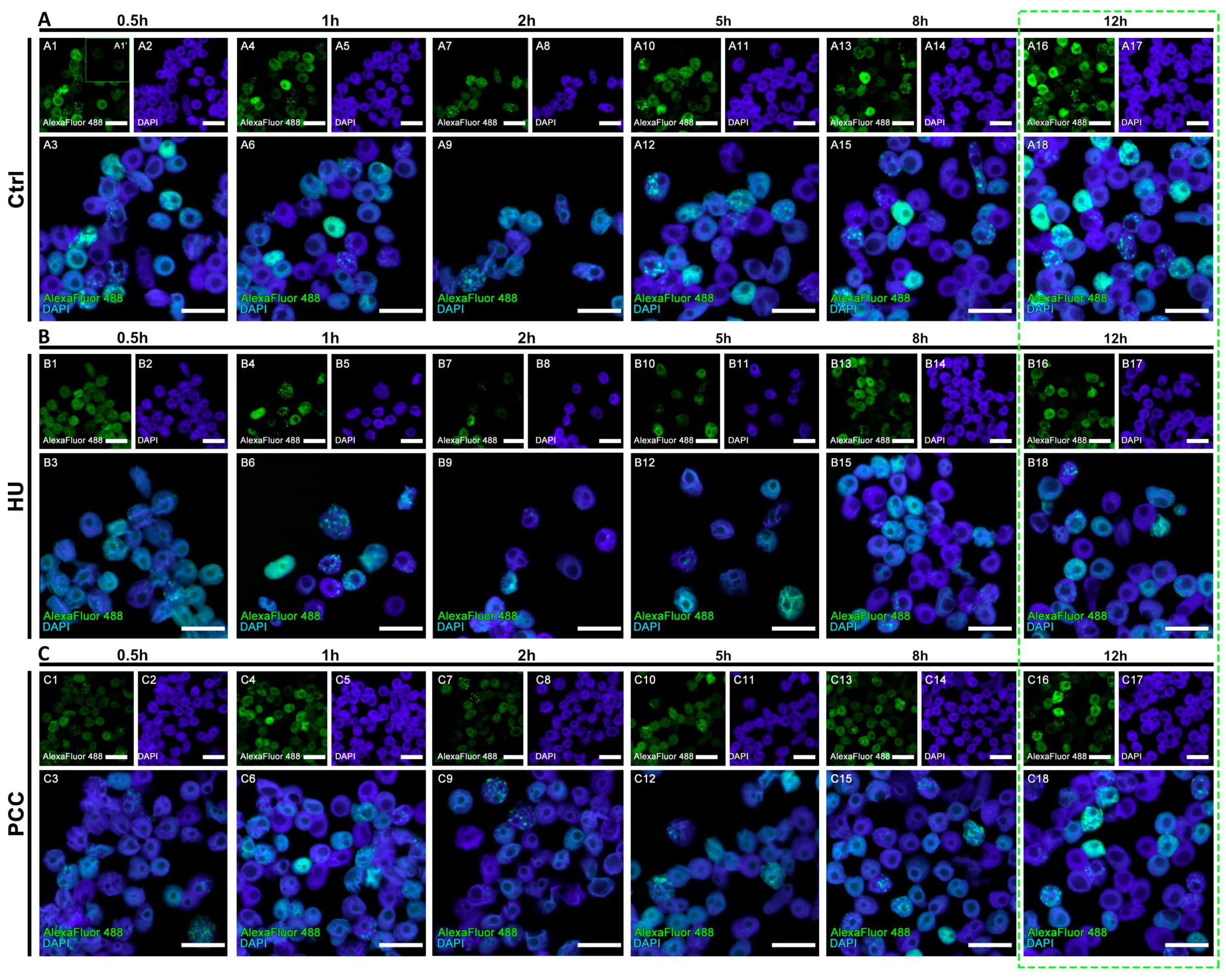
2.2. Comet Assay
2.3. Flow Cytometric Analysis
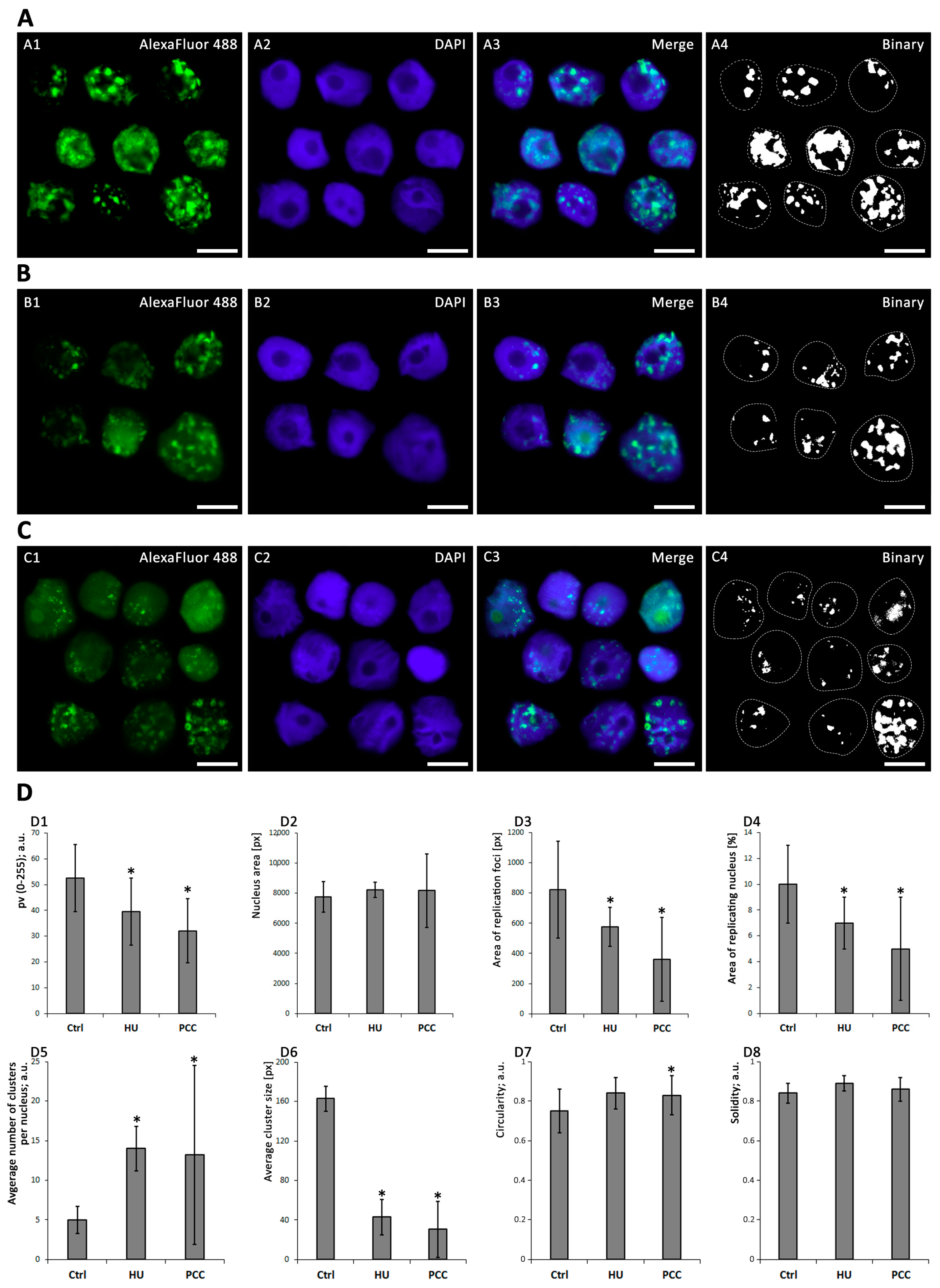
2.4. Heterochromatin Clusters Analysis
2.5. Statistical Analysis and Image Processing
3. Results
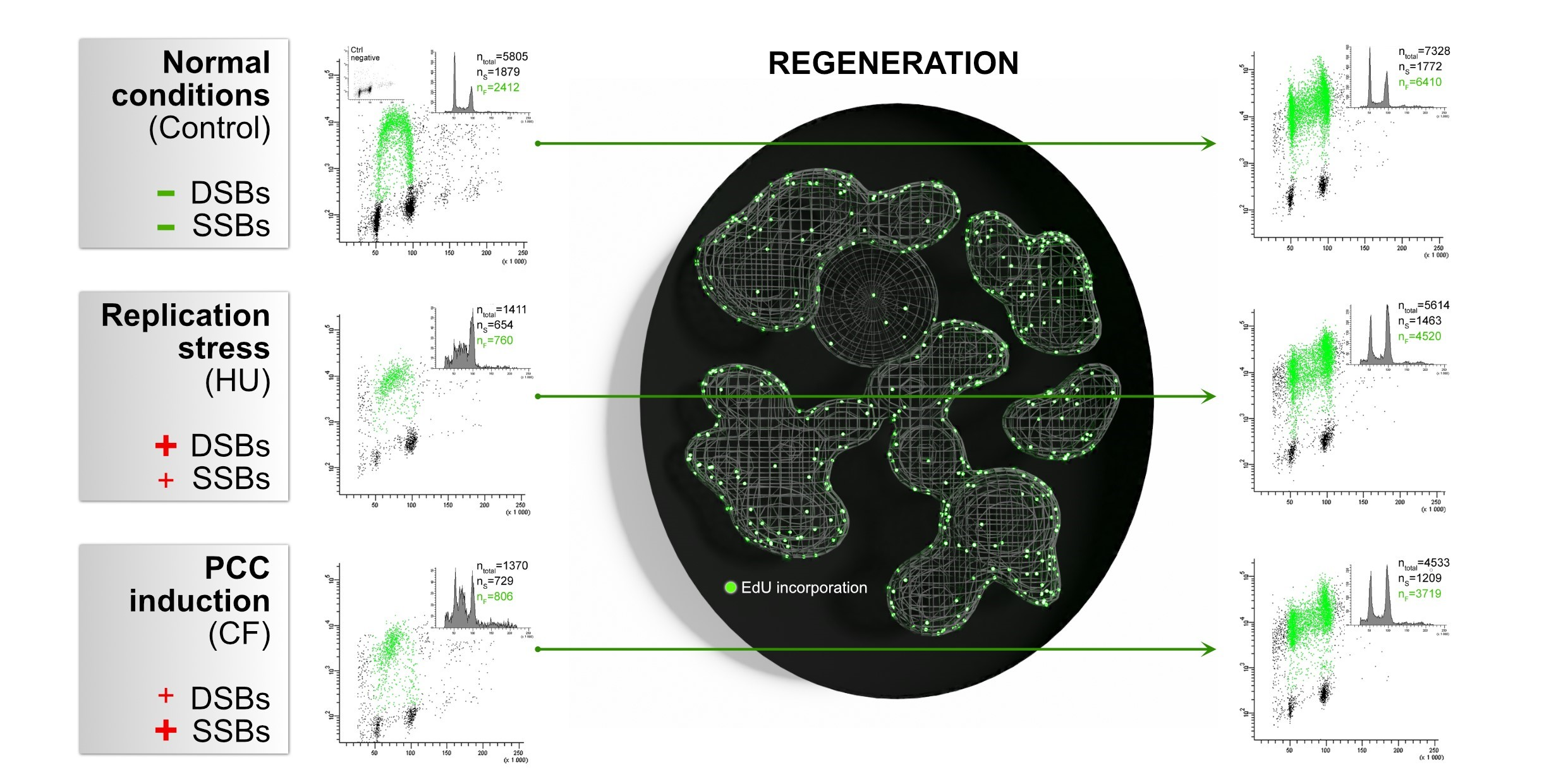
3.1. Replication Restarts and Bursts just after the Release from the Replication Block or PCC
3.2. The Profiles of Heterochromatin (HC) Replication Clusters Are Altered in the PCC-Induced Cells
3.3. The Amount of DNA Damage Sites Are Temporarily Increased during the Repair Process, the Regeneration from PCC Induction Is Slower than the Regeneration from the Replication Stress
3.4. The Starting Point of the Regeneration Is Vital for the Whole Regeneration Process
4. Discussion
4.1. Starting Point of Regeneration—DNA Damage Repair, Replication Restart and Cluster Constitution
4.2. Regeneration Period
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
List of Abbreviations
| ATM | ataxia telangiectasia-mutated kinase |
| ATR | ataxia telangiectasia and Rad3-related kinase |
| CF | caffeine |
| dNTPs | deoxyribonucleoside triphosphates |
| DSB | double-strand break |
| EC | euchromatin |
| HC | heterochromatin |
| HR | homologous recombination |
| HU | hydroxyurea |
| PCC | premature chromosome condensation |
| PCD | programmed cell death |
| PCP II | principal control point II |
| SSB | single-strand break |
References
- Ovejero, S.; Bueno, A.; Sacristán, M.P. Working on genome stability: From the S-phase to mitosis. Genes 2020, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Winnicki, K. ATM/ATR-dependent Tyr15 phosphorylation of cyclin-dependent kinases in response to hydroxyurea in Vicia faba root meristem cells. Protoplasma 2013, 250, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Hübner, B.; Strickfaden, H.; Müller, S.; Cremer, M.; Cremer, T. Chromosome shattering: A mitotic catastrophe due to chromosome condensation failure. Eur. Biophys. J. 2009, 38, 729–747. [Google Scholar]
- Rybaczek, D.; Kowalewicz-Kulbat, M. Premature chromosome condensation induced by caffeine, 2-aminopurine, staurosporine and sodium metavanadate in S-phase arrested HeLa cells is associated with a decrease in Chk1 phosphorylation, formation of phospho-H2AX and minor cytoskeletal rearrangements. Histochem. Cell Biol. 2011, 135, 263–280. [Google Scholar] [PubMed]
- Masamsetti, V.P.; Low, R.R.J.; Mak, K.S.; O’Connor, A.; Riffkin, C.D.; Lamm, N.; Crabbe, L.; Karlseder, J.; Huang, D.C.S.; Hayashi, M.T.; et al. Replication stress induces mitotic death through parallel pathways regulated by WAPL and telomere deprotection. Nat. Commun. 2019, 10, 4224–4239. [Google Scholar] [CrossRef]
- Zeman, M.K.; Cimprich, K.A. Causes and consequences of replication stress. Nat. Cell Biol. 2014, 16, 2–9. [Google Scholar] [CrossRef]
- Moiseeva, T.N.; Yin, Y.; Calderon, M.J.; Qian, C.; Schamus-Haynes, S.; Sugitani, N.; Osmanbeyoglu, H.U.; Rothenberg, E.; Watkins, S.C.; Bakkenist, C.J. An ATR and CHK1 kinase signaling mechanism that limits origin firing during unperturbed DNA replication. Proc. Nat. Acad. Sci. USA 2019, 116, 13374–13383. [Google Scholar] [CrossRef]
- Moiseeva, T.N.; Qian, C.; Sugitani, N.; Osmanbeyoglu, H.U.; Bakkenist, C.J. WEE1 kinase inhibitor AZD1775 induces CDK1 kinase-dependent origin firing in unperturbed G1- and S-phase cells. Proc. Nat. Acad. Sci. USA 2019, 116, 23891–23893. [Google Scholar] [CrossRef]
- Barnum, K.J.; O’Connell, M.J. Cell cycle regulation by checkpoints. Methods Mol. Biol. 2014, 1170, 29–40. [Google Scholar]
- Elledge, S.J. Cell cycle checkpoints: Preventing an identity crisis. Science 1996, 274, 1664–1672. [Google Scholar] [CrossRef]
- Sarkaria, J.N.; Busby, E.C.; Tibbetts, R.S.; Roos, P.; Taya, Y.; Karnitz, L.M.; Abraham, R.T. Inhibition of ATM and ATR kinase activities by the radiosensitizing agent, caffeine. Cancer Res. 1999, 59, 4375–4382. [Google Scholar]
- De Veylder, L.; Joubès, J.; Inzé, D. Plant cell cycle transitions. Curr. Opin. Plant Biol. 2003, 6, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.H.; Liu, Q.; Elledge, S.J.; Rosen, J.M. Chk1 is haploinsufficient for multiple functions critical to tumor suppression. Cancer Cell 2004, 6, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.J.; Baltimore, D. ATR disruption lead to chromosomal fragmentation and early embryonic lethality. Genes Dev. 2000, 14, 397–402. [Google Scholar] [PubMed]
- Offenbacher, A.R.; Barry, B.A. A proton wire mediates proton coupled electron transfer from hydroxyurea and other hydroxamic acids to tyrosyl radical in class Ia ribonucleotide reductase. J. Phys. Chem. B 2020, 124, 345–354. [Google Scholar] [CrossRef]
- Ünal, E.; Arbel-Eden, A.; Satter, U.; Shroff, R.; Lichten, M.; Haber, J.E.; Koshland, D. DNA damage response pathway uses histone modification to assemble a double-strand break-specific cohesin domain. Mol. Cell 2004, 16, 991–1002. [Google Scholar] [CrossRef]
- Rybaczek, D.; Musiałek, M.W.; Balcerczyk, A. Caffeine-induced premature chromosome condensation results in the apoptosis-like programmed cell death in root meristems of Vicia faba. PLoS ONE 2015, 10, e0142307. [Google Scholar] [CrossRef]
- Rybaczek, D.; Bodys, A.; Maszewski, J. H2AX foci in late S/G2- and M-phase cells after hydroxyurea- and aphidicolin-induced DNA replication stress in Vicia. Histochem. Cell Biol. 2007, 128, 227–241. [Google Scholar] [CrossRef]
- Rybaczek, D.; Maszewski, J. Phosphorylation of H2AX histones in response to double-strand breaks and induction of premature chromatin condensation in hydroxyurea-treated root meristem cells of Raphanus sativus, Vicia faba, and Allium porrum. Protoplasma 2007, 230, 31–39. [Google Scholar] [CrossRef]
