Effects of Sub-Optimal Temperatures on Seed Germination of Three Warm-Season Turfgrasses with Perspectives of Cultivation in Transition Zone


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Germination Assay: Experimental Setting
2.2. Soil Temperature Data
2.3. Data Analyses
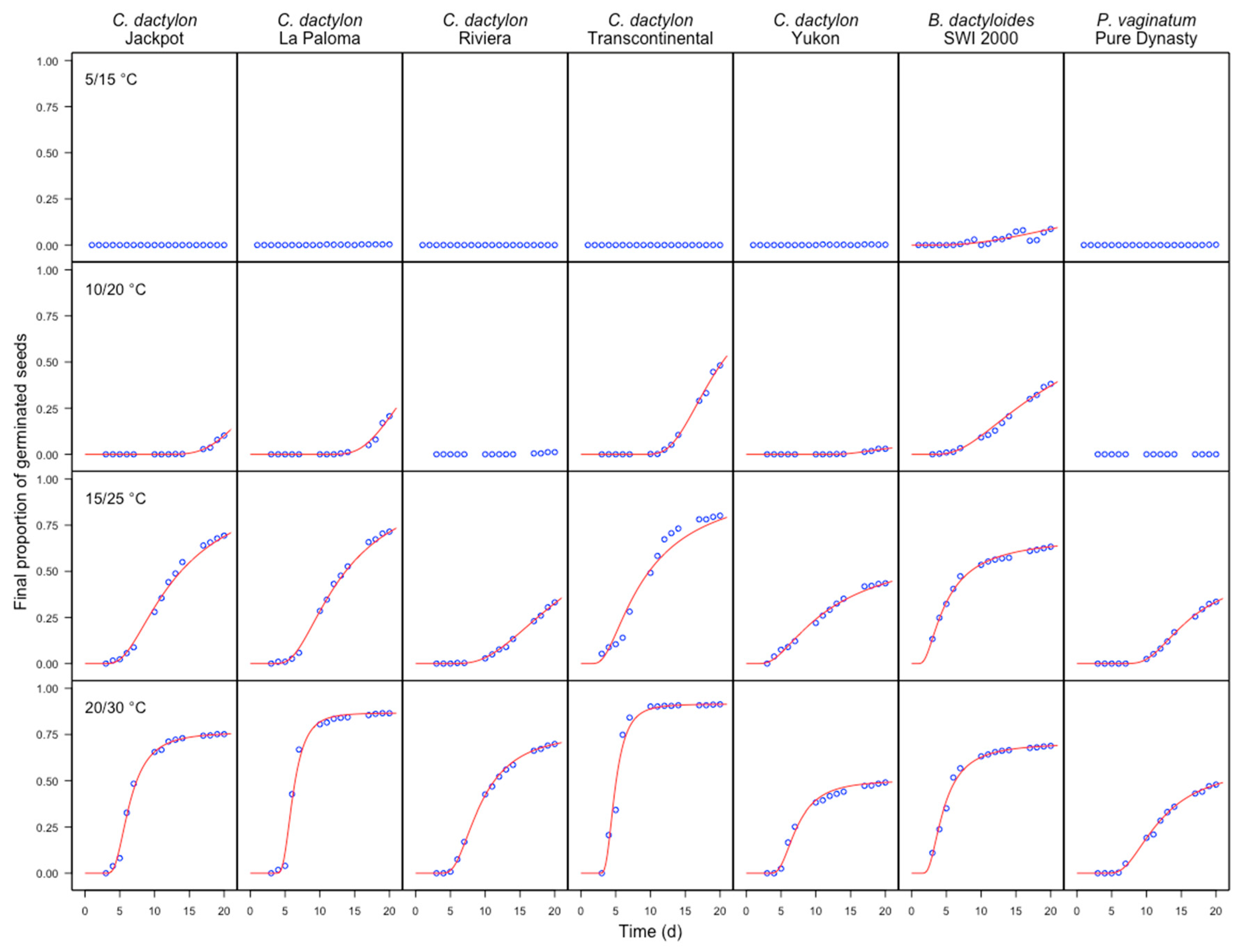
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Green, R.L.; Beard, J.B.; Casnoff, D.M. Leaf Blade Stomatal Characterizations and Evapotranspiration Rates of 12 Cool-season Perennial Grasses. HortScience 1990, 25, 760–761. [Google Scholar]
- Huang, B. Turfgrass Water Requirements and Factors Affecting Water Usage; Beard, J.B., Kenna, M.P., Eds.; CAST Special Publication; Water Quality and Quantity Issues for Turfgrass in Urban Landscapes; PSI Printing Services, Inc.: Belmond, IWA, USA.
