IL-33 and Superantigenic Activation of Human Lung Mast Cells Induce the Release of Angiogenic and Lymphangiogenic Factors

 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Human Monoclonal IgM and IgE and Human Polyclonal IgG
2.3. Isolation of HLMCs
2.4. Histamine Release
2.5. VEGF-A and VEGF-C Release
2.6. Statistical Analysis
3. Results
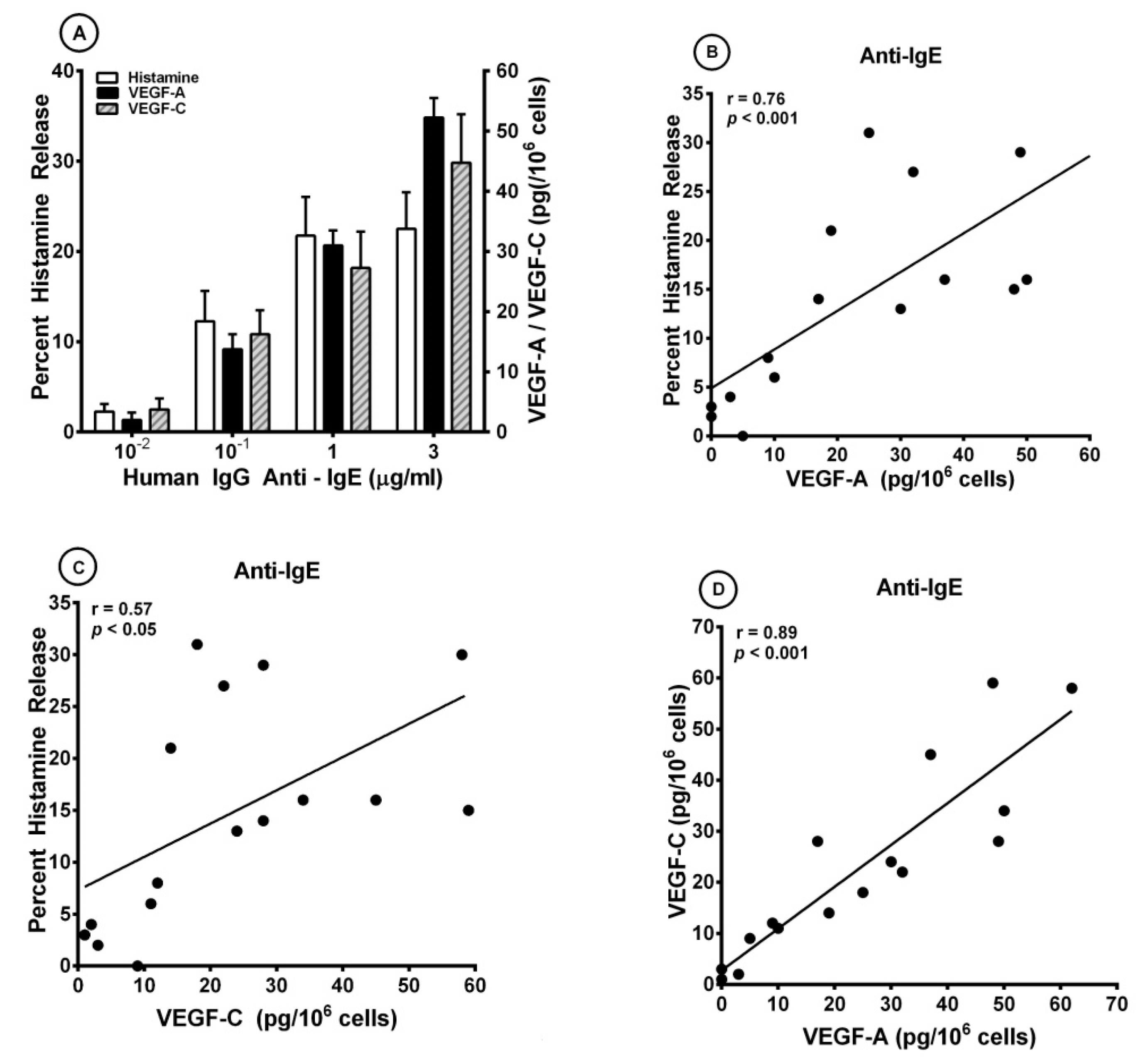
3.1. Effects of Human IgG Anti-IgE on the Release of Angiogenic and Lymphangiogenic Factors from HLMCs
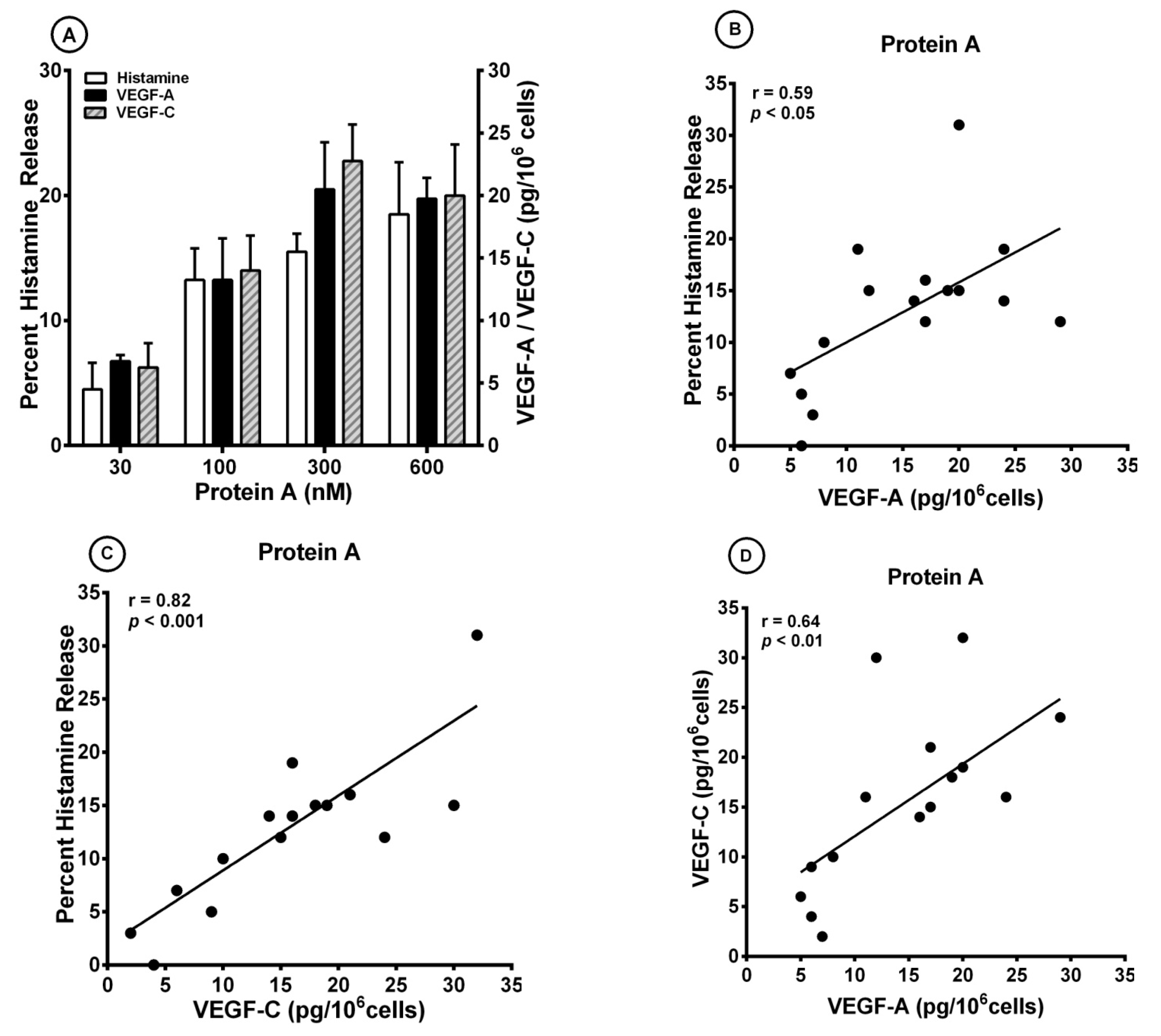
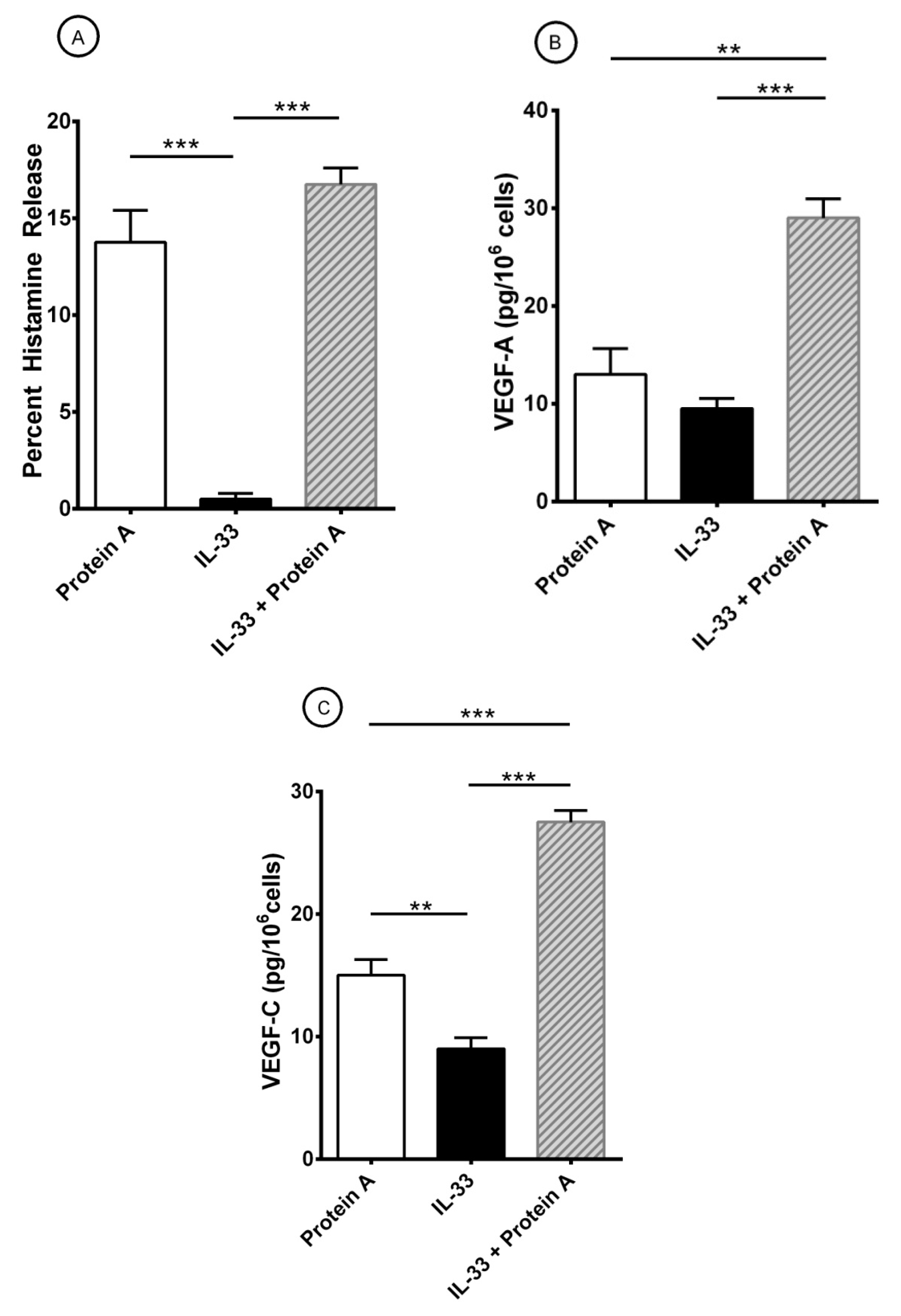
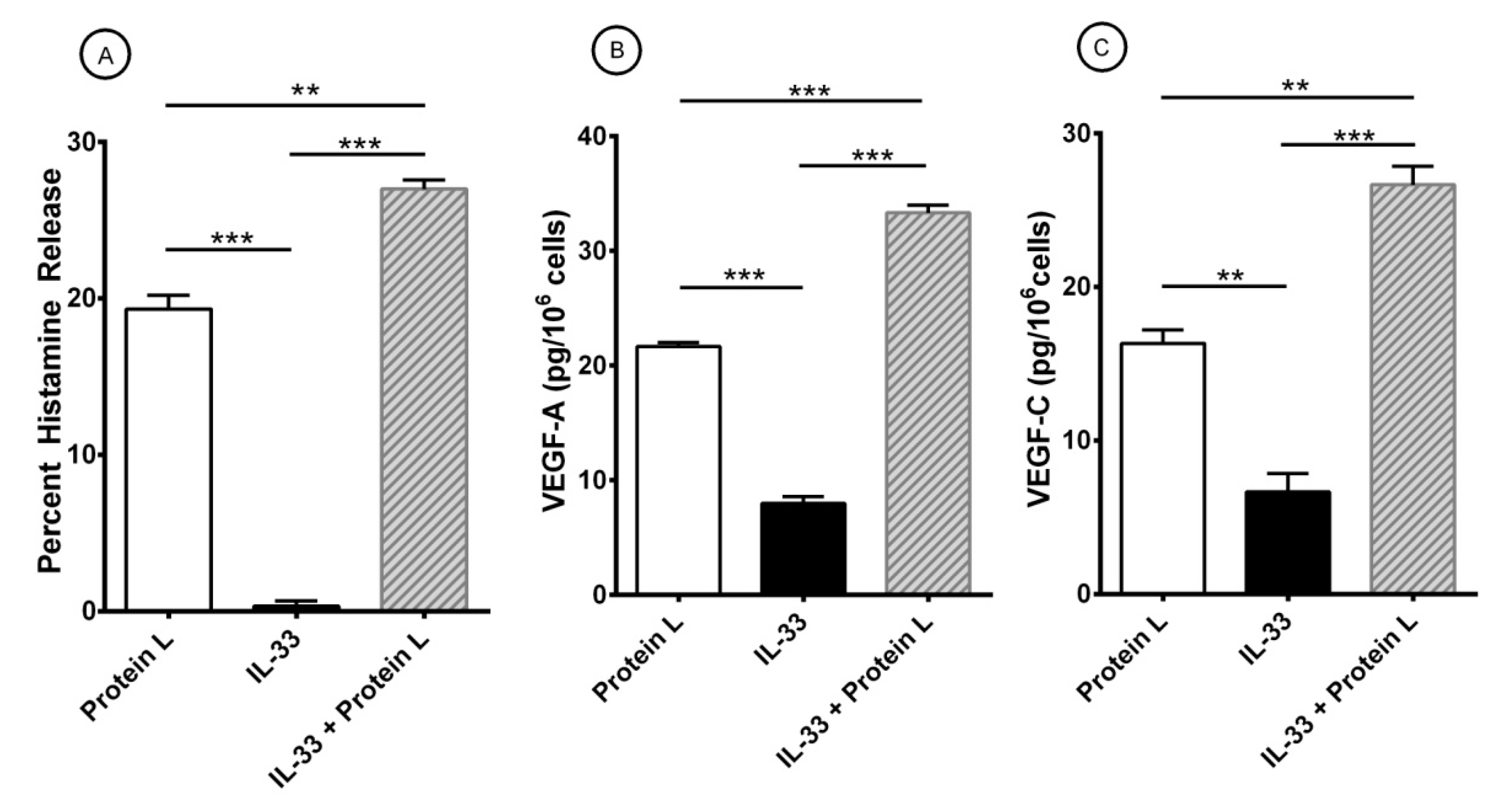
3.2. Effect of Superantigenic Protein A on the Release of Angiogenic and Lymphangiogenic Factors from HLMCs
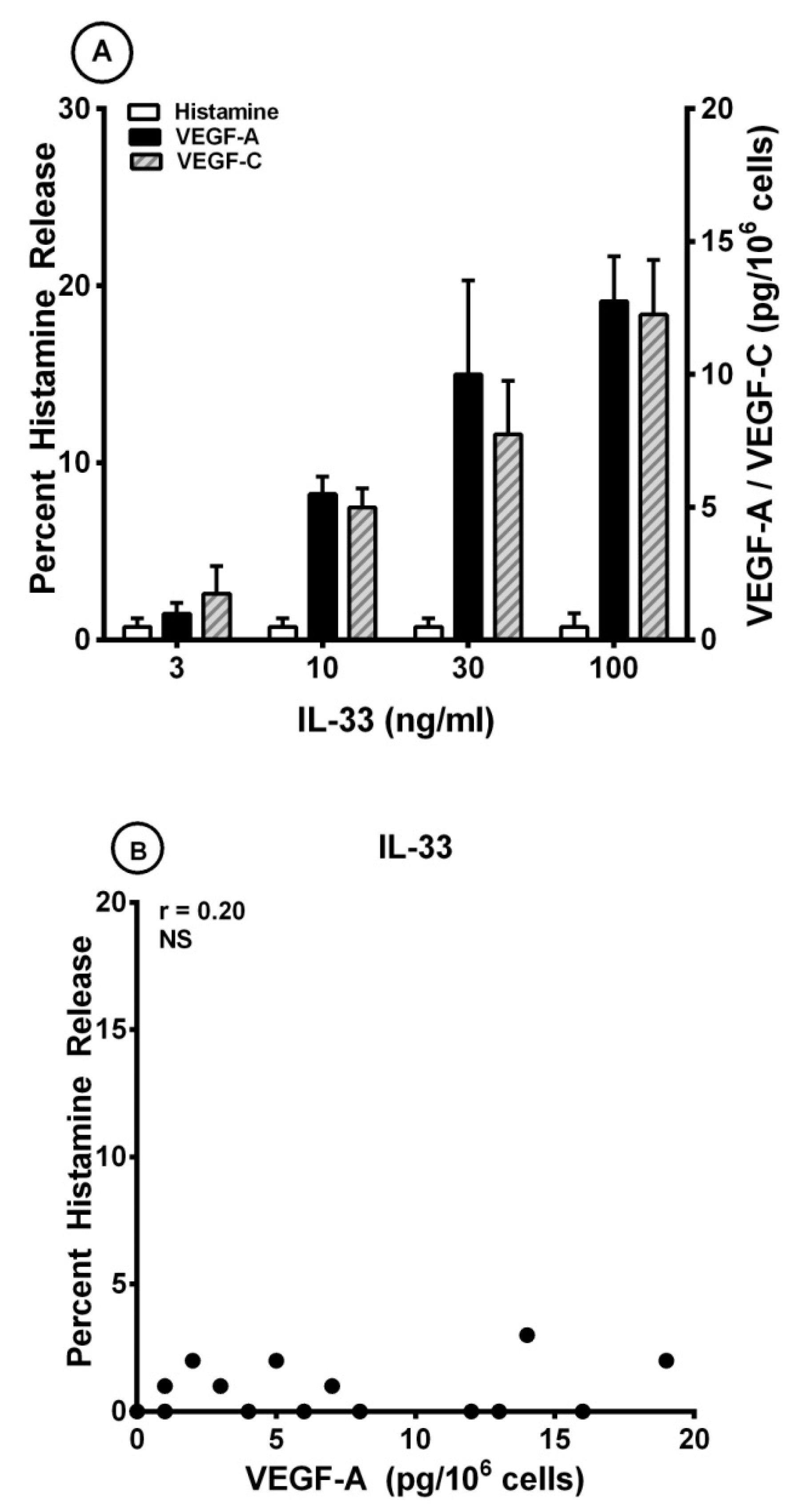
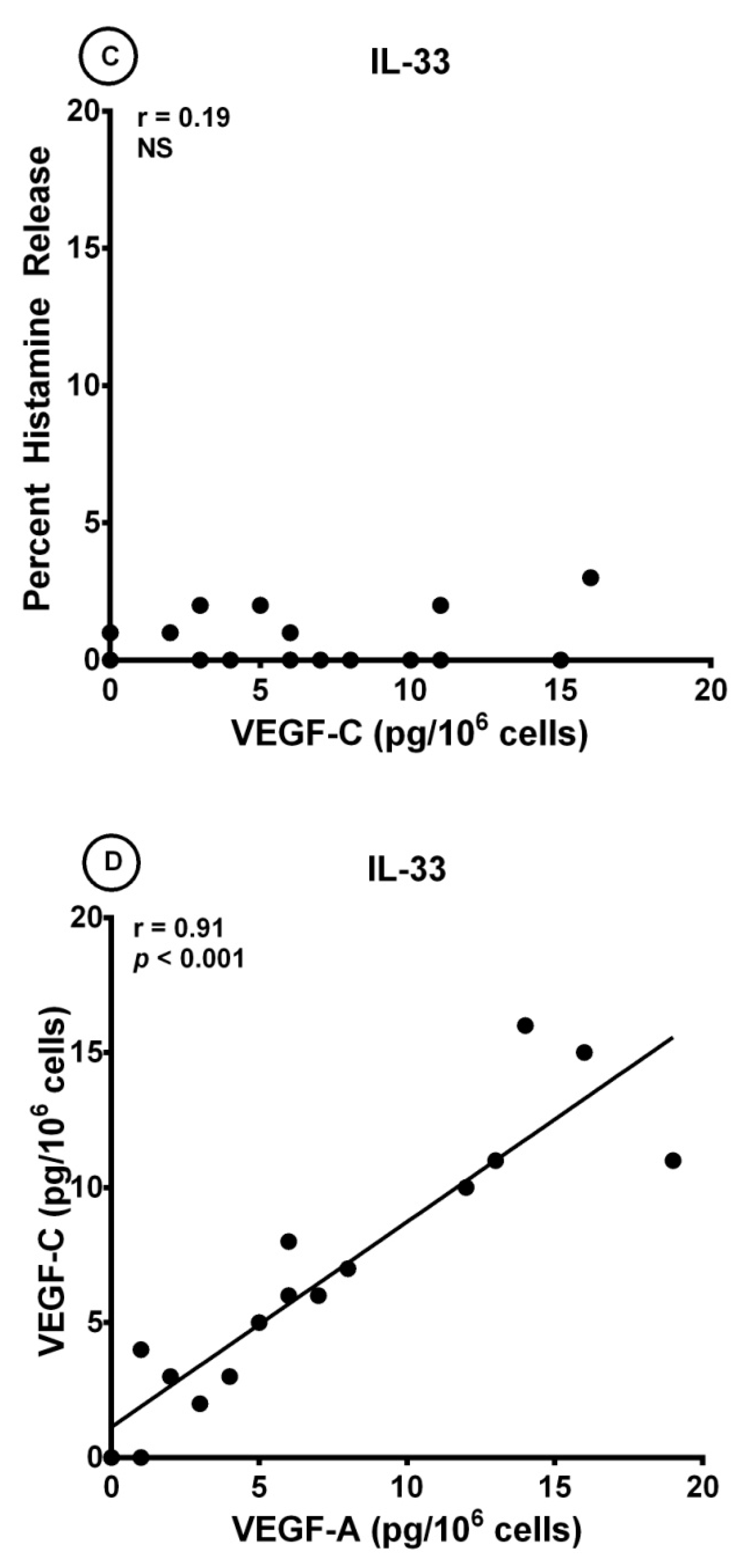
3.3. Effects of IL-33 on the Release of Angiogenic and Lymphangiogenic Factors from HLMCs
3.4. Effect of Short-Term Priming by IL-33 on Superantigenic Release of Mediators from HLMCs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AHR | Airway hyperresponsiveness |
| ASM | Airway smooth muscle |
| BSA | Bovine serum albumin |
| COPD | Chronic obstructive pulmonary disease |
| CBMC | Cord blood-derived mast cell |
| FcεRI | High-affinity receptor for IgE |
| FCS | Fetal calf serum |
| H | Heavy |
| H-aIgE | Human IgG anti-IgE |
| HLMC | Human lung mast cell |
| Ig | Immunoglobulin |
| IL-33 | Interleukin-33 |
| IL | Interleukin |
| L | Light |
| LTC4 | Cysteinyl leukotriene C4 |
| mAb | Monoclonal antibody |
| P. magnus | Peptostreptococcus magnus |
| PBMC | Peripheral blood-derived mast cell |
| PGD2 | Prostaglandin D2 |
| S. aureus | Staphylococcus aureus |
| SAg | Superantigen |
| SE | Staphylococcus aureus enterotoxins |
| SP | Substance P |
| TCR | T cell receptor |
| Treg | regulatory T cell |
| TSLP | Thymic stromal lymphopoietin |
| V | Variable |
| VEGF | Vascular endothelial growth factor |
References
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; de Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 179, 247–261. [Google Scholar] [CrossRef]
- Borriello, F.; Granata, F.; Varricchi, G.; Genovese, A.; Triggiani, M.; Marone, G. Immunopharmacological Modulation of Mast Cells. Curr. Opin. Pharmacol. 2014, 17, 45–57. [Google Scholar] [CrossRef]
- Casolaro, V.; Galeone, D.; Giacummo, A.; Sanduzzi, A.; Melillo, G.; Marone, G. Human Basophil/Mast Cell Releasability. V. Functional Comparisons of Cells Obtained from Peripheral Blood, Lung Parenchyma, and Bronchoalveolar Lavage in Asthmatics. Am. Rev. Respir. Dis. 1989, 139, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Zilionis, R.; Engblom, C.; Pfirschke, C.; Savova, V.; Zemmour, D.; Saatcioglu, H.D.; Krishnan, I.; Maroni, G.; Meyerovitz, C.V.; Kerwin, C.M.; et al. Single-Cell Transcriptomics of Human and Mouse Lung Cancers Reveals Conserved Myeloid Populations across Individuals and Species. Immunity 2019, 50, 1317–1334.e10. [Google Scholar] [CrossRef] [PubMed]
- Olivera, A.; Beaven, M.A.; Metcalfe, D.D. Mast Cells Signal Their Importance in Health and Disease. J. Allergy Clin. Immunol. 2018, 142, 381–393. [Google Scholar] [PubMed]
- Piliponsky, A.M.; Romani, L. The Contribution of Mast Cells to Bacterial and Fungal Infection Immunity. Immunol. Rev. 2018, 282, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Raap, U.; Rivellese, F.; Marone, G.; Gibbs, B.F. Human Mast Cells and Basophils-How Are They Similar How Are They Different? Immunol. Rev. 2018, 282, 8–34. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast Cells as Sources of Cytokines, Chemokines, and Growth Factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Bradding, P.; Arthur, G. Mast Cells in Asthma--State of the Art. Clin. Exp. Allergy 2016, 46, 194–263. [Google Scholar]
- Marone, G.; Varricchi, G.; Loffredo, S.; Galdiero, M.R.; Rivellese, F.; de Paulis, A. Are Basophils and Mast Cells Masters in Hiv Infection? Int. Arch. Allergy Immunol. 2016, 171, 158–165. [Google Scholar]
- Suurmond, J.; Rivellese, F.; Dorjee, A.L.; Bakker, A.M.; Rombouts, Y.J.; Rispens, T.; Wolbink, G.; Zaldumbide, A.; Hoeben, R.C.; Huizinga, T.W.; et al. Toll-Like Receptor Triggering Augments Activation of Human Mast Cells by Anti-Citrullinated Protein Antibodies. Ann. Rheum. Dis. 2015, 74, 1915–1923. [Google Scholar] [CrossRef]
- Piliponsky, A.M.; Acharya, M.; Shubin, N.J. Mast Cells in Viral, Bacterial, and Fungal Infection Immunity. Int. J. Mol. Sci. 2019, 20, 2851. [Google Scholar] [CrossRef]
- Voehringer, D. Protective and Pathological Roles of Mast Cells and Basophils. Nat. Rev. Immunol. 2013, 13, 362–375. [Google Scholar] [CrossRef]
- Detoraki, A.; Staiano, R.I.; Granata, F.; Giannattasio, G.; Prevete, N.; de Paulis, A.; Ribatti, D.; Genovese, A.; Triggiani, M.; Marone, G. Vascular Endothelial Growth Factors Synthesized by Human Lung Mast Cells Exert Angiogenic Effects. J. Allergy Clin. Immunol. 2009, 123, 1142–1149. [Google Scholar] [CrossRef]
- Varricchi, G.; Loffredo, S.; Galdiero, M.R.; Marone, G.; Cristinziano, L.; Granata, F. Innate Effector Cells in Angiogenesis and Lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 152–160. [Google Scholar] [CrossRef]
- Marone, G.; Varricchi, G.; Loffredo, S.; Granata, F. Mast Cells and Basophils in Inflammatory and Tumor Angiogenesis and Lymphangiogenesis. Eur. J. Pharmacol. 2016, 778, 146–151. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Zhang, B.; Kempuraj, D.; Tagen, M.; Vasiadi, M.; Angelidou, A.; Alysandratos, K.D.; Kalogeromitros, D.; Asadi, S.; Stavrianeas, N.; et al. Il-33 Augments Substance P-Induced Vegf Secretion from Human Mast Cells and Is Increased in Psoriatic Skin. Proc. Natl. Acad. Sci. USA 2010, 107, 4448–4453. [Google Scholar] [CrossRef]
- Detoraki, A.; Granata, F.; Staibano, S.; Rossi, F.W.; Marone, G.; Genovese, A. Angiogenesis and Lymphangiogenesis in Bronchial Asthma. Allergy 2010, 65, 946–958. [Google Scholar]
- Varricchi, G.; Granata, F.; Loffredo, S.; Genovese, A.; Marone, G. Angiogenesis and Lymphangiogenesis in Inflammatory Skin Disorders. J. Am. Acad. Dermatol. 2015, 73, 144–153. [Google Scholar] [CrossRef]
- Rivellese, F.; Suurmond, J.; Habets, K.; Dorjee, A.L.; Ramamoorthi, N.; Townsend, M.J.; de Paulis, A.; Marone, G.; Huizinga, T.W.; Pitzalis, C.; et al. Ability of Interleukin-33- and Immune Complex-Triggered Activation of Human Mast Cells to Down-Regulate Monocyte-Mediated Immune Responses. Arthritis Rheumatol. 2015, 67, 2343–2353. [Google Scholar] [CrossRef]
- Rivellese, F.; Nerviani, A.; Rossi, F.W.; Marone, G.; Matucci-Cerinic, M.; de Paulis, A.; Pitzalis, C. Mast Cells in Rheumatoid Arthritis: Friends or Foes? Autoimmun. Rev. 2017, 16, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Rivellese, F.; Mauro, D.; Nerviani, A.; Pagani, S.; Fossati-Jimack, L.; Messemaker, T.; Kurreeman, F.A.S.; Toes, R.E.M.; Ramming, A.; Rauber, S.; et al. Mast Cells in Early Rheumatoid Arthritis Associate with Disease Severity and Support B Cell Autoantibody Production. Ann. Rheum. Dis. 2018, 77, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Visciano, C.; Liotti, F.; Prevete, N.; Cali, G.; Franco, R.; Collina, F.; de Paulis, A.; Marone, G.; Santoro, M.; Melillo, R.M. Mast Cells Induce Epithelial-to-Mesenchymal Transition and Stem Cell Features in Human Thyroid Cancer Cells through an Il-8-Akt-Slug Pathway. Oncogene 2015, 34, 5175–5186. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Varricchi, G.; Marone, G. The Immune Network in Thyroid Cancer. Oncoimmunology 2016, 5, e1168556. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Marone, G.; Iannone, R.; Granata, F. Are Mast Cells Masters in Cancer? Front. Immunol. 2017, 8, 424. [Google Scholar] [CrossRef]
- Varricchi, G.; Galdiero, M.R.; Marone, G.; Granata, F.; Borriello, F. Controversial Role of Mast Cells in Skin Cancers. Exp. Dermatol. 2017, 26, 11–17. [Google Scholar] [CrossRef]
- Coleman, J.W.; Godfrey, R.C. The Number and Affinity of Ige Receptors on Dispersed Human Lung Mast Cells. Immunology 1981, 44, 859–863. [Google Scholar]
- Guo, C.B.; Kagey-Sobotka, A.; Lichtenstein, L.M.; Bochner, B.S. Immunophenotyping and Functional Analysis of Purified Human Uterine Mast Cells. Blood 1992, 79, 708–712. [Google Scholar] [CrossRef]
- Ravetch, J.V.; Kinet, J.P. Fc Receptors. Annu. Rev. Immunol. 1991, 9, 457–492. [Google Scholar] [CrossRef]
- Blank, U.; Ra, C.S.; Kinet, J.P. Characterization of Truncated Alpha Chain Products from Human, Rat, and Mouse High Affinity Receptor for Immunoglobulin E. J. Biol. Chem. 1991, 266, 2639–2646. [Google Scholar] [CrossRef]
- Kinet, J.P. The High-Affinity Ige Receptor (Fc Epsilon Ri): From Physiology to Pathology. Annu. Rev. Immunol. 1999, 17, 931–972. [Google Scholar] [CrossRef]
- Dvorak, A.M.; Schulman, E.S.; Peters, S.P.; MacGlashan, D.W.; Newball, H.H., Jr.; Schleimer, R.P.; Lichtenstein, L.M. Immunoglobulin E-Mediated Degranulation of Isolated Human Lung Mast Cells. Lab Investig. 1985, 53, 45–56. [Google Scholar]
- MacGlashan, D., Jr.; Lichtenstein, L.M. Basic Characteristics of Human Lung Mast Cell Desensitization. J. Immunol. 1987, 139, 501–505. [Google Scholar] [PubMed]
- Burke, S.M.; Issekutz, T.B.; Mohan, K.; Lee, P.W.; Shmulevitz, M.; Marshall, J.S. Human Mast Cell Activation with Virus-Associated Stimuli Leads to the Selective Chemotaxis of Natural Killer Cells by a Cxcl8-Dependent Mechanism. Blood 2008, 111, 5467–5476. [Google Scholar] [CrossRef]
- Allakhverdi, Z.; Smith, D.E.; Comeau, M.R.; Delespesse, G. Cutting Edge: The St2 Ligand Il-33 Potently Activates and Drives Maturation of Human Mast Cells. J. Immunol. 2007, 179, 2051–2054. [Google Scholar]
- Staiano, R.I.; Loffredo, S.; Borriello, F.; Iannotti, F.A.; Piscitelli, F.; Orlando, P.; Secondo, A.; Granata, F.; Lepore, M.T.; Fiorelli, A.; et al. Human Lung-Resident Macrophages Express Cb1 and Cb2 Receptors Whose Activation Inhibits the Release of Angiogenic and Lymphangiogenic Factors. J. Leukoc Biol. 2016, 99, 531–540. [Google Scholar] [CrossRef]
- De Paulis, A.; Prevete, N.; Fiorentino, I.; Rossi, F.W.; Staibano, S.; Montuori, N.; Ragno, P.; Longobardi, A.; Liccardo, B.; Genovese, A.; et al. Expression and Functions of the Vascular Endothelial Growth Factors and Their Receptors in Human Basophils. J. Immunol. 2006, 177, 7322–7331. [Google Scholar]
- Loffredo, S.; Borriello, F.; Iannone, R.; Ferrara, A.L.; Galdiero, M.R.; Gigantino, V.; Esposito, P.; Varricchi, G.; Lambeau, G.; Cassatella, M.A.; et al. Group V Secreted Phospholipase A2 Induces the Release of Proangiogenic and Antiangiogenic Factors by Human Neutrophils. Front. Immunol. 2017, 8, 443. [Google Scholar] [CrossRef]
- Thomas, D.; Chou, S.; Dauwalder, O.; Lina, G. Diversity in Staphylococcus Aureus Enterotoxins. Chem. Immunol. Allergy 2007, 93, 24–41. [Google Scholar]
- Abdurrahman, G.; Schmiedeke, F.; Bachert, C.; Broker, B.M.; Holtfreter, S. Allergy-a New Role for T Cell Superantigens of Staphylococcus Aureus? Toxins (Basel) 2020, 12, 176. [Google Scholar]
- Becker, S.; Frankel, M.B.; Schneewind, O.; Missiakas, D. Release of Protein a from the Cell Wall of Staphylococcus Aureus. Proc. Natl. Acad. Sci. USA 2014, 111, 1574–1579. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, A.; Sjoquist, J. Protein a from S. Aureus. I. Pseudo-Immune Reaction with Human Gamma-Globulin. J. Immunol. 1966, 97, 822–827. [Google Scholar] [PubMed]
- Inganas, M. Comparison of Mechanisms of Interaction between Protein a from Staphylococcus Aureus and Human Monoclonal Igg, Iga and Igm in Relation to the Classical Fc Gamma and the Alternative F(Ab’)2 Epsilon Protein a Interactions. Scand. J. Immunol. 1981, 13, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Goodyear, C.S.; Silverman, G.J. B Cell Superantigens: A Microbe’s Answer to Innate-Like B Cells and Natural Antibodies. Springer Semin. Immunopathol. 2005, 26, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Bjorck, L.; Protein, L. A Novel Bacterial Cell Wall Protein with Affinity for Ig L Chains. J. Immunol. 1988, 140, 1194–1197. [Google Scholar] [PubMed]
- Patella, V.; Casolaro, V.; Bjorck, L.; Marone, G.; Protein, L. A Bacterial Ig-Binding Protein That Activates Human Basophils and Mast Cells. J. Immunol. 1990, 145, 3054–3061. [Google Scholar]
- Genovese, A.; Bouvet, J.P.; Florio, G.; Lamparter-Schummert, B.; Bjorck, L.; Marone, G. Bacterial Immunoglobulin Superantigen Proteins a and L Activate Human Heart Mast Cells by Interacting with Immunoglobulin E. Infect Immun. 2000, 68, 5517–5524. [Google Scholar] [CrossRef]
- Genovese, A.; Borgia, G.; Bjorck, L.; Petraroli, A.; de Paulis, A.; Piazza, M.; Marone, G. Immunoglobulin Superantigen Protein L Induces Il-4 and Il-13 Secretion from Human Fc Epsilon Ri+ Cells through Interaction with the Kappa Light Chains of Ige. J. Immunol. 2003, 170, 1854–1861. [Google Scholar] [CrossRef]
- Nilson, B.H.; Solomon, A.; Bjorck, L.; Akerstrom, B. Protein L from Peptostreptococcus Magnus Binds to the Kappa Light Chain Variable Domain. J. Biol. Chem. 1992, 267, 2234–2239. [Google Scholar] [CrossRef]
- Marone, G.; Rossi, F.W.; Detoraki, A.; Granata, F.; Genovese, A.; Spadaro, G. Role of Superallergens in Allergic Disorders. Chem. Immunol. Allergy 2007, 93, 195–213. [Google Scholar]
- Rha, M.S.; Kim, S.W.; Chang, D.Y.; Lee, J.K.; Kim, J.; Park, S.H.; Khalmuratova, R.; Lim, H.S.; Eun, K.M.; Hong, S.N.; et al. Superantigen-Related Th2 Cd4(+) T Cells in Nonasthmatic Chronic Rhinosinusitis with Nasal Polyps. J. Allergy Clin. Immunol. 2020, 145, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Teufelberger, A.R.; Broker, B.M.; Krysko, D.V.; Bachert, C.; Krysko, O. Staphylococcus Aureus Orchestrates Type 2 Airway Diseases. Trends Mol. Med. 2019, 25, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Tsilochristou, O.; Toit, G.d.; Sayre, P.H.; Roberts, G.; Lawson, K.; Sever, M.L.; Bahnson, H.T.; Radulovic, S.; Basting, M.; Plaut, M.; et al. Association of Staphylococcus Aureus Colonization with Food Allergy Occurs Independently of Eczema Severity. J. Allergy Clin. Immunol. 2019, 144, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Bachert, C.; Gevaert, P.; Zhang, N.; van Zele, T.; Perez-Novo, C. Role of Staphylococcal Superantigens in Airway Disease. Chem. Immunol. Allergy 2007, 93, 214–236. [Google Scholar] [PubMed]
- Kim, Y.C.; Won, H.K.; Lee, J.W.; Sohn, K.H.; Kim, M.H.; Kim, T.B.; Chang, Y.S.; Lee, B.J.; Cho, S.H.; Bachert, C.; et al. Staphylococcus Aureus Nasal Colonization and Asthma in Adults: Systematic Review and Meta-Analysis. J. Allergy Clin. Immunol. Pract. 2019, 7, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, J.; Lu, Y.W.; Wu, S.; Wang, M.R.; Zhu, J.M. Possible Role of Staphylococcal Enterotoxin B in the Pathogenesis of Autoimmune Diseases. Viral Immunol. 2015, 28, 354–359. [Google Scholar] [CrossRef]
- Viau, M.; Zouali, M. B-Lymphocytes, Innate Immunity, and Autoimmunity. Clin. Immunol. 2005, 114, 17–26. [Google Scholar] [CrossRef]
- He, Y.; Sun, Y.; Ren, Y.; Qiao, L.; Guo, R.; Du, J.; Zhu, X.; Liu, Y.; Lin, J. The T Cell Activating Properties and Antitumour Activity of Staphylococcal Enterotoxin-Like Q. Med. Microbiol. Immunol. 2019, 208, 781–792. [Google Scholar] [CrossRef]
- Golob-Urbanc, A.; Rajcevic, U.; Strmsek, Z.; Jerala, R. Design of Split Superantigen Fusion Proteins for Cancer Immunotherapy. J. Biol. Chem. 2019, 294, 6294–6305. [Google Scholar] [CrossRef]
- Agheli, R.; Emkanian, B.; Halabian, R.; Mehrabadi, J.F.; Fooladi, A.A.I. Recombinant Staphylococcal Enterotoxin Type a Stimulate Antitumoral Cytokines. Technol. Cancer Res. Treat 2017, 16, 125–132. [Google Scholar] [CrossRef]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. Il-33, an Interleukin-1-Like Cytokine That Signals Via the Il-1 Receptor-Related Protein St2 and Induces T Helper Type 2-Associated Cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J.P. Interleukin-33 (Il-33): A Nuclear Cytokine from the Il-1 Family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Italiani, P.; Weil, S.; Martin, M.U. The Family of the Interleukin-1 Receptors. Immunol. Rev. 2018, 281, 197–232. [Google Scholar] [PubMed]
- Martin, N.T.; Martin, M.U. Interleukin 33 Is a Guardian of Barriers and a Local Alarmin. Nat. Immunol. 2016, 17, 122–131. [Google Scholar] [PubMed]
- Moussion, C.; Ortega, N.; Girard, J.P. The Il-1-Like Cytokine Il-33 Is Constitutively Expressed in the Nucleus of Endothelial Cells and Epithelial Cells in Vivo: A Novel ‘Alarmin’? PLoS ONE 2008, 3, e3331. [Google Scholar] [CrossRef]
- Afferni, C.; Buccione, C.; Andreone, S.; Galdiero, M.R.; Varricchi, G.; Marone, G.; Mattei, F.; Schiavoni, G. The Pleiotropic Immunomodulatory Functions of Il-33 and Its Implications in Tumor Immunity. Front. Immunol. 2018, 9, 2601. [Google Scholar] [CrossRef]
- Drake, L.Y.; Kita, H. Il-33: Biological Properties, Functions, and Roles in Airway Disease. Immunol. Rev. 2017, 278, 173–184. [Google Scholar] [CrossRef]
- Babina, M.; Wang, Z.; Franke, K.; Guhl, S.; Artuc, M.; Zuberbier, T. Yin-Yang of Il-33 in Human Skin Mast Cells: Reduced Degranulation, but Augmented Histamine Synthesis through P38 Activation. J. Investig. Dermatol. 2019, 139, 1516–1525. [Google Scholar] [CrossRef]
- Joulia, R.; L’Faqihi, F.E.; Valitutti, S.; Espinosa, E. Il-33 Fine Tunes Mast Cell Degranulation and Chemokine Production at the Single-Cell Level. J. Allergy Clin. Immunol. 2017, 140, 497–509. [Google Scholar] [CrossRef]
- Morita, H.; Arae, K.; Unno, H.; Miyauchi, K.; Toyama, S.; Nambu, A.; Oboki, K.; Ohno, T.; Motomura, K.; Matsuda, A.; et al. An Interleukin-33-Mast Cell-Interleukin-2 Axis Suppresses Papain-Induced Allergic Inflammation by Promoting Regulatory T Cell Numbers. Immunity 2015, 43, 175–186. [Google Scholar] [CrossRef]
- Taracanova, A.; Alevizos, M.; Karagkouni, A.; Weng, Z.; Norwitz, E.; Conti, P.; Leeman, S.E.; Theoharides, T.C. Sp and Il-33 Together Markedly Enhance Tnf Synthesis and Secretion from Human Mast Cells Mediated by the Interaction of Their Receptors. Proc. Natl. Acad. Sci. USA 2017, 114, E4002–E4009. [Google Scholar] [CrossRef] [PubMed]
- Bandara, G.; Beaven, M.A.; Olivera, A.; Gilfillan, A.M.; Metcalfe, D.D. Activated Mast Cells Synthesize and Release Soluble St2-a Decoy Receptor for Il-33. Eur. J. Immunol. 2015, 45, 3034–3044. [Google Scholar] [CrossRef] [PubMed]
- Salamon, P.; Shefler, I.; Moshkovits, I.; Munitz, A.; Klotzman, D.H.; Mekori, Y.A.; Hershko, A.Y. Il-33 and Ige Stimulate Mast Cell Production of Il-2 and Regulatory T Cell Expansion in Allergic Dermatitis. Clin. Exp. Allergy 2017, 47, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Guhl, S.; Franke, K.; Artuc, M.; Zuberbier, T.; Babina, M. Il-33 and Mrgprx2-Triggered Activation of Human Skin Mast Cells-Elimination of Receptor Expression on Chronic Exposure, but Reinforced Degranulation on Acute Priming. Cells 2019, 8, 341. [Google Scholar] [CrossRef]
- Tjota, M.Y.; Williams, J.W.; Lu, T.; Clay, B.S.; Byrd, T.; Hrusch, C.L.; Decker, D.C.; de Araujo, C.A.; Bryce, P.J.; Sperling, A.I. Il-33-Dependent Induction of Allergic Lung Inflammation by Fcgammariii Signaling. J. Clin. Investig. 2013, 123, 2287–2297. [Google Scholar] [CrossRef]
- Loxham, M.; Davies, D.E. Phenotypic and Genetic Aspects of Epithelial Barrier Function in Asthmatic Patients. J. Allergy Clin. Immunol. 2017, 139, 1736–1751. [Google Scholar] [CrossRef]
- Cayrol, C.; Duval, A.; Schmitt, P.; Roga, S.; Camus, M.; Stella, A.; Burlet-Schiltz, O.; Gonzalez-de-Peredo, A.; Girard, J.P. Environmental Allergens Induce Allergic Inflammation through Proteolytic Maturation of Il-33. Nat. Immunol. 2018, 19, 375–385. [Google Scholar] [CrossRef]
- Altman, M.C.; Lai, Y.; Nolin, J.D.; Long, S.; Chen, C.C.; Piliponsky, A.M.; Altemeier, W.A.; Larmore, M.; Frevert, C.W.; Mulligan, M.S.; et al. Airway Epithelium-Shifted Mast Cell Infiltration Regulates Asthmatic Inflammation Via Il-33 Signaling. J. Clin. Investig. 2019, 129, 4979–4991. [Google Scholar] [CrossRef]
- Werder, R.B.; Zhang, V.; Lynch, J.P.; Snape, N.; Upham, J.W.; Spann, K.; Phipps, S. Chronic Il-33 Expression Predisposes to Virus-Induced Asthma Exacerbations by Increasing Type 2 Inflammation and Dampening Antiviral Immunity. J. Allergy Clin. Immunol. 2018, 141, 1607–1619. [Google Scholar] [CrossRef]
- Alves-Filho, J.C.; Sonego, F.; Souto, F.O.; Freitas, A.; Verri, W.A.; Auxiliadora-Martins, M., Jr.; Basile-Filho, A.; McKenzie, A.N.; Xu, D.; Cunha, F.Q.; et al. Interleukin-33 Attenuates Sepsis by Enhancing Neutrophil Influx to the Site of Infection. Nat. Med. 2010, 16, 708–712. [Google Scholar] [CrossRef]
- Kearley, J.; Silver, J.S.; Sanden, C.; Liu, Z.; Berlin, A.A.; White, N.; Mori, M.; Pham, T.H.; Ward, C.K.; Criner, G.J.; et al. Cigarette Smoke Silences Innate Lymphoid Cell Function and Facilitates an Exacerbated Type I Interleukin-33-Dependent Response to Infection. Immunity 2015, 42, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Zizzo, G.; Cohen, P.L. Imperfect Storm: Is Interleukin-33 the Achilles Heel of Covid-19? Lancet Rheumatol. 2020, 2, e779–e790. [Google Scholar] [CrossRef]
- Liew, F.Y.; Girard, J.P.; Turnquist, H.R. Interleukin-33 in Health and Disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Hollande, C.; Boussier, J.; Ziai, J.; Nozawa, T.; Bondet, V.; Phung, W.; Lu, B.; Duffy, D.; Paradis, V.; Mallet, V.; et al. Inhibition of the Dipeptidyl Peptidase Dpp4 (Cd26) Reveals Il-33-Dependent Eosinophil-Mediated Control of Tumor Growth. Nat. Immunol. 2019, 20, 257–264. [Google Scholar] [CrossRef]
- Marone, G.; Casolaro, V.; Paganelli, R.; Quinti, I. Igg Anti-Ige from Atopic Dermatitis Induces Mediator Release from Basophils and Mast Cells. J. Investig. Dermatol. 1989, 93, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Spadaro, G.; Palumbo, C.; Condorelli, G. The Anti-Ige/Anti-Fcepsilonrialpha Autoantibody Network in Allergic and Autoimmune Diseases. Clin. Exp. Allergy 1999, 29, 17–27. [Google Scholar] [CrossRef]
- Patella, V.; Marino, I.; Arbustini, E.; Lamparter-Schummert, B.; Verga, L.; Adt, M.; Marone, G. Stem Cell Factor in Mast Cells and Increased Mast Cell Density in Idiopathic and Ischemic Cardiomyopathy. Circulation 1998, 97, 971–978. [Google Scholar] [CrossRef]
- Patella, V.; Giuliano, A.; Bouvet, J.P.; Marone, G. Endogenous Superallergen Protein Fv Induces Il-4 Secretion from Human Fc Epsilon Ri+ Cells through Interaction with the Vh3 Region of Ige. J. Immunol. 1998, 161, 5647–5655. [Google Scholar]
- Marone, G.; Tamburini, M.; Giudizi, M.G.; Biagiotti, R.; Almerigogna, F.; Romagnani, S. Mechanism of Activation of Human Basophils by Staphylococcus Aureus Cowan 1. Infect Immun. 1987, 55, 803–809. [Google Scholar] [CrossRef]
- Sala, P.; Tonutti, E.; Ruscio, M.; Colle, R.; Antonutto, G.; Falconieri, G. Ige Myeloma. Report of a New Case and Review of the Literature. Haematologica 1981, 66, 787–795. [Google Scholar]
- Patella, V.; de Crescenzo, G.; Marino, I.; Genovese, A.; Adt, M.; Gleich, G.J.; Marone, G. Eosinophil Granule Proteins Activate Human Heart Mast Cells. J. Immunol. 1996, 157, 1219–1225. [Google Scholar] [PubMed]
- Siraganian, R.P. An Automated Continuous-Flow System for the Extraction and Fluorometric Analysis of Histamine. Anal. Biochem. 1974, 57, 383–394. [Google Scholar] [CrossRef]
- Loffredo, S.; Ferrara, A.L.; Bova, M.; Borriello, F.; Suffritti, C.; Veszeli, N.; Petraroli, A.; Galdiero, M.R.; Varricchi, G.; Granata, F.; et al. Secreted Phospholipases A2 in Hereditary Angioedema with C1-Inhibitor Deficiency. Front. Immunol. 2018, 9, 1721. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Rossi, F.W.; Pecoraro, A.; Pucino, V.; Criscuolo, G.; Paulis, A.; Spadaro, G.; Varricchi, G. Hiv Gp120 Induces the Release of Proinflammatory, Angiogenic, and Lymphangiogenic Factors from Human Lung Mast Cells. Vaccines (Basel) 2020, 8, 208. [Google Scholar] [CrossRef] [PubMed]
- Stentzel, S.; Teufelberger, A.; Nordengrun, M.; Kolata, J.; Schmidt, F.; van Crombruggen, K.; Michalik, S.; Kumpfmuller, J.; Tischer, S.; Schweder, T.; et al. Staphylococcal Serine Protease-Like Proteins Are Pacemakers of Allergic Airway Reactions to Staphylococcus Aureus. J. Allergy Clin. Immunol. 2017, 139, 492–500. [Google Scholar] [CrossRef]
- Huvenne, W.; Callebaut, I.; Plantinga, M.; Vanoirbeek, J.A.; Krysko, O.; Bullens, D.M.; Gevaert, P.; van Cauwenberge, P.; Lambrecht, B.N.; Ceuppens, J.L.; et al. Staphylococcus Aureus Enterotoxin B Facilitates Allergic Sensitization in Experimental Asthma. Clin. Exp. Allergy 2010, 40, 1079–1090. [Google Scholar] [CrossRef]
- Rocha-de-Souza, C.M.; Berent-Maoz, B.; Mankuta, D.; Moses, A.E.; Levi-Schaffer, F. Human Mast Cell Activation by Staphylococcus Aureus: Interleukin-8 and Tumor Necrosis Factor Alpha Release and the Role of Toll-Like Receptor 2 and Cd48 Molecules. Infect. Immun. 2008, 76, 4489–4497. [Google Scholar] [CrossRef]
- Varricchi, G.; Loffredo, S.; Borriello, F.; Pecoraro, A.; Rivellese, F.; Genovese, A.; Spadaro, G.; Marone, G. Superantigenic Activation of Human Cardiac Mast Cells. Int. J. Mol. Sci. 2019, 20, 1828. [Google Scholar] [CrossRef]
- Kaur, D.; Gomez, E.; Doe, C.; Berair, R.; Woodman, L.; Saunders, R.; Hollins, F.; Rose, F.R.; Amrani, Y.; May, R.; et al. Il-33 Drives Airway Hyper-Responsiveness through Il-13-Mediated Mast Cell: Airway Smooth Muscle Crosstalk. Allergy 2015, 70, 556–567. [Google Scholar] [CrossRef]
- Ho, L.H.; Ohno, T.; Oboki, K.; Kajiwara, N.; Suto, H.; Iikura, M.; Okayama, Y.; Akira, S.; Saito, H.; Galli, S.J.; et al. Il-33 Induces Il-13 Production by Mouse Mast Cells Independently of Ige-Fcepsilonri Signals. J. Leukoc Biol. 2007, 82, 1481–1490. [Google Scholar] [CrossRef]
- Kashiwakura, J.; Yanagisawa, M.; Lee, H.; Okamura, Y.; Sasaki-Sakamoto, T.; Saito, S.; Tokuhashi, Y.; Ra, C.; Okayama, Y. Interleukin-33 Synergistically Enhances Immune Complex-Induced Tumor Necrosis Factor Alpha and Interleukin-8 Production in Cultured Human Synovium-Derived Mast Cells. Int. Arch. Allergy Immunol. 2013, 161 (Suppl. 2), 32–36. [Google Scholar] [CrossRef] [PubMed]
- Myhre, E.B.; Erntell, M. A Non-Immune Interaction between the Light Chain of Human Immunoglobulin and a Surface Component of a Peptococcus Magnus Strain. Mol. Immunol. 1985, 22, 879–885. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.C.; Wenzel, S.E. Intersection of Biology and Therapeutics: Type 2 Targeted Therapeutics for Adult Asthma. Lancet 2020, 395, 371–383. [Google Scholar] [CrossRef]