- Ercilla, A.; Feu, S.; Aranda, S.; Llopis, A.; Brynjólfsdóttir, S.H.; Sørensen, C.S.; Toledo, L.I.; Agell, N. Acute hydroxyurea-induced replication blockade results in replisome components disengagement from nascent DNA without causing fork collapse. Cell. Mol. Life Sci. 2020, 77, 735–749. [Google Scholar] [CrossRef]
- Moser, B.A.; Brondello, J.-M.; Baber-Furnari, B.; Russell, P. Mechanism of caffeine-induced checkpoint override in fission yeast. Mol. Cell. Biol. 2000, 20, 4288–4294. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, M.E.; Facca, C.; Fatmi, Z.; Baïlle, D.; Bénakli, S.; Vernis, L. DNA replication inhibitor hydroxyurea alters Fe-S centers by producing reactive oxygen species in vivo. Sci. Rep. 2016, 6, 29361–29372. [Google Scholar] [CrossRef]
- Nozaki, T.; Imari, R.; Tanbo, M.; Nagashima, R.; Tamura, S.; Tani, T.; Joti, Y.; Tomita, M.; Hibino, K.; Kanemaki, M.T.; et al. Dynamic organization of chromatin domains revealed by super-resolution live-cell imaging. Mol. Cell 2017, 67, 282–293.e7. [Google Scholar] [CrossRef] [PubMed]
- Allshire, R.C.; Hiten, D.M. Ten principles of heterochromatin formation and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Reinberg, D. Facultative heterochromatin: Is there a distinctive molecular signature? Mol. Cell 2007, 28, 1–13. [Google Scholar] [CrossRef]
- Al-Suhaibani, N.A. Influence of early water deficit on seed yield and quantity of faba bean under arid environment of Saudi Arabia. J. Agric. Environ. Sci. 2009, 5, 649–654. [Google Scholar]
- Almadini, A.M. Biological and mineral nitrogen supply impacts on salinity response of faba bean (Vicia faba L.). Eur. J. Sci. Res. 2011, 49, 187–199. [Google Scholar]
- Vioque, J.; Alaiz, M.; Girón-Calle, J. Nutritional and functional properties of Vicia faba protein isolated and related fractions. Food Chem. 2012, 132, 67–72. [Google Scholar] [CrossRef]
