Pathologically Decreased CSF Levels of Synaptic Marker NPTX2 in DLB Are Correlated with Levels of Alpha-Synuclein and VGF

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics, Subject Selection, and Description
2.2. CSF Biomarkers
2.3. Neuropsychological Examination
- •
- Memory domain:
- ○
- Rays auditory verbal learning test (RAVLT) immediate recall
- ○
- RAVLT delayed recall
- ○
- Visual association test A was used
- •
- Attention
- ○
- Digit span forward
- ○
- Trail making test A
- ○
- Stroop 1
- ○
- Stroop 2
- •
- Executive function
- ○
- Digit span backward
- ○
- Stroop 3
- ○
- Trail making test B
- ○
- Letter fluency
- ○
- Frontal assessment battery
- •
- Language
- ○
- Dutch version of the Boston naming test
- ○
- Category fluency test
- •
- Visual spatial
- ○
- Visual object and space perception (VOSP) number location
- ○
- VOSP dot counting
- ○
- VOSP frat let
2.4. Statistics
3. Results
3.1. Study Population
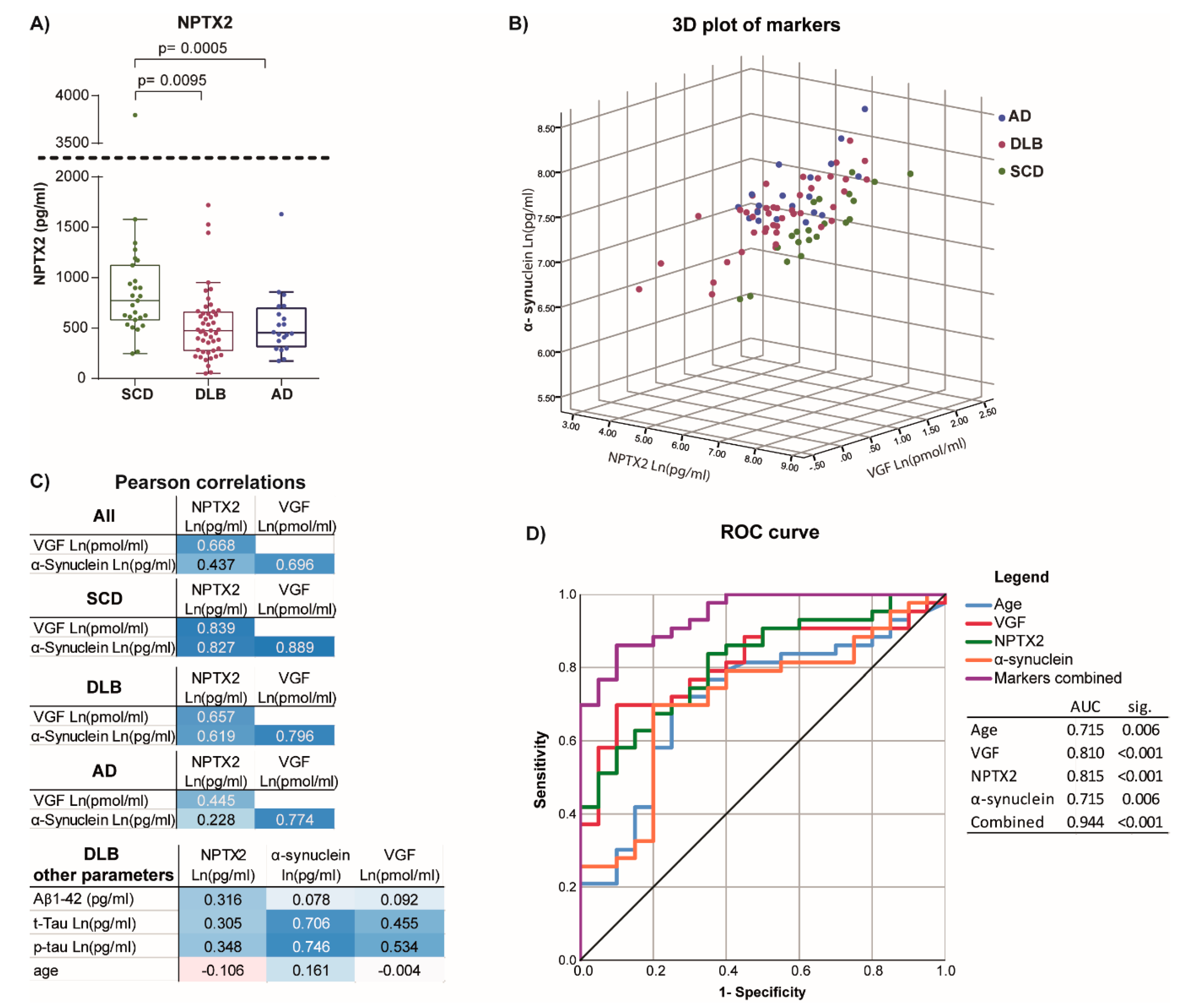
3.2. Correlation to Other Synaptic Markers
3.3. Differentiating Potential of Combined Markers
3.4. NPTX2 Levels Relate to Cognitive Function and Decline
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Toledo, J.B.; Cairns, N.J.; Da, X.; Chen, K.; Carter, D.; Fleisher, A.; Householder, E.; Ayutyanont, N.; Roontiva, A.; Bauer, R.J.; et al. Clinical and multimodal biomarker correlates of ADNI neuropathological findings. Acta Neuropathol. Commun. 2013, 1, 65. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.; McAleese, K.E.; Thomas, A.J.; Johnson, M.; Martin-Ruiz, C.; Parker, C.; Colloby, S.J.; Jellinger, K.; Attems, J. Neuropathologically mixed Alzheimer’s and Lewy body disease: Burden of pathological protein aggregates differs between clinical phenotypes. Acta Neuropathol. 2015, 129, 729–748. [Google Scholar] [CrossRef] [PubMed]
- van Steenoven, I.; Aarsland, D.; Weintraub, D.; Londos, E.; Blanc, F.; van der Flier, W.M.; Teunissen, C.E.; Mollenhauer, B.; Fladby, T.; Kramberger, M.G.; et al. Cerebrospinal Fluid Alzheimer’s Disease Biomarkers Across the Spectrum of Lewy Body Diseases: Results from a Large Multicenter Cohort. J. Alzheimers Dis. 2016, 54, 287–295. [Google Scholar] [CrossRef]
- Mukaetova-Ladinska, E.B.; Andras, A.; Milne, J.; Abdel-All, Z.; Borr, I.; Jaros, E.; Perry, R.H.; Honer, W.G.; Cleghorn, A.; Doherty, J.; et al. Synaptic proteins and choline acetyltransferase loss in visual cortex in dementia with Lewy bodies. J. Neuropathol. Exp. Neurol. 2013, 72, 53–60. [Google Scholar] [CrossRef]
- Schulz-Schaeffer, W.J. The synaptic pathology of alpha-synuclein aggregation in dementia with Lewy bodies, Parkinson’s disease and Parkinson’s disease dementia. Acta Neuropathol. 2010, 120, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Khundakar, A.A.; Hanson, P.S.; Erskine, D.; Lax, N.Z.; Roscamp, J.; Karyka, E.; Tsefou, E.; Singh, P.; Cockell, S.J.; Gribben, A.; et al. Analysis of primary visual cortex in dementia with Lewy bodies indicates GABAergic involvement associated with recurrent complex visual hallucinations. Acta Neuropathol. Commun. 2016, 4, 66. [Google Scholar] [CrossRef] [PubMed]
- Azar, M.; Chapman, S.; Gu, Y.; Leverenz, J.B.; Stern, Y.; Cosentino, S. Cognitive tests aid in clinical differentiation of Alzheimer’s disease versus Alzheimer’s disease with Lewy body disease: Evidence from a pathological study. Alzheimers Dement. 2020, 16, 1173–1181. [Google Scholar] [CrossRef]
- Hamilton, J.M.; Landy, K.M.; Salmon, D.P.; Hansen, L.A.; Masliah, E.; Galasko, D. Early visuospatial deficits predict the occurrence of visual hallucinations in autopsy-confirmed dementia with Lewy bodies. Am. J. Geriatr. Psychiatr. 2012, 20, 773–781. [Google Scholar] [CrossRef]
- McKeith, I.G.; Boeve, B.F.; Dickson, D.W.; Halliday, G.; Taylor, J.P.; Weintraub, D.; Aarsland, D.; Galvin, J.; Attems, J.; Ballard, C.G.; et al. Diagnosis and management of dementia with Lewy bodies: Fourth consensus report of the DLB Consortium. Neurology 2017, 89, 88–100. [Google Scholar] [CrossRef]
- Sanford, A.M. Lewy Body Dementia. Clin. Geriatr. Med. 2018, 34, 603–615. [Google Scholar] [CrossRef]
- Bate, C.; Gentleman, S.; Williams, A. alpha-synuclein induced synapse damage is enhanced by amyloid-beta1-42. Mol. Neurodegener. 2010, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.L.; Schulz-Schaeffer, W.J. Presynaptic alpha-synuclein aggregates, not Lewy bodies, cause neurodegeneration in dementia with Lewy bodies. J. Neurosci. 2007, 27, 1405–1410. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Mila-Aloma, M.; Suarez-Calvet, M.; Molinuevo, J.L. Latest advances in cerebrospinal fluid and blood biomarkers of Alzheimer’s disease. Adv. Neurol. Disord. 2019, 12, 1756286419888819. [Google Scholar] [CrossRef] [PubMed]
- van Steenoven, I.; Koel-Simmelink, M.J.A.; Vergouw, L.J.M.; Tijms, B.M.; Piersma, S.R.; Pham, T.V.; Bridel, C.; Ferri, G.L.; Cocco, C.; Noli, B.; et al. Identification of novel cerebrospinal fluid biomarker candidates for dementia with Lewy bodies: A proteomic approach. Mol. Neurodegener. 2020, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Sathe, G.; Na, C.H.; Renuse, S.; Madugundu, A.K.; Albert, M.; Moghekar, A.; Pandey, A. Quantitative Proteomic Profiling of Cerebrospinal Fluid to Identify Candidate Biomarkers for Alzheimer’s Disease. Proteom. Clin. Appl. 2019, 13, e1800105. [Google Scholar] [CrossRef]
- Chang, M.C.; Park, J.M.; Pelkey, K.A.; Grabenstatter, H.L.; Xu, D.; Linden, D.J.; Sutula, T.P.; McBain, C.J.; Worley, P.F. Narp regulates homeostatic scaling of excitatory synapses on parvalbumin-expressing interneurons. Nat. Neurosci. 2010, 13, 1090–1097. [Google Scholar] [CrossRef]
- Mariga, A.; Glaser, J.; Mathias, L.; Xu, D.; Xiao, M.; Worley, P.; Ninan, I.; Chao, M.V. Definition of a Bidirectional Activity-Dependent Pathway Involving BDNF and Narp. Cell Rep. 2015, 13, 1747–1756. [Google Scholar] [CrossRef]
- Xu, D.; Hopf, C.; Reddy, R.; Cho, R.W.; Guo, L.; Lanahan, A.; Petralia, R.S.; Wenthold, R.J.; O’Brien, R.J.; Worley, P. Narp and NP1 form heterocomplexes that function in developmental and activity-dependent synaptic plasticity. Neuron 2003, 39, 513–528. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Xu, D.; Petralia, R.S.; Steward, O.; Huganir, R.L.; Worley, P. Synaptic clustering of AMPA receptors by the extracellular immediate-early gene product Narp. Neuron 1999, 23, 309–323. [Google Scholar] [CrossRef]
- Xiao, M.F.; Xu, D.; Craig, M.T.; Pelkey, K.A.; Chien, C.C.; Shi, Y.; Zhang, J.; Resnick, S.; Pletnikova, O.; Salmon, D.; et al. NPTX2 and cognitive dysfunction in Alzheimer’s Disease. Elife 2017, 6, e23798. [Google Scholar] [CrossRef] [PubMed]
- Galasko, D.; Xiao, M.; Xu, D.; Smirnov, D.; Salmon, D.P.; Dewit, N.; Vanbrabant, J.; Jacobs, D.; Vanderstichele, H.; Vanmechelen, E.; et al. Synaptic biomarkers in CSF aid in diagnosis, correlate with cognition and predict progression in MCI and Alzheimer’s disease. Alzheimers Dement. (N. Y.) 2019, 5, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Soldan, A.; Moghekar, A.; Walker, K.A.; Pettigrew, C.; Hou, X.; Lu, H.; Miller, M.I.; Alfini, A.; Albert, M.; Xu, D.; et al. Resting-State Functional Connectivity Is Associated With Cerebrospinal Fluid Levels of the Synaptic Protein NPTX2 in Non-demented Older Adults. Front. Aging Neurosci. 2019, 11, 132. [Google Scholar] [CrossRef]
- Belbin, O.; Xiao, M.F.; Xu, D.; Carmona-Iragui, M.; Pegueroles, J.; Benejam, B.; Videla, L.; Fernandez, S.; Barroeta, I.; Nunez-Llaves, R.; et al. Cerebrospinal fluid profile of NPTX2 supports role of Alzheimer’s disease-related inhibitory circuit dysfunction in adults with Down syndrome. Mol. Neurodegener. 2020, 15, 46. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, E.J.; Abner, E.L.; Jicha, G.A.; Kapogiannis, D.; Schwartz, J.B. Declining levels of functionally specialized synaptic proteins in plasma neuronal exosomes with progression of Alzheimer’s disease. FASEB J. 2018, 32, 888–893. [Google Scholar] [CrossRef]
- Alder, J.; Thakker-Varia, S.; Bangasser, D.A.; Kuroiwa, M.; Plummer, M.R.; Shors, T.J.; Black, I.B. Brain-derived neurotrophic factor-induced gene expression reveals novel actions of VGF in hippocampal synaptic plasticity. J. Neurosci. 2003, 23, 10800–10808. [Google Scholar] [CrossRef]
- Lewis, J.E.; Brameld, J.M.; Jethwa, P.H. Neuroendocrine Role for VGF. Front. Endocrinol. (Lausanne) 2015, 6, 3. [Google Scholar] [CrossRef]
- Beckmann, N.D.; Lin, W.J.; Wang, M.; Cohain, A.T.; Charney, A.W.; Wang, P.; Ma, W.; Wang, Y.C.; Jiang, C.; Audrain, M.; et al. Multiscale causal networks identify VGF as a key regulator of Alzheimer’s disease. Nat. Commun. 2020, 11, 3942. [Google Scholar] [CrossRef] [PubMed]
- Possenti, R.; Eldridge, J.D.; Paterson, B.M.; Grasso, A.; Levi, A. A protein induced by NGF in PC12 cells is stored in secretory vesicles and released through the regulated pathway. EMBO J. 1989, 8, 2217–2223. [Google Scholar] [CrossRef]
- van Steenoven, I.; Noli, B.; Cocco, C.; Ferri, G.L.; Oeckl, P.; Otto, M.; Koel-Simmelink, M.J.A.; Bridel, C.; van der Flier, W.M.; Lemstra, A.W.; et al. VGF Peptides in Cerebrospinal Fluid of Patients with Dementia with Lewy Bodies. Int. J. Mol. Sci. 2019, 20, 4674. [Google Scholar] [CrossRef]
- Duits, F.H.; Brinkmalm, G.; Teunissen, C.E.; Brinkmalm, A.; Scheltens, P.; Van der Flier, W.M.; Zetterberg, H.; Blennow, K. Synaptic proteins in CSF as potential novel biomarkers for prognosis in prodromal Alzheimer’s disease. Alzheimers Res. 2018, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Burre, J.; Sharma, M.; Sudhof, T.C. Cell Biology and Pathophysiology of alpha-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8, a024091. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, B.; Li, X.; Fu, C.; Wang, C.; Kang, X. alpha-Synuclein: A Multifunctional Player in Exocytosis, Endocytosis, and Vesicle Recycling. Front. Neurosci. 2019, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Edwards, R.H. The physiological role of alpha-synuclein and its relationship to Parkinson’s Disease. J. Neurochem. 2019, 150, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Chiasserini, D.; Biscetti, L.; Eusebi, P.; Salvadori, N.; Frattini, G.; Simoni, S.; De Roeck, N.; Tambasco, N.; Stoops, E.; Vanderstichele, H.; et al. Differential role of CSF fatty acid binding protein 3, alpha-synuclein, and Alzheimer’s disease core biomarkers in Lewy body disorders and Alzheimer’s dementia. Alzheimers Res. 2017, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Mavroudis, I.; Petridis, F.; Chatzikonstantinou, S.; Kazis, D. Alpha-synuclein Levels in the Differential Diagnosis of Lewy Bodies Dementia and Other Neurodegenerative Disorders: A Meta-analysis. Alzheimer Dis. Assoc. Disord. 2020, 34, 220–224. [Google Scholar] [CrossRef]
- van Steenoven, I.; Majbour, N.K.; Vaikath, N.N.; Berendse, H.W.; van der Flier, W.M.; van de Berg, W.D.J.; Teunissen, C.E.; Lemstra, A.W.; El-Agnaf, O.M.A. alpha-Synuclein species as potential cerebrospinal fluid biomarkers for dementia with lewy bodies. Mov. Disord. 2018, 33, 1724–1733. [Google Scholar] [CrossRef]
- Viode, A.; Epelbaum, S.; Benyounes, I.; Verny, M.; Dubois, B.; Junot, C.; Fenaille, F.; Lamari, F.; Becher, F. Simultaneous quantification of tau and alpha-synuclein in cerebrospinal fluid by high-resolution mass spectrometry for differentiation of Lewy Body Dementia from Alzheimer’s Disease and controls. Analyst 2019, 144, 6342–6351. [Google Scholar] [CrossRef]
- van der Flier, W.M.; Pijnenburg, Y.A.; Prins, N.; Lemstra, A.W.; Bouwman, F.H.; Teunissen, C.E.; van Berckel, B.N.; Stam, C.J.; Barkhof, F.; Visser, P.J.; et al. Optimizing patient care and research: The Amsterdam Dementia Cohort. J. Alzheimers Dis. 2014, 41, 313–327. [Google Scholar] [CrossRef]
- Jessen, F.; Amariglio, R.E.; van Boxtel, M.; Breteler, M.; Ceccaldi, M.; Chetelat, G.; Dubois, B.; Dufouil, C.; Ellis, K.A.; van der Flier, W.M.; et al. A conceptual framework for research on subjective cognitive decline in preclinical Alzheimer’s disease. Alzheimers Dement. 2014, 10, 844–852. [Google Scholar] [CrossRef]
- Tijms, B.M.; Willemse, E.A.J.; Zwan, M.D.; Mulder, S.D.; Visser, P.J.; van Berckel, B.N.M.; van der Flier, W.M.; Scheltens, P.; Teunissen, C.E. Unbiased Approach to Counteract Upward Drift in Cerebrospinal Fluid Amyloid-beta 1-42 Analysis Results. Clin. Chem. 2018, 64, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Mollenhauer, B.; Bowman, F.D.; Drake, D.; Duong, J.; Blennow, K.; El-Agnaf, O.; Shaw, L.M.; Masucci, J.; Taylor, P.; Umek, R.M.; et al. Antibody-based methods for the measurement of alpha-synuclein concentration in human cerebrospinal fluid-method comparison and round robin study. J. Neurochem. 2019, 149, 126–138. [Google Scholar] [CrossRef] [PubMed]
- van de Beek, M.; Babapour Mofrad, R.; van Steenoven, I.; Vanderstichele, H.; Scheltens, P.; Teunissen, C.E.; Lemstra, A.W.; van der Flier, W.M. Sex-specific associations with cerebrospinal fluid biomarkers in dementia with Lewy bodies. Alzheimers Res. 2020, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Reichert Plaska, C.; Clark, D.P.A.; Zetterberg, H.; Blennow, K.; Verbeek, M.M.; Pomara, N. CSF alpha-synuclein correlates with CSF neurogranin in late-life depression. Int. J. Neurosci. 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; van Steenoven, I.; Huisman, E.; Oosterveld, L.; Berendse, H.; van der Flier, W.M.; Del Campo, M.; Lemstra, A.W.; van de Berg, W.D.J.; Teunissen, C.E. Contactin-1 Is Reduced in Cerebrospinal Fluid of Parkinson’s Disease Patients and Is Present within Lewy Bodies. Biomolecules 2020, 10, 177. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| SCD n = 27 | DLB n = 48 | AD n = 20 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean (SD) | n(%) | Mean (SD) | n(%) | Mean (SD) | n(%) | F-Test | SCD-DLB | SCD-AD | DLB-AD | |
| Female % | 4(15) | 6 (13) | 2 (10) | |||||||
| Age | 63.7 (5.9) | 67.7 (6.4) | 65.3 (6.0) | 0.018 | 0.015 | 0.827 | 0.560 | |||
| Education | 5.7 (1.0) | 5.1 (1.3) | 5.3 (1.0) | 0.138 | N.R. | N.R. | N.R. | |||
| Global | −0.21 (0.53) | 21 | −2.48 (1.16) | 40 | −4.00 (2.03) | 17 | <0.001 | <0.001 | <0.001 | 0.139 |
| Memory | −0.15 (0.8) | 21 | −2.76 (1.7) | 40 | −2.99 (1.95) | 13 | <0.001 | <0.001 | <0.001 | 1.000 |
| Attention | −0.28 (0.71) | 21 | −2.86 (2.15) | 40 | −3.63 (2.48) | 13 | <0.001 | <0.001 | <0.001 | 1.000 |
| Executive function | −0.23 (0.73) | 21 | −2.49 (1.29) | 40 | −3.08 (1.35) | 13 | <0.001 | <0.001 | <0.001 | 0.792 |
| Language | −0.13 (0.61) | 21 | −1.14 (0.68) | 40 | −1.99 (1.74) | 12 | <0.001 | <0.001 | <0.001 | 0.780 |
| Visual spatial | −0.21 (0.89) | 14 | −2.22 (1.85) | 30 | −4.79 (3.27) | 10 | <0.001 | 0.003 | <0.001 | 0.166 |
| MMSE | 28.12 (2.02) | 18 | 23.07 (4.40) | 42 | 18.67 (542.) | 18 | <0.001 | <0.001 | <0.001 | 0.048 |
| Median [95%interval], n | Median [95%interval], n | Median [95%interval], n | F-test | SCD-DLB | SCD-AD | DLB-AD | ||||
| Aβ1–42 (pg/mL) | 1032 [982–1112], 26 | 788 [760–905], 47 | 586 [531–614], 19 | <0.001 | 0.002 | <0.001 | <0.001 | |||
| T-Tau (pg/mL) | 199 [182–237], 26 | 299 [285–393],48 | 596 [522–949], 19 | <0.001 | 0.003 | <0.001 | <0.001 | |||
| p-Tau (pg/mL) | 41.0 [34.7–42.8], 26 | 47.0 [43.4–58.6], 47 | 87.5 [77.6–120.7], 20 | <0.001 | 0.101 | <0.001 | <0.001 | |||
| NPTX2 (pg/mL) | 773 [581–1123], 27 | 474 [279–659], 48 | 453 [317–696], 20 | <0.001 | <0.001 | 0.009 | 1.000 | |||
| α-synuclein (pg/mL) | 1508 [1281–1971], 22 | 1811 [1537–2299], 44 | 2009 [1764–2658], 19 | 0.004 | 0.156 | 0.003 | 0.166 | |||
| VGF (pmol/mL) | 3.66 [2.97–5.15], 22 | 2.58 [2.04–3.42] 44 | 3.33 [2.40–4.41] 20 | 0.007 | 0.009 | 1.000 | 0.136 | |||
| 95% Confidence | ||||
|---|---|---|---|---|
| Cognitive function | β | Sig. | Lower | Upper |
| Global median n = 39 | 0.2642 | 0.045 | 0.006 | 0.522 |
| Memory n = 39 | 0.1931 | 0.595 | −0.538 | 0.925 |
| Attention n = 39 | 1.0938 | 0.019 | 0.194 | 1.994 |
| Executive =39 | −0.0957 | 0.755 | −0.714 | 0.523 |
| Language n = 39 | 0.2681 | 0.063 | −0.015 | 0.552 |
| Visual n = 30 | −0.7258 | 0.197 | −1.853 | 0.401 |
| MMSE n = 40 | 0.7791 | 0.406 | −1.102 | 2.660 |
| Cognitive decline | ||||
| Global median n = 32 | 0.0006 | 0.837 | −0.005 | 0.007 |
| Memory n = 32 | 0.0019 | 0.831 | −0.016 | 0.020 |
| Attention n = 32 | 0.0056 | 0.753 | −0.030 | 0.041 |
| Executive n = 32 | 0.0001 | 0.985 | −0.014 | 0.014 |
| Language n = 32 | 0.0031 | 0.393 | −0.004 | 0.010 |
| Visual n = 28 | 0.0436 | 0.002 | 0.016 | 0.071 |
| MMSE n = 33 | 0.0014 | 0.950 | −0.041 | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boiten, W.A.; van Steenoven, I.; Xiao, M.-F.; Worley, P.F.; Noli, B.; Cocco, C.; Ferri, G.-L.; Lemstra, A.W.; Teunissen, C.E. Pathologically Decreased CSF Levels of Synaptic Marker NPTX2 in DLB Are Correlated with Levels of Alpha-Synuclein and VGF. Cells 2021, 10, 38. https://doi.org/10.3390/cells10010038
Boiten WA, van Steenoven I, Xiao M-F, Worley PF, Noli B, Cocco C, Ferri G-L, Lemstra AW, Teunissen CE. Pathologically Decreased CSF Levels of Synaptic Marker NPTX2 in DLB Are Correlated with Levels of Alpha-Synuclein and VGF. Cells. 2021; 10(1):38. https://doi.org/10.3390/cells10010038
Chicago/Turabian StyleBoiten, Walter A., Inger van Steenoven, Mei-Fang Xiao, Paul F. Worley, Barbara Noli, Cristina Cocco, Gian-Luca Ferri, Afina W. Lemstra, and Charlotte E. Teunissen. 2021. "Pathologically Decreased CSF Levels of Synaptic Marker NPTX2 in DLB Are Correlated with Levels of Alpha-Synuclein and VGF" Cells 10, no. 1: 38. https://doi.org/10.3390/cells10010038
APA StyleBoiten, W. A., van Steenoven, I., Xiao, M.-F., Worley, P. F., Noli, B., Cocco, C., Ferri, G.-L., Lemstra, A. W., & Teunissen, C. E. (2021). Pathologically Decreased CSF Levels of Synaptic Marker NPTX2 in DLB Are Correlated with Levels of Alpha-Synuclein and VGF. Cells, 10(1), 38. https://doi.org/10.3390/cells10010038