- Minelli, A.; De Luca, A.; Croce, P.; Cevenini, L.; Zuffa, D. Transition from cool-season to warm-season grass: Environmental effects in a golf course in the North of Italy. In Proceedings of the 4th European Turfgrass Society Conference, Osnabrueck, Germany, 6–9 July 2014; pp. 1–4. [Google Scholar]
- Puhalla, J.; Krans, J.; Goatley, M. Sports Fields: A Manual for Design, Construction and Maintenance; John Wiley & Sons, Inc.: Hoboken, NJ, USA; ISBN 978-1-57504-070-7.
- Sever Mutlu, S.; Mutlu, N.; Shearman, R.; Gurbuz, E.; Gulsen, O.; Hocagil, M.M.; Karaguzel, O.; Heng-Moss, T.; Riordan, T.; Gaussoin, R. Establishment and Turf Qualities of Warm-season Turfgrasses in the Mediterranean Region. HortTechnology 2011, 21, 67–81. [Google Scholar] [CrossRef]
- Rimi, F.; Macolino, S.; Richardson, M.D.; Karcher, D.E.; Leinauer, B. Influence of Three Nitrogen Fertilization Schedules on Bermudagrass and Seashore Paspalum: I. Spring Green-up and Fall Color Retention. Crop Sci. 2013, 53, 1161–1167. [Google Scholar] [CrossRef]
- Rimi, F.; Macolino, S.; Richardson, M.D.; Karcher, D.E.; Leinauer, B. Influence of Three Nitrogen Fertilization Schedules on Bermudagrass and Seashore Paspalum: II. Carbohydrates and Crude Protein in Stolons. Crop Sci. 2013, 53, 1168–1178. [Google Scholar] [CrossRef]
- Magni, S.; Gaetani, M.E.; Grossi, N.; Caturegli, L.; Bella, S.L.; Leto, C.; Virga, G.; Tuttolomondo, T.; Lulli, F. Bermudagrass adaptation in the Mediterranean climate: Phenotypic traits of 44 accessions. Adv. Hortic. Sci. 2014, 28, 29–34. [Google Scholar]
- Seppoloni, I.; Staglianò, N.; Cecchi, S.; Argenti, G. Performance of warm-season turfgrasses in an area of central Italy. Adv. Hortic. Sci. 2015, 29, 53–58. [Google Scholar]
- Schiavon, M.; Macolino, S.; Leinauer, B.; Ziliotto, U. Seasonal Changes in Carbohydrate and Protein Content of Seeded Bermudagrasses and Their Effect on Spring Green-Up. J. Agron. Crop Sci. 2016, 202, 151–160. [Google Scholar] [CrossRef]
- Munshaw, G.C.; Williams, D.W.; Powell, A.J.; Dougherty, C.T. Growth and development of seeded versus vegetative bermudagrass varieties. Agron. Abstr. 1998, 136. [Google Scholar]
- Pessarakli, A.J. Handbook of Turfgrass Management and Physiology; CRC Press, Taylor & Francis Publishing Company: Boca Raton, FL, USA, 2007; ISBN 978-1-4200-0648-3. [Google Scholar]
- Patton, A.J.; Hardebeck, G.A.; Williams, D.W.; Reicher, Z.J. Establishment of Bermudagrass and Zoysiagrass by Seed. Crop Sci. 2004, 44, 2160–2167. [Google Scholar] [CrossRef]
- Hanna, W.W.; Anderson, W.F. Development and Impact of Vegetative Propagation in Forage and Turf Bermudagrasses. Agron. J. 2008, 100, S-103–S-107. [Google Scholar] [CrossRef]
- Patton, A.J.; Richardson, M.D.; Karcher, D.E.; Boyd, J.W.; Reicher, Z.J.; Fry, J.D.; McElroy, J.S.; Munshaw, G.C. A Guide to Establishing Seeded Bermudagrass in the Transition Zone. Ats 2008, 5, 19. [Google Scholar] [CrossRef]
- Pornaro, C.; Macolino, S.; Leinauer, B. Seeding time affects establishment of warm-season turfgrasses. Acta Hortic. 2016, 1122, 27–34. [Google Scholar] [CrossRef]
- Rimi, F.; Macolino, S.; Leinauer, B.; Ziliotto, U. Green-up of Seeded Bermudagrass Cultivars as Influenced by Spring Scalping. HortTechnology 2011, 21, 230–235. [Google Scholar] [CrossRef]
- HNMS Hellenic National Meteorological Service. Available online: http://www.emy.gr/emy/en/ (accessed on 6 May 2019).
- Musser, H.B.; Perkins, A.T. Guide to Seedbed Preparation. In Hanson, A.A., and Juska, F.V. Turfgrass Science; American Society of Agronomy: Madison, WI, USA, 1969; pp. 462–473. [Google Scholar]
- Richardson, M.D.; Karcher, D.E.; Berger, P.; Boyd, J.W. Utilizing improved seeded bermudagrasses on transition-zone sports fields. Acta Hortic. 2004, 661, 369–374. [Google Scholar] [CrossRef]
- Shaver, B.R.; Richardson, M.D.; McCalla, J.H.; Karcher, D.E.; Berger, P.J. Dormant Seeding Bermudagrass Cultivars in a Transition-Zone Environment. Crop Sci. 2006, 46, 1787–1792. [Google Scholar] [CrossRef]
- Deaton, M.T.; Williams, D.W. Temperature Effects on the Speed and Completion of Germination of 19 Commercially Available Seeded Bermudagrass Cultivars. HortTechnology 2013, 23, 82–85. [Google Scholar] [CrossRef]
- Giolo, M.; Ferrari, F.; Macolino, S. Estimation of Base Germination Temperature of Ten Seeded-Type Bermudagrass Cultivars. Eur. J. Hortic. Sci. 2014, 79, 129–134. [Google Scholar]
- Fontenot, D.P. Evaluating Seashore Paspalum Seed Germination and Enhancement, Erosion Abatement and Potential Use as a Vegetative Landfarm Cap. Master’s Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2007. [Google Scholar]
- Unruh, J.B.; Gaussoin, R.E.; Wiest, S.C. Basal Growth Temperatures and Growth Rate Constants of Warm-Season Turfgrass Species. Crop Sci. 1996, 36, 997–999. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds:Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4693-4. [Google Scholar]
- Garcia-Huidobro, J.; Monteith, J.L.; Squire, G.R. Time, Temperature and Germination of Pearl Millet (Pennisetum typhoides S. & H.): I. constant temperature. J. Exp. Bot. 1982, 33, 288–296. [Google Scholar]
- Sandlin, T.N.; Munshaw, G.; Philley, H.W.; Baldwin, B.S.; Steward, B.S. Temperature Affects Germination of Seeded Bermudagrasses. In Proceedings of the ASA-CSSA-SSSA International Annual Meeting, Indianapolis, IN, USA, 12–16 November 2006. [Google Scholar]
- Wu, L.; Huff, D.; Harivandi, M. Buffalograss as a low-maintenance turf. Calif. Agric. 1989, 43, 23–25. [Google Scholar]
- Johnson, P.G.; Riordan, T.P.; Johnson-Cicalese, J. Low-Mowing Tolerance in Buffalograss. Crop Sci. 2000, 40, 1339–1343. [Google Scholar] [CrossRef]
- ISTA (International Seed Testing Association). International Rules for Seed Testing, 2013 ed.; ISTA: Zurich, Switzerland, 2013; ISBN 13 978-3-906549-72-9. [Google Scholar]
- Svoboda, J.F. Seedling Germination and Establishment of Buffalograss Caryopses vs. Burs. Master’s Thesis, Faculty of The Graduate College of the University of Nebraska, Lincoln, NE, USA, 1991. [Google Scholar]
- Regional Agency for Environmental Protection of Veneto Region (ARPAV). Dipartimento per la Sicurezza del Territorio. Centro Meterologico: Teolo, Padova, Italy, 2019. Available online: http://www.arpa.veneto.it/temi-ambientali/climatologia/dati (accessed on 11 April 2019).
- Weisberg, S. Applied Linear Regression|Wiley Series in Probability and Statistics, 3rd ed.; John Wiley & Sons Inc. Books: Hoboken, NJ, USA, 2005. [Google Scholar]
- Ritz, C.; Pipper, C.B.; Streibig, J.C. Analysis of germination data from agricultural experiments. Eur. J. Agron. 2013, 45, 1–6. [Google Scholar] [CrossRef]
- Onofri, A.; Benincasa, P.; Mesgaran, M.B.; Ritz, C. Hydrothermal-time-to-event models for seed germination. Eur. J. Agron. 2018, 101, 129–139. [Google Scholar] [CrossRef]
- Masin, R.; Onofri, A.; Gasparini, V.; Zanin, G. Can alternating temperatures be used to estimate base temperature for seed germination? Weed Res. 2017, 57, 390–398. [Google Scholar] [CrossRef]
- Box, G.E.P.; Cox, D.R. An Analysis of Transformations. J. R. Stat. Soc. Ser. B (Methodol.) 1964, 26, 211–252. [Google Scholar] [CrossRef]
- McMaster, G. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Moore, J.L.; Remais, J.V. Developmental models for estimating ecological responses to environmental variability: Structural, parametric, and experimental issues. Acta Biotheor. 2014, 62, 69–90. [Google Scholar] [CrossRef]
- R Core Team. R 3.5.0 R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Ritz, C.; Strebig, J.C. Analysis of Dose-Response Curves, “drc”, R package version 3.0-1; 2016. Available online: https://rdrr.io/cran/drc/ (accessed on 15 May 2019).
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, aka Least-Squares Means. 2018. Available online: https://rdrr.io/cran/emmeans/man/emmeans.html (accessed on 30 May 2019).
- Bradford, K.J. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- The Council of the European Economic Community, Council Directive of 14 June 1966 on the Marketing of Fodder Plant Seed. Conditions to be Satisfied by the Seed; EU Publications: Luxembourg, Luxembourg, 1966. [Google Scholar]
- Shin, J.S.; Raymer, P.; Kim, W. Environmental factors influencing germination in seeded seashore paspalum. Hortscience 2006, 41, 1330–1331. [Google Scholar] [CrossRef]
- Budak, H.; Shearman, R.C.; Parmaksiz, I.; Dweikat, I. Comparative analysis of seeded and vegetative biotype buffalograsses based on phylogenetic relationship using ISSRs, SSRs, RAPDs, and SRAPs. Theor. Appl. Genet. 2004, 109, 280–288. [Google Scholar] [CrossRef]
- Larsen, S.; Martin Bibby, B. Differences in Thermal Time Requirement for Germination of Three Turfgrass Species. Crop Sci. 2005, 45, 2030–2037. [Google Scholar] [CrossRef]
- OECD. The OECD List of Varieties Eligible for Seed Certification, 2019th Edition. Available online: http://www.oecd.org/agriculture/seeds/documents/codes-and-schemes-list-of-varieties-eligible-for-seed-certification.pdf (accessed on 5 May 2019).



{kind=link}
{kind=link}
| Temp. | Month | 10-Day Interval | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | Average | St. Dev. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| max | March | 1 | 10.0 | 10.0 | 6.9 | 9.5 | 12.8 | 11.7 | 12.4 | 10.5 | 2.03 |
| mean | 7.0 | 7.9 | 4.5 | 4.4 | 7.5 | 7.7 | 8.9 | 6.8 | 1.73 | ||
| min | 4.7 | 6.0 | 2.9 | 0.6 | 3.7 | 4.8 | 6.4 | 4.2 | 1.98 | ||
| max | 2 | 12.2 | 11.8 | 9.4 | 13.2 | 16.7 | 10.7 | 16.1 | 12.9 | 2.69 | |
| mean | 8.9 | 8.3 | 5.6 | 10.1 | 10.0 | 7.5 | 10.4 | 8.7 | 1.72 | ||
| min | 6.2 | 5.3 | 2.7 | 7.5 | 5.1 | 5.0 | 6.3 | 5.4 | 1.49 | ||
| max | 3 | 11.1 | 11.2 | 14.0 | 15.6 | 18.3 | 9.4 | 17.4 | 13.9 | 3.41 | |
| mean | 7.7 | 8.6 | 11.3 | 10.4 | 12.2 | 6.8 | 11.6 | 9.8 | 2.10 | ||
| min | 4.8 | 6.4 | 9.2 | 6.0 | 7.0 | 4.6 | 6.8 | 6.4 | 1.54 | ||
| max | April | 1 | 13.7 | 20.8 | 15.4 | 20.6 | 15.6 | 12.0 | 20.1 | 16.9 | 3.59 |
| mean | 10.1 | 14.4 | 11.2 | 14.5 | 12.3 | 9.4 | 14.6 | 12.4 | 2.20 | ||
| min | 7.0 | 10.4 | 7.9 | 9.6 | 9.3 | 7.4 | 10.1 | 8.8 | 1.36 | ||
| max | 2 | 15.2 | 23.7 | 16.9 | 21.0 | 14.3 | 18.4 | 20.3 | 18.5 | 3.36 | |
| mean | 12.1 | 14.9 | 12.7 | 14.0 | 11.9 | 14.5 | 14.5 | 13.5 | 1.25 | ||
| min | 9.7 | 9.5 | 9.3 | 8.5 | 9.9 | 11.2 | 9.6 | 9.7 | 0.81 | ||
| max | 3 | 16.4 | 20.5 | 20.4 | 22.9 | 16.8 | 18.7 | 21.5 | 19.6 | 2.41 | |
| mean | 13.5 | 13.7 | 16.1 | 16.5 | 13.4 | 15.3 | 16.8 | 15.0 | 1.49 | ||
| min | 11.3 | 9.5 | 12.7 | 11.7 | 10.7 | 12.1 | 13.3 | 11.6 | 1.27 | ||
| max | May | 1 | 18.2 | 25.4 | 20.4 | 26.0 | 24.4 | 23.1 | 22.9 | 22.9 | 2.78 |
| mean | 14.8 | 17.5 | 15.5 | 17.8 | 18.9 | 19.1 | 17.8 | 17.3 | 1.62 | ||
| min | 11.9 | 12.2 | 11.3 | 11.2 | 14.1 | 16.0 | 13.8 | 12.9 | 1.77 | ||
| max | 2 | 19.0 | 27.3 | 22.0 | 27.5 | 22.6 | 20.8 | 23.4 | 23.2 | 3.18 | |
| mean | 16.0 | 20.5 | 16.8 | 20.2 | 17.4 | 17.4 | 18.0 | 18.0 | 1.69 | ||
| min | 13.2 | 15.8 | 13.1 | 14.0 | 12.6 | 14.4 | 13.7 | 13.8 | 1.06 | ||
| max | 3 | 24.2 | 29.1 | 26.8 | 31.0 | 24.4 | 20.3 | 25.8 | 25.9 | 3.50 | |
| mean | 19.5 | 22.0 | 19.7 | 23.1 | 20.2 | 16.5 | 20.7 | 20.2 | 2.09 | ||
| min | 15.8 | 16.7 | 14.2 | 16.6 | 16.8 | 13.3 | 16.7 | 15.7 | 1.42 |
| Species | Cultivar | Temperature Regime | |||
|---|---|---|---|---|---|
| 5/15 °C | 10/20 °C | 15/25 °C | 20/30 °C | ||
| FGP (%) | |||||
| bermudagrass | Jackpot | 0.0 (0.0) | 10.2 (1.8) | 69.5 (3.5) | 75.5 (3.1) |
| bermudagrass | La Paloma | 0.3 (0.3) | 20.5 (2.8) | 71.7 (3.4) | 86.8 (2.2) |
| bermudagrass | Riviera | 0.0 (0.0) | 1.1 (0.5) | 33.0 (3.6) | 70.1 (3.5) |
| bermudagrass | Transcontinental | 0.0 (0.0) | 48.2 (4.0) | 80.4 (2.8) | 91.6 (1.7) |
| bermudagrass | Yukon | 0.2 (0.2) | 2.9 (0.9) | 43.4 (4.0) | 49.0 (4.0) |
| buffalograss | SWI 2000 | 8.7 (1.6) | 38.1 (3.8) | 63.5 (3.8) | 69.1 (3.5) |
| seashore paspalum | Pure Dynasty | 0.2 (0.2) | 0.0 (0.0) | 33.3 (3.7) | 47.8 (4.0) |
| Species | Cultivar | θT(10) | θT(20) | θT(30) | Tb (°C) |
|---|---|---|---|---|---|
| (°C d) | |||||
| bermudagrass | Jackpot | 47.3 (3.0) | 57.7 (4.1) | 65.1 (4.8) | 14.1 (0.17) |
| bermudagrass | La Paloma | 53.9 (3.8) | 63.5 (4.8) | 72.2 (5.7) | 13.1 (0.28) |
| bermudagrass | Riviera | 47.3 (5.6) | 62.7 (8.0) | 75.9 (10.2) | 17.0 (0.52) |
| bermudagrass | Transcontinental | 45.2 (3.1) | 54.3 (3.9) | 61.2 (4.6) | 11.5 (0.43) |
| bermudagrass | Yukon | 45.9 (2.9) | 59.3 (4.2) | 76.8 (6.2) | 15.0 (0.03) |
| buffalograss | SWI 2000 | 30.5 (1.8) | 41.3 (2.6) | 51.3 (3.5) | 11.4 (0.39) |
| seashore papsalum | Pure Dynasty | 78.4 (6.4) | 105.8 (10.0) | 144.0 (15.9) | 15.0 (0.03) |
| Species | Cultivar | Month and Decade | |||||||
|---|---|---|---|---|---|---|---|---|---|
| March | April | May | |||||||
| 2° | 3° | 1° | 2° | 3° | 1° | 2° | 3° | ||
| Cumulated degree days | |||||||||
| buffalograss | SWI 2000 | 7.5 | 20 | 47.5 | 83 | 125 | 190 | 261 | 355 |
| bermudagrass | Transcontin. | 7 | 19 | 46 | 81 | 122 | 186 | 256 | 349 |
| bermudagrass | La Paloma | 19 | 46 | 78.5 | 127.5 | 181.5 | 258.5 | ||
| bermudagrass | Jackpot | 14 | 36 | 63.5 | 107.5 | 153 | 220 | ||
| bermudagrass | Yukon | 9.5 | 27 | 50 | 89.5 | 130.5 | 188.5 | ||
| seashore paspalum | Pure Dynasty | 9.5 | 27 | 50 | 89.5 | 130.5 | 188.5 | ||
| bermudagrass | Riviera | 7.5 | 20.5 | 50 | 81 | 125.5 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giolo, M.; Benincasa, P.; Anastasi, G.; Macolino, S.; Onofri, A. Effects of Sub-Optimal Temperatures on Seed Germination of Three Warm-Season Turfgrasses with Perspectives of Cultivation in Transition Zone. Agronomy 2019, 9, 421. https://doi.org/10.3390/agronomy9080421
Giolo M, Benincasa P, Anastasi G, Macolino S, Onofri A. Effects of Sub-Optimal Temperatures on Seed Germination of Three Warm-Season Turfgrasses with Perspectives of Cultivation in Transition Zone. Agronomy. 2019; 9(8):421. https://doi.org/10.3390/agronomy9080421
Chicago/Turabian StyleGiolo, Maurizio, Paolo Benincasa, Giuseppe Anastasi, Stefano Macolino, and Andrea Onofri. 2019. "Effects of Sub-Optimal Temperatures on Seed Germination of Three Warm-Season Turfgrasses with Perspectives of Cultivation in Transition Zone" Agronomy 9, no. 8: 421. https://doi.org/10.3390/agronomy9080421
APA StyleGiolo, M., Benincasa, P., Anastasi, G., Macolino, S., & Onofri, A. (2019). Effects of Sub-Optimal Temperatures on Seed Germination of Three Warm-Season Turfgrasses with Perspectives of Cultivation in Transition Zone. Agronomy, 9(8), 421. https://doi.org/10.3390/agronomy9080421