Genetic Diversity of Sweet Potato (Ipomoea batatas L. Lam) Germplasms Collected Worldwide Using Chloroplast SSR Markers

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. DNA Extraction
2.3. Chloroplast SSR Genotyping
2.4. Data Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ramanatha Rao, V.; Hodgkin, T. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Organ Cult. 2002, 68, 1–19. [Google Scholar] [CrossRef]
- Cadima Fuentes, X.; van Treuren, R.; Hoekstra, R.; van den Berg, R.G.; Sosef, M.S.M. Genetic diversity of Bolivian wild potato germplasm: Changes during ex situ conservation management and comparisons with resampled in situ populations. Genet. Resour. Crop Evol. 2017, 64, 331–344. [Google Scholar] [CrossRef]
- Xiao, L.Q.; Ge, X.J.; Gong, X.; Hao, G.; Zheng, S.X. ISSR variation in the endemic and endangered plant Cycas guizhouensis (Cycadaceae). Ann. Bot. 2004, 94, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Katayama, K.; Kobayashi, A.; Sakai, T.; Kuranouchi, T.; Kai, Y. Recent progress in sweetpotato breeding and cultivars for diverse applications in Japan. Breed. Sci. 2017, 67, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Cervantes, J.; Huamán, Z.; Carey, E.; Ghislain, M. Assessing genetic diversity of sweet potato (Ipomoea batatas (L.) Lam.) cultivars from tropical America using AFLP. Genet. Resour. Crop Evol. 2000, 47, 659–665. [Google Scholar] [CrossRef]
- Elameen, A.; Fjellheim, S.; Larsen, A.; Rognli, O.A.; Sundheim, L.; Msolla, S.; Masumba, E.; Mtunda, K.; Klemsdal, S.S. Analysis of genetic diversity in a sweet potato (Ipomoea batatas L.) germplasm collection from Tanzania as revealed by AFLP. Genet. Resour. Crop Evol. 2008, 55, 397–408. [Google Scholar] [CrossRef]
- Woolfe, J.A. Sweet Potato: An Untapped Food Resource; Cambridge University Press: Cambridge, UK; International Potato Center: Lima, Peru, 1992. [Google Scholar]
- Yang, X.S.; Su, W.J.; Wang, L.J.; Lei, J.; Chai, S.S.; Liu, Q.C. Molecular diversity and genetic structure of 380 sweetpotato accessions as revealed by SSR markers. J. Integr. Agric. 2015, 14, 633–641. [Google Scholar] [CrossRef]
- Zhang, D.; Rossel, G.; Kriegner, A.; Hijmans, R. AFLP assessment of diversity in sweetpotato from Latin America and the Pacific region: Its implications on the dispersal of the crop. Genet. Resour. Crop Evol. 2004, 51, 115–120. [Google Scholar] [CrossRef]
- Gichuki, S.T.; Berenyi, M.; Zhang, D.; Hermann, M.; Schmidt, J.; Glössl, J.; Burg, K. Genetic diversity in sweetpotato [Ipomoea batatas (L.) Lam.] in relationship to geographic sources as assessed with RAPD markers. Genet. Resour. Crop Evol. 2003, 50, 429–437. [Google Scholar] [CrossRef]
- Meng, Y.S.; Zhao, N.; Li, H.; Zhai, H.; He, S.Z.; Liu, Q.C. SSR fingerprinting of 203 sweetpotato (Ipomoea batatas (L.) Lam.) varieties. J. Integr. Agric. 2018, 17, 86–93. [Google Scholar] [CrossRef]
- Roullier, C.; Kambouo, R.; Paofa, J.; McKey, D.; Lebot, V. On the origin of sweet potato (Ipomoea batatas (L.) Lam.) genetic diversity in New Guinea, a secondary centre of diversity. Heredity 2013, 110, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Roullier, C.; Rossel, G.; Tay, D.; McKey, D.; Lebot, V. Combining chloroplast and nuclear microsatellites to investigate origin and dispersal of New World sweet potato landraces. Mol. Ecol. 2011, 20, 3963–3977. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J.; Bell, C.D.; Soltis, P.S.; Soltis, D.E. Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms. Proc. Natl. Acad. Sci. USA 2007, 104, 19363–19368. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, T.; Zhang, C.; Faheem, M.; Mu, Q.; Fang, J. Chloroplast based genetic diversity among Chinese grapes genotypes. Mitochondrial DNA Part A 2017, 28, 565–569. [Google Scholar] [CrossRef]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef]
- McCauley, D.E. The use of chloroplast DNA polymorphism in studies of gene flow in plants. Trends Ