1. Introduction
Winter rapeseed (
Brassica napus L.) is the most important oil crop in Europe. In recent years (2015–2017), this species was cultivated in the area of 8.1–8.8 mln ha, [
1]. During this period, the average production yields (2.8–3.0 t∙ha
−1) were 33–38% lower than potential yields [
1]. Therefore, it is important to look for methods that can lead to higher rapeseed production and can increase its resilience to the pressure from changing environmental factors (drought, pressure of pathogens and pests, nutrient deficiency). Application of biostimulats, such as marine algae, is an alternative method that can support growth and hardiness a plant [
2,
3,
4]. Beneficial effects of algae extracts indicate their usefulness not only for horticulture crops but also for grown on large-scale commodity crops [
2,
3]. After, applications of biostimulants plants are more resistant to stress [
4,
5]. It has also been proved that biostimulants improve the uptake and accumulation of macro and microelements [
6,
7,
8,
9,
10,
11,
12,
13], which may have a favorable effect on yields of cultivated crops. For winter rapeseed, K and P are important nutrients. Nutritional requirements of rapeseed for these macroelements are much higher than for cereal crops, but only a small part of them is removed with seeds [
14,
15]. Despite the high soil abundance of P and K, plants may be malnourished [
16]. Potassium is taken up by rapeseed in the largest amounts [
17,
18]. Insufficient availability of this nutrient strongly limits the growth and yield of rapeseed [
19,
20]. According to Szczepaniak [
18], the number of seeds in pods, as a prerequisite of the final yield, depends on the K management pattern in rapeseed canopy before and after the flowering stage. In turn, P deficiency restricts both aboveground biomass and root growth [
20,
21]. Increase of available P in soil have led to a significant increase in rapeseed yield [
22]. Winter rapeseed demonstrates a high demand for S [
23], but soil enrichment from atmospheric S pollution has been reduced by 85% in the last three decades [
24]. Combined with an increased S removal by high yielding cultivars of rapeseed, and suboptimal S fertilization regimes, it appears that soil S deficiency has become a problem over the recent years [
25].
Many researchers indicate the mutual relationship of nutrients (N, P, K and others) in affecting their accumulation, aboveground and belowground biomass production and seed yield of rapeseed [
18,
19,
26,
27]. Deficiency of any nutrient limits the growth and crop yield [
14,
15,
16]. Increasing rates of one component, in turn, results in higher requirements for the others [
15]. According to Wang et al. [
21], a balanced application of nutrients should be considered with the aim to build an appropriate population structure with balanced plant density for optimum growth of every individual rapeseed plant.
The aim of this study was to assess P and K accumulation by the shoots and roots of winter rapeseed at successive stages of generative development: Flowering, fruit development and ripening, after the application of a seaweed biostimulant under varied NPK fertilization levels (high and low), as well as after and without fertilization with S.
3. Results
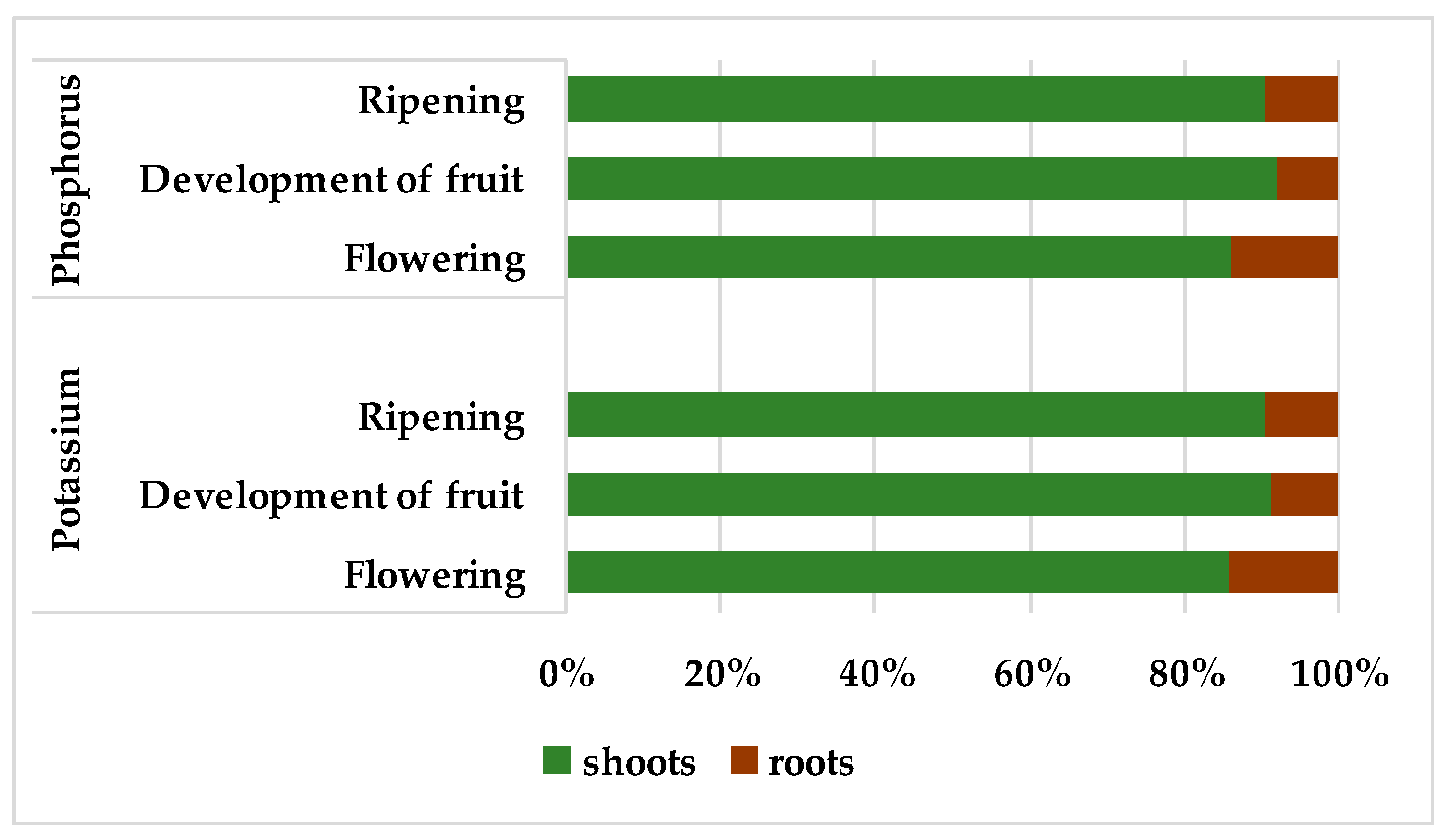
The percentage of shoots in the total P and K accumulation in the whole plant of winter rapeseed, accounted for approximately 89% for both macronutrients, on average across flowering, development of fruit and ripening stages (
Figure 1). The accumulation of P and K in roots, on average for these three generative stages, accounted for 11%, and it was the highest (14%) at flowering.
The application of the seaweed biostimulant, on average for the other experimental treatments, increased P accumulation in shoots at each developmental stage (
Table 3 and
Table 4). Moreover, at all growing stages, after the biostimulant application, P uptake in the rapeseed shoots at the lower NPK fertilization level was similar to that after the use of higher fertilization rates without the biostimulant. A similar effect of the seaweed extract was found in treatments with varying S fertilization levels.
Accumulation of P in rapeseed shoots increased with crop development progression, and the increase was the highest between the flowering stage and fruit development (
Table 4).
Favorable effect of the biostimulant on the averaged P accumulation, at the flowering and ripening stages, was also prominent in rapeseed roots (
Table 3 and
Table 5). Interaction of the biostimulant and S fertilization at flowering and ripening indicates that P uptake in the roots was higher after the biostimulant application only when rapeseed was fertilized with S.
On average, for the other factors, increased NPK fertilization level caused a significant increase in P accumulation in the shoots and roots at all developmental stages (
Table 3,
Table 4 and
Table 5). Rapeseed fertilized with S accumulated significantly more P in the shoots only at the flowering and fruit development stages. However, no significant effect of S fertilization on P accumulation in roots was found. At the fruit development stage, however, a significant interaction of NPK and S fertilization on P accumulation in shoots and roots was indicated. Only at the higher NPK fertilization level rapeseed fertilized with S accumulated significantly more P, compared with the same treatment that did not receive S.
Application of the biostimulant, the use of the higher NPK fertilization level or fertilization with S caused an increase in K accumulation in shoots at each analyzed stage of generative development (
Table 6 and
Table 7). A significant interaction of NPK and S fertilization at the fruit development stage indicates that only at the higher NPK rates, the application of S caused an increase in K accumulation in shoots. Application of biostimulat resulted that K accumulation in shoots of rapeseed, at each development stage, under either lower NPK fertilization level or without fertilization with S was similar to treatments receiving higher NPK rates or S without biostimulat application.
Accumulation of K in winter rapeseed shoots was 2.7 times higher at the fruit development stage than at the flowering stage (
Table 7). A further increase in accumulation of this nutrient until the ripening stage was slow—reaching only 14.5%.
In the case of rapeseed roots, the increase of K accumulation at the higher NPK fertilization level was recorded at each developmental stage and after the biostimulant application, but there were no reactions to S fertilization (
Table 6 and
Table 8).
On average in the period of generative growth, there was an increase in seed yield per 1 kg of the nutrient accumulated in the aboveground part (IE), amounted to 103 kg ha
−1 for P and 20 kg ha
−1 for K (
Table 9). IE was lower at the higher cultivation technology level (after the application of high NPK rates, as well as after the application of S or the biostimulant).
Uptake of both nutrients (P and K) per seed yield unit (RIE) increased at successive stages of generative development. RIE of P during the ripening stage was close to 2 and K was 3.4 times higher compared with the flowering stage. RIE of P and K, on average at the generative developmental stages, was similar at a higher and lower level of NPK fertilization. Sulphur fertilization, in turn, caused an increase in RIE as compared with the lack of fertilization with that element by 7.6% and 6.2%, respectively. Similarly, after the biostimulant application, RIE of P and K was higher than without the application by 8.8% and 7.0%, respectively.
4. Discussion
Winter rapeseed accumulates the highest amount of nutrients in shoots, and only their small portion in roots [
6]. In the present study, the proportion of aboveground part in the total P and K accumulation on average in the period of generative development accounted for about 90% (
Figure 1). This proportion for both elements was slightly lower at the flowering stage as compared with the fruit development and ripening stages. In the study by Billard et al. [
6], the proportion of P and K accumulation in the aboveground biomass of young (37 day) rapeseed plants were still lower and amounted to 74%.
P and K accumulation in shoots increased at the successive stages of generative development, and this increase was the highest between the flowering and fruit development stages (
Table 4 and
Table 7). Barłóg and Grzebisz [
36] also report a considerable increase in P and K accumulation during that period. According to these authors, increasing the accumulation of nutrients during the generative growth results from a very intensive increase in dry matter production. Szczepaniak [
18] reports that the pattern of K uptake by stems, since the onset of flowering significantly affected the structural yield components. This relationship can be explained by the requirements of growing seeds during plant ripening.
In our study, the application of the biostimulant increased P and K accumulation in the aboveground biomass of rapeseed at the flowering, fruit development and ripening stages (
Table 4 and
Table 7). Also, a favorable effect of the biostimulant on the accumulation of these elements in roots was recorded (P at the flowering and ripening stages, and K at each developmental stage) (
Table 5 and
Table 8). After the biostimulant application, P and K uptake in the aboveground part at the lower NPK fertilization level was similar to that after the use of higher rates without the preparation. Similar effect of the biostimulant was found in treatments with varied fertilization with S. Increase in P and K uptake after the application of biostimulants was also shown in the studies on different crops [
9]. The effect of these preparations results from the stimulation of many physiological processes responsible for the growth and development of above- and underground biomass [
37,
38,
39,
40]. In the case of rapeseed, it was stated that the extract from algae increased the accumulation of P, as well as of N and Ca [
6]. Those increases were explained by plant growth stimulation. The study by Jannin et al. [
7] proved an increase in the expression of genes responsible for nutrient uptake of rapeseed after the application of preparations from algae. Whereas, Billard et al. [
6] proved a decrease in the number of chloroplasts per leaf cell and root biomass production. According to Schnepf et al. [
41], young parts of the roots system are responsible for most of the P uptake. Increase in NPK fertilization rate causes, on average for the biostimulant application and fertilization with S, an increase in P and K accumulation in the aboveground part and in the roots at all the studied developmental stages (
Table 4,
Table 5,
Table 6,
Table 7 and
Table 8). Rapeseed fertilized with S accumulated significantly more K in the aboveground part at each stage of generative development and more P at the flowering and fruit development stages. Similar relationships are presented by Szczepaniak [
18]. In his study, K accumulation in rapeseed at flowering and ripening was higher if the plants were fertilized with NPK as compared with the control. Zou et al. [
35] proved an increase in K concentration in plants after the fertilization of rapeseed with this macronutrient. Wang et al. [
21] report that the accumulation of nutrients in the plant changes depending on their concentration in plant and dry matter production, whereas the dry matter production is fundamentally important. Reduction in the rates of elements limits biomass production [
14,
19,
21], as well as the element concentration [
16,
21], and thus, the degree of their accumulation [
21]. According to Brennan and Bolland [
26], the response of rapeseed to P and K fertilization also depends on the content of these elements in the soil. Similarly, Gaj [
17] shows the essential role of soil resources of available K forms in the state of plant nutrition with this element.
At the fruit development stage, there was a significant interaction between NPK and S fertilization in P and K accumulation in the aboveground parts, as well as in P accumulation in the roots (
Table 3 and
Table 6). Only at the high NPK fertilization level rapeseed fertilized with S accumulated significantly more P and K as compared with that not fertilized. The literature data shows that the accumulation of the given nutrients by rapeseed depends on the availability of the other ones. Wang et al. [
21] claim that P uptake is reduced by N and K deficiency, and K uptake is lower both at the lesser availability of K, and also the deficiency of N and P. According to Gaj [
16], K deficiency limits N accumulation and transport, which leads to a reduction in growth and assimilation area. Szczepaniak [
18] proved that N accumulation at the beginning of the flowering of rapeseed depended on K accumulation in the aboveground part. Application of some nutrients (e.g., N) has a strong effect on dry matter increase, which results in increased demand for the other elements. Therefore, balanced fertilization is essential for increasing the yield. Giovahi and Saffari [
19] indicated K and S interaction in dry matter production. Brenan and Bolland [
15,
27] proved that for obtaining high yields of rapeseed at a growing level of fertilization with N, it is necessary to increase P and K rates.
Increase in seed yield per 1 kg of P and K accumulated in the aboveground part (IE) was reduced, and the uptake of those elements per 1000 kg ha
−1 of seed yield (RIE) was greater at successive stages of generative development (
Table 9). IE was smaller at the higher level of cultivation practices (after the application of higher NPK rates, as well as in the case of fertilization with S or application of the biostimulant). Similarly, Zou et al. [
35] indicated a decrease in this indicator value at the increased fertilization value. When plants were well fertilized, with an additional uptake stimulation coming from biostimulant application, the nutrients accumulation in the plant increased; but, the increase in yield was not proportional. According to Ren et al. [
34], P and K uptake by rapeseed increases linearly until the yield reaches 0.6–0.7 of the potential yield. However, when the target yield approaches the yield potential, a decrease in IE is detected.





{kind=link}