- Ammar, M.H.; Anwar, F.; El-Harty, E.H.; Migdadi, H.M.; Abdel-Khalik, S.M.; Al-Faifi, S.A.; Farooq, M.; Alghamdi, S.S. Physiological and yield responses of faba bean (Vicia faba L.) to drought stress in managed and open field environments. J. Agron. Crop Sci. 2015, 201, 280–287. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought stress in gain legumes during reproduction and gain filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Hussain, M.; Barthakur, S.; Paul, S.; Bharadwaj, N.; Migdadi, H.M.; Alghamdi, S.S.; Siddique, K.H.M. Effect, tolerance mechanisms and management of salt stress in grain legumes. Plant Physiol. Biochem. 2017, 118, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Saban, N.; Bujak, M. Hydroxyurea and hydroxamic acid derivatives as antitumor drugs. Cancer Chemother. Pharmacol. 2009, 64, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.; Marques, F.; Fokt, H.; Oliveira, R.; Johansson, B. Measuring oxidative DNA damage and DNA repair using the yeast comet assay. Yeast 2011, 28, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Klaude, M.; Eriksson, S.; Nygren, J.; Ahnström, G. The comet assay: Mechanisms and technical considerations. Mutat. Res. 1996, 363, 89–96. [Google Scholar] [CrossRef]
- Potocki, L.; Lewinska, A.; Klukowska-Rötzler, J.; Bielak-Zmijewska, A.; Grabowska, W.; Rzeszutek, I.; Kaminska, P.; Roga, E.; Bugno-Poniewierska, M.; Slota, E.; et al. Sarcoid-derived fibroblasts: Links between genomic instability, energy metabolism and senescence. Biochimie 2014, 97, 163–172. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529–554. [Google Scholar] [CrossRef]
- Wear, E.E.; Concia, L.; Brooks, A.M.; Markham, E.A.; Lee, T.-J.; Allen, G.C.; Thompson, W.F.; Hanley-Bowdoin, L. Isolation of plant nuclei at defined cell cycle stages using EdU labeling and flow cytometry. In Plant Cell Division; Part of the Methods in Molecular Biology Book Series; Humana Press: New York, NY, USA, 2012; Volume 1370, pp. 69–86. [Google Scholar]
- Vrána, J.; Šimková, H.; Kubaláková, M.; Číhalíková, J.; Doležel, J. Flow cytometric chromosome sorting in plants: The next generation. Methods 2012, 57, 331–337. [Google Scholar] [CrossRef]
- Doležel, J.; Sgorbati, S.; Lucretti, S. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol. Plantarum. 1992, 85, 625–631. [Google Scholar] [CrossRef]
- Gotoh, E. G2 premature chromosome condensation/chromosome aberration assay: Drug-induced premature chromosome condensation (PCC) protocols and cytogenetic approaches in mitotic chromosome and interphase chromatin for radiation biology. Methods Mol. Biol. 2019, 1984, 47–60. [Google Scholar]
- Nghiem, P.; Park, P.K.; Kim, Y.; Vaziri, C.; Schreiber, S.L. ATR inhibition selectively sensitizes G1 checkpoint-deficient cells to lethal premature chromatin condensation. Proc. Nat. Acad. Sci. USA 2002, 98, 9092–9097. [Google Scholar] [CrossRef]
- Donaldson, A.D.; Blow, J.J. The regulation of replication origin activation. Curr. Opin. Genet. Dev. 1999, 9, 62–68. [Google Scholar] [CrossRef]
- Musiałek, M.W.; Rybaczek, D. Behavior of replication origins in Eukaryota—Spatio-temporal dynamics of licensing and firing. Cell Cycle 2015, 14, 2251–2264. [Google Scholar] [CrossRef] [PubMed]
- Mazurczyk, M.; Rybaczek, D. Replication and re-replication: Different implications of the same mechanism. Biochimie 2015, 108, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Fortuny, A.; Polo, S.E. The response to DNA damage in heterochromatin domains. Chromosoma 2018, 127, 291–300. [Google Scholar] [CrossRef]
- Chiolo, I.; Minoda, A.; Colmenares, S.U.; Polyzos, A.; Costes, S.V.; Karpen, G.H. Double-strand breaks in heterochromatin move outside of a dynamic HP1a domain to complete recombinational repair. Cell 2011, 144, 732–744. [Google Scholar] [CrossRef]
- Meir, A.; Kong, M.; Xue, C.; Green, E.C. DNA curtains shed light on complex molecular systems during homologous recombination. J. Vis. Exp. 2020, 160, e61320. [Google Scholar] [CrossRef]
- Bétermier, M.; Bertrand, P.; Lopez, B.S. Is non-homologous end-joining really an inherently error-prone process? PLoS Genet. 2014, 10, e1004086. [Google Scholar] [CrossRef]
- Kramara, J.; Osia, B.; Malkova, A. Break induced replication: The where, the why, and the how. Trends Genet. 2018, 34, 518–531. [Google Scholar] [CrossRef]
- Balcerczyk, A.; Rybaczek, D.; Wojtala, M.; Pirola, L.; Okabe, J.; El-Osta, A. Pharmacological inhibition of arginine and lysine methyltransferases induces nuclear abnormalities and suppresses angiogenesis in human endothelial cells. Biochem. Pharmacol. 2016, 121, 18–32. [Google Scholar] [CrossRef]
- Wojtala, M.; Dąbek, A.; Rybaczek, D.; Śliwińska, A.; Świderska, E.; Słapek, K.; El-Osta, A.; Balcerczyk, A. Silencing lysine-specific histone demethylase 1 (LSD1) causes increased HP1-positive chromatin, stimulation of DNA repair processes, and dysregulation of proliferation by Chk1 phosphorylation in human endothelial cells. Cells 2019, 8, 1212. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rybaczek, D.; Musiałek, M.W.; Vrána, J.; Petrovská, B.; Pikus, E.G.; Doležel, J. Kinetics of DNA Repair in Vicia faba Meristem Regeneration Following Replication Stress. Cells 2021, 10, 88. https://doi.org/10.3390/cells10010088
Rybaczek D, Musiałek MW, Vrána J, Petrovská B, Pikus EG, Doležel J. Kinetics of DNA Repair in Vicia faba Meristem Regeneration Following Replication Stress. Cells. 2021; 10(1):88. https://doi.org/10.3390/cells10010088
Chicago/Turabian StyleRybaczek, Dorota, Marcelina W. Musiałek, Jan Vrána, Beáta Petrovská, Ewa G. Pikus, and Jaroslav Doležel. 2021. "Kinetics of DNA Repair in Vicia faba Meristem Regeneration Following Replication Stress" Cells 10, no. 1: 88. https://doi.org/10.3390/cells10010088
APA StyleRybaczek, D., Musiałek, M. W., Vrána, J., Petrovská, B., Pikus, E. G., & Doležel, J. (2021). Kinetics of DNA Repair in Vicia faba Meristem Regeneration Following Replication Stress. Cells, 10(1), 88. https://doi.org/10.3390/cells10010088