- Maun, H.R.; Jackman, J.K.; Choy, D.F.; Loyet, K.M.; Staton, T.L.; Jia, G.; Dressen, A.; Hackney, J.A.; Bremer, M.; Walters, B.T.; et al. An Allosteric Anti-Tryptase Antibody for the Treatment of Mast Cell-Mediated Severe Asthma. Cell 2019, 179, 417–431. [Google Scholar] [CrossRef]
- Bigna, J.J.; Kenne, A.M.; Asangbeh, S.L.; Sibetcheu, A.T. Prevalence of Chronic Obstructive Pulmonary Disease in the Global Population with Hiv: A Systematic Review and Meta-Analysis. Lancet Glob. Health 2018, 6, e193–e202. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Bjermer, L. Mast Cell-Mediated Orchestration of the Immune Responses in Human Allergic Asthma: Current Insights. Clin. Rev. Allergy Immunol. 2019, 56, 234–247. [Google Scholar] [CrossRef]
- Salamon, P.; Mekori, Y.A.; Shefler, I. Lung Cancer-Derived Extracellular Vesicles: A Possible Mediator of Mast Cell Activation in the Tumor Microenvironment. Cancer Immunol. Immunother. 2020, 69, 373–381. [Google Scholar] [CrossRef]
- McDowell, P.J.; Heaney, L.G. Different Endotypes and Phenotypes Drive the Heterogeneity in Severe Asthma. Allergy 2020, 75, 302–310. [Google Scholar] [CrossRef]
- Sze, E.; Bhalla, A.; Nair, P. Mechanisms and Therapeutic Strategies for Non-T2 Asthma. Allergy 2020, 75, 311–325. [Google Scholar] [CrossRef]
- Samitas, K.; Zervas, E.; Gaga, M. T2-Low Asthma: Current Approach to Diagnosis and Therapy. Curr. Opin. Pulm. Med. 2017, 23, 48–55. [Google Scholar] [CrossRef]
- Davis, M.F.; Peng, R.D.; McCormack, M.C.; Matsui, E.C. Staphylococcus Aureus Colonization Is Associated with Wheeze and Asthma among Us Children and Young Adults. J. Allergy Clin. Immunol. 2015, 135, 811–813. [Google Scholar] [CrossRef]
- Bachert, C.; van Steen, K.; Zhang, N.; Holtappels, G.; Cattaert, T.; Maus, B.; Buhl, R.; Taube, C.; Korn, S.; Kowalski, M.; et al. Specific Ige against Staphylococcus Aureus Enterotoxins: An Independent Risk Factor for Asthma. J. Allergy Clin. Immunol. 2012, 130, 376–381. [Google Scholar] [CrossRef]
- Alitalo, K. The Lymphatic Vasculature in Disease. Nat. Med. 2011, 17, 1371–1380. [Google Scholar] [CrossRef]
- Brakenhielm, E.; Alitalo, K. Cardiac Lymphatics in Health and Disease. Nat. Rev. Cardiol. 2019, 16, 56–68. [Google Scholar] [CrossRef]
- Bachert, C.; Zhang, N.; Holtappels, G.; de Lobel, L.; van Cauwenberge, P.; Liu, S.; Lin, P.; Bousquet, J.; van Steen, K. Presence of Il-5 Protein and Ige Antibodies to Staphylococcal Enterotoxins in Nasal Polyps Is Associated with Comorbid Asthma. J. Allergy Clin. Immunol. 2010, 126, 962–968.e6. [Google Scholar] [CrossRef]
- Aman, M.J. Superantigens of a Superbug: Major Culprits of Staphylococcus Aureus Disease? Virulence 2017, 8, 607–610. [Google Scholar] [CrossRef][Green Version]
- Teufelberger, A.R.; Nordengrun, M.; Braun, H.; Maes, T.; de Grove, K.; Holtappels, G.; O’Brien, C.; Provoost, S.; Hammad, H.; Goncalves, A.; et al. The Il-33/St2 Axis Is Crucial in Type 2 Airway Responses Induced by Staphylococcus Aureus-Derived Serine Protease-Like Protein D. J. Allergy Clin. Immunol. 2018, 141, 549–559. [Google Scholar] [CrossRef]
- Bachert, C.; Claeys, S.E.; Tomassen, P.; van Zele, T.; Zhang, N. Rhinosinusitis and Asthma: A Link for Asthma Severity. Curr. Allergy Asthma Rep. 2010, 10, 194–201. [Google Scholar] [CrossRef]
- Tomassen, P.; Vandeplas, G.; van Zele, T.; Cardell, L.O.; Arebro, J.; Olze, H.; Forster-Ruhrmann, U.; Kowalski, M.L.; Olszewska-Ziaber, A.; Holtappels, G.; et al. Inflammatory Endotypes of Chronic Rhinosinusitis Based on Cluster Analysis of Biomarkers. J. Allergy Clin. Immunol. 2016, 137, 1449–1456. [Google Scholar] [CrossRef]
- Vigorito, C.; Giordano, A.; Cirillo, R.; Genovese, A.; Rengo, F.; Marone, G. Metabolic and Hemodynamic Effects of Peptide Leukotriene C4 and D4 in Man. Int. J. Clin. Lab. Res. 1997, 27, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Galdiero, M.R.; Tocchetti, C.G. Cardiac Toxicity of Immune Checkpoint Inhibitors: Cardio-Oncology Meets Immunology. Circulation 2017, 136, 1989–1992. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Marone, G.; Kovanen, P.T. Cardiac Mast Cells: Underappreciated Immune Cells in Cardiovascular Homeostasis and Disease. Trends Immunol. 2020, 41, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.F.; Thompson, L.J.; Ziegler, S.F. Tslp Drives Acute Th2-Cell Differentiation in Lungs. J. Allergy Clin. Immunol. 2020, 146, 1406. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Spadaro, G.; Braile, M.; Poto, R.; Criscuolo, G.; Pahima, H.; Loffredo, S.; Levi-Schaffer, F.; Varricchi, G. Tezepelumab: A Novel Biological Therapy for the Treatment of Severe Uncontrolled Asthma. Expert Opin. Investig. Drugs 2019, 28, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Pecoraro, A.; Marone, G.; Criscuolo, G.; Spadaro, G.; Genovese, A. Thymic Stromal Lymphopoietin Isoforms, Inflammatory Disorders, and Cancer. Front. Immunol. 2018, 9, 1595. [Google Scholar] [CrossRef]
- Chu, D.K.; Llop-Guevara, A.; Walker, T.D.; Flader, K.; Goncharova, S.; Boudreau, J.E.; Moore, C.L.; In, T.S.; Waserman, S.; Coyle, A.J.; et al. Il-33, but Not Thymic Stromal Lymphopoietin or Il-25, Is Central to Mite and Peanut Allergic Sensitization. J. Allergy Clin. Immunol. 2013, 131, 187–200.e1-8. [Google Scholar] [CrossRef]
- Iijima, K.; Kobayashi, T.; Hara, K.; Kephart, G.M.; Ziegler, S.F.; McKenzie, A.N.; Kita, H. Il-33 and Thymic Stromal Lymphopoietin Mediate Immune Pathology in Response to Chronic Airborne Allergen Exposure. J. Immunol. 2014, 193, 1549–1559. [Google Scholar] [CrossRef]
- Prefontaine, D.; Nadigel, J.; Chouiali, F.; Audusseau, S.; Semlali, A.; Chakir, J.; Martin, J.G.; Hamid, Q. Increased Il-33 Expression by Epithelial Cells in Bronchial Asthma. J. Allergy Clin. Immunol. 2010, 125, 752–754. [Google Scholar] [CrossRef]
- Beale, J.; Jayaraman, A.; Jackson, D.J.; Macintyre, J.D.R.; Edwards, M.R.; Walton, R.P.; Zhu, J.; Ching, Y.M.; Shamji, B.; Edwards, M.; et al. Rhinovirus-Induced Il-25 in Asthma Exacerbation Drives Type 2 Immunity and Allergic Pulmonary Inflammation. Sci. Transl. Med. 2014, 6, 256ra134. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. The Immunology of Asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; O’Connor, B.; Ratoff, J.; Meng, Q.; Mallett, K.; Cousins, D.; Robinson, D.; Zhang, G.; Zhao, J.; Lee, T.H.; et al. Thymic Stromal Lymphopoietin Expression Is Increased in Asthmatic Airways and Correlates with Expression of Th2-Attracting Chemokines and Disease Severity. J. Immunol. 2005, 174, 8183–8190. [Google Scholar] [CrossRef] [PubMed]
- Enoksson, M.; Lyberg, K.; Moller-Westerberg, C.; Fallon, P.G.; Nilsson, G.; Lunderius-Andersson, C. Mast Cells as Sensors of Cell Injury through Il-33 Recognition. J. Immunol. 2011, 186, 2523–2528. [Google Scholar] [CrossRef] [PubMed]
- Tatari, N.; Movassagh, H.; Shan, L.; Koussih, L.; Gounni, A.S. Semaphorin 3e Inhibits House Dust Mite-Induced Angiogenesis in a Mouse Model of Allergic Asthma. Am. J. Pathol. 2019, 189, 762–772. [Google Scholar] [CrossRef]
- Chatterjee, S.; Heukamp, L.C.; Siobal, M.; Schottle, J.; Wieczorek, C.; Peifer, M.; Frasca, D.; Koker, M.; Konig, K.; Meder, L.; et al. Tumor Vegf:Vegfr2 Autocrine Feed-Forward Loop Triggers Angiogenesis in Lung Cancer. J. Clin. Investig. 2013, 123, 1732–1740. [Google Scholar] [CrossRef]
- Bosisio, D.; Ronca, R.; Salvi, V.; Presta, M.; Sozzani, S. Dendritic Cells in Inflammatory Angiogenesis and Lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 180–186. [Google Scholar] [CrossRef]
- Wilson, A.M.; Shao, Z.; Grenier, V.; Mawambo, G.; Daudelin, J.F.; Dejda, A.; Pilon, F.; Popovic, N.; Boulet, S.; Parinot, C.; et al. Neuropilin-1 Expression in Adipose Tissue Macrophages Protects against Obesity and Metabolic Syndrome. Sci. Immunol. 2018, 3, eaan4626. [Google Scholar] [CrossRef]
- Stump, B.; Cui, Y.; Kidambi, P.; Lamattina, A.M.; El-Chemaly, S. Lymphatic Changes in Respiratory Diseases: More Than Just Remodeling of the Lung? Am. J. Respir. Cell Mol. Biol. 2017, 57, 272–279. [Google Scholar] [CrossRef]
- Yao, L.C.; Baluk, P.; Feng, J.; McDonald, D.M. Steroid-Resistant Lymphatic Remodeling in Chronically Inflamed Mouse Airways. Am. J. Pathol. 2010, 176, 1525–1541. [Google Scholar] [CrossRef]
- Hardavella, G.; Tzortzaki, E.G.; Siozopoulou, V.; Galanis, P.; Vlachaki, E.; Avgousti, M.; Stefanou, D.; Siafakas, N.M. Lymphangiogenesis in Copd: Another Link in the Pathogenesis of the Disease. Respir. Med. 2012, 106, 687–693. [Google Scholar] [CrossRef]
- Mori, M.; Andersson, C.K.; Graham, G.J.; Lofdahl, C.G.; Erjefalt, J.S. Increased Number and Altered Phenotype of Lymphatic Vessels in Peripheral Lung Compartments of Patients with Copd. Respir. Res. 2013, 14, 65. [Google Scholar] [CrossRef]
- Aspelund, A.; Robciuc, M.R.; Karaman, S.; Makinen, T.; Alitalo, K. Lymphatic System in Cardiovascular Medicine. Circ. Res. 2016, 118, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; de Paulis, A.; Marone, G.; Galli, S.J. Future Needs in Mast Cell Biology. Int. J. Mol. Sci. 2019, 20, 4397. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Aspelund, A.; Alitalo, K. Lymphangiogenic Factors, Mechanisms, and Applications. J. Clin. Investig. 2014, 124, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kataru, R.P.; Koh, G.Y. Inflammation-Associated Lymphangiogenesis: A Double-Edged Sword? J. Clin. Investig. 2014, 124, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Nath, S.; Meininger, C.J.; Gashev, A.A. Emerging Roles of Mast Cells in the Regulation of Lymphatic Immuno-Physiology. Front. Immunol. 2020, 11, 1234. [Google Scholar] [CrossRef]
- Sammarco, G.; Varricchi, G.; Ferraro, V.; Ammendola, M.; de Fazio, M.; Altomare, D.F.; Luposella, M.; Maltese, L.; Curro, G.; Marone, G.; et al. Mast Cells, Angiogenesis and Lymphangiogenesis in Human Gastric Cancer. Int. J. Mol. Sci. 2019, 20, 2106. [Google Scholar] [CrossRef]
- Fankhauser, M.; Broggi, M.A.S.; Potin, L.; Bordry, N.; Jeanbart, L.; Lund, A.W.; da Costa, E.; Hauert, S.; Rincon-Restrepo, M.; Tremblay, C.; et al. Tumor Lymphangiogenesis Promotes T Cell Infiltration and Potentiates Immunotherapy in Melanoma. Sci. Transl. Med. 2017, 9, eaal4712. [Google Scholar] [CrossRef]
- Henri, O.; Pouehe, C.; Houssari, M.; Galas, L.; Nicol, L.; Edwards-Levy, F.; Henry, J.P.; Dumesnil, A.; Boukhalfa, I.; Banquet, S.; et al. Selective Stimulation of Cardiac Lymphangiogenesis Reduces Myocardial Edema and Fibrosis Leading to Improved Cardiac Function Following Myocardial Infarction. Circulation 2016, 133, 1484–1497. [Google Scholar] [CrossRef]
- Klotz, L.; Norman, S.; Vieira, J.M.; Masters, M.; Rohling, M.; Dube, K.N.; Bollini, S.; Matsuzaki, F.; Carr, C.A.; Riley, P.R. Cardiac Lymphatics Are Heterogeneous in Origin and Respond to Injury. Nature 2015, 522, 62–67. [Google Scholar] [CrossRef]
- Huang, L.H.; Lavine, K.J.; Randolph, G.J. Cardiac Lymphatic Vessels, Transport, and Healing of the Infarcted Heart. JACC Basic Transl. Sci. 2017, 2, 477–483. [Google Scholar] [CrossRef]
- Vieira, J.M.; Norman, S.; del Campo, C.V.; Cahill, T.J.; Barnette, D.N.; Gunadasa-Rohling, M.; Johnson, L.A.; Greaves, D.R.; Carr, C.A.; Jackson, D.G.; et al. The Cardiac Lymphatic System Stimulates Resolution of Inflammation Following Myocardial Infarction. J. Clin. Investig. 2018, 128, 3402–3412. [Google Scholar] [CrossRef] [PubMed]
- Hallstrand, T.S.; Hackett, T.L.; Altemeier, W.A.; Matute-Bello, G.; Hansbro, P.M.; Knight, D.A. Airway Epithelial Regulation of Pulmonary Immune Homeostasis and Inflammation. Clin. Immunol. 2014, 151, 1–15. [Google Scholar] [CrossRef]
- Kuchuk, O.; Tuccitto, A.; Citterio, D.; Huber, V.; Camisaschi, C.; Milione, M.; Vergani, B.; Villa, A.; Alison, M.R.; Carradori, S.; et al. Ph Regulators to Target the Tumor Immune Microenvironment in Human Hepatocellular Carcinoma. Oncoimmunology 2018, 7, e1445452. [Google Scholar] [CrossRef] [PubMed]
- Gulliksson, M.; Carvalho, R.F.; Ulleras, E.; Nilsson, G. Mast Cell Survival and Mediator Secretion in Response to Hypoxia. PLoS ONE 2010, 5, e12360. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Moreno, I.G.; Ibarra-Sanchez, A.; Castillo-Arellano, J.I.; Blank, U.; Gonzalez-Espinosa, C. Mast Cells Localize in Hypoxic Zones of Tumors and Secrete Ccl-2 under Hypoxia through Activation of L-Type Calcium Channels. J. Immunol. 2020, 204, 1056–1068. [Google Scholar] [CrossRef]
- Abebayehu, D.; Spence, A.J.; Qayum, A.A.; Taruselli, M.T.; McLeod, J.J.; Caslin, H.L.; Chumanevich, A.P.; Kolawole, E.M.; Paranjape, A.; Baker, B.; et al. Lactic Acid Suppresses Il-33-Mediated Mast Cell Inflammatory Responses Via Hypoxia-Inducible Factor-1alpha-Dependent Mir-155 Suppression. J. Immunol. 2016, 197, 2909–2917. [Google Scholar] [CrossRef] [PubMed]
- Gorzalczany, Y.; Akiva, E.; Klein, O.; Merimsky, O.; Sagi-Eisenberg, R. Mast Cells Are Directly Activated by Contact with Cancer Cells by a Mechanism Involving Autocrine Formation of Adenosine and Autocrine/Paracrine Signaling of the Adenosine A3 Receptor. Cancer Lett. 2017, 397, 23–32. [Google Scholar] [CrossRef]
- Cekic, C.; Linden, J. Purinergic Regulation of the Immune System. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monoclonal Antibody (mAb) Anti-FcεRI (μg/mL) | ||||
|---|---|---|---|---|
| 10−1 | 1 | 3 | ||
| Percent Histamine Release | 14.0 ± 3.21 | 23.33 ± 4.80 | 24.66 ± 3.84 | |
| VEGF-A (pg/106 cells) | 16.33 ± 3.28 | 28.66 ± 1.45 | 51.33 ± 3.84 | |
| VEGF-C (pg/106 cells) | 16.33 ± 4.91 | 29.33 ± 6.43 | 45.0 ± 8.14 | |
| Human Polyclonal IgG (μg/mL) | ||||
|---|---|---|---|---|
| 10−2 | 10−1 | 1 | 3 | |
| Percent Histamine Release | 1.66 ± 1.20 | 1.66 ± 0.88 | 1.33 ± 0.88 | 1.83 ± 1.01 |
| VEGF-A (pg/106 cells) | 0.33 ± 0.33 | 2.30 ± 0.33 | 1.66 ± 1.20 | 0.66 ± 0.66 |
| VEGF-C (pg/106 cells) | 1.16 ± 0.60 | 0.50 ± 0.50 | 1.4 ± 0.94 | 0.83 ± 0.44 |
| Stimulus | Percent Histamine Release |
|---|---|
| Protein A | 18.3 ± 0.9 |
| IgM VH3+ | 0.3 ± 0.3 |
| IgM VH3+ + Protein A | 3.0 ± 0.6 *** |
| IgM VH6+ | 0.7 ± 0.6 |
| IgM VH6+ + Protein A | 18.7 ± 0.3 |
| Stimulus | Percent Histamine Release |
|---|---|
| Protein L (100 nM) | 19.0 ± 1.5 |
| IgE λ (0.3 μg/mL) + Protein L | 18.7 ± 1.8 |
| IgE λ (1 μg/mL) + Protein L | 18.3 ± 2.0 |
| IgE κ (0.3 μg/mL) + Protein L | 13.7 ± 0.7 * |
| IgE κ (1 μg/mL) + Protein L | 3.7 ± 1.2 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristinziano, L.; Poto, R.; Criscuolo, G.; Ferrara, A.L.; Galdiero, M.R.; Modestino, L.; Loffredo, S.; de Paulis, A.; Marone, G.; Spadaro, G.; et al. IL-33 and Superantigenic Activation of Human Lung Mast Cells Induce the Release of Angiogenic and Lymphangiogenic Factors. Cells 2021, 10, 145. https://doi.org/10.3390/cells10010145
Cristinziano L, Poto R, Criscuolo G, Ferrara AL, Galdiero MR, Modestino L, Loffredo S, de Paulis A, Marone G, Spadaro G, et al. IL-33 and Superantigenic Activation of Human Lung Mast Cells Induce the Release of Angiogenic and Lymphangiogenic Factors. Cells. 2021; 10(1):145. https://doi.org/10.3390/cells10010145
Chicago/Turabian StyleCristinziano, Leonardo, Remo Poto, Gjada Criscuolo, Anne Lise Ferrara, Maria Rosaria Galdiero, Luca Modestino, Stefania Loffredo, Amato de Paulis, Gianni Marone, Giuseppe Spadaro, and et al. 2021. "IL-33 and Superantigenic Activation of Human Lung Mast Cells Induce the Release of Angiogenic and Lymphangiogenic Factors" Cells 10, no. 1: 145. https://doi.org/10.3390/cells10010145
APA StyleCristinziano, L., Poto, R., Criscuolo, G., Ferrara, A. L., Galdiero, M. R., Modestino, L., Loffredo, S., de Paulis, A., Marone, G., Spadaro, G., & Varricchi, G. (2021). IL-33 and Superantigenic Activation of Human Lung Mast Cells Induce the Release of Angiogenic and Lymphangiogenic Factors. Cells, 10(1), 145. https://doi.org/10.3390/cells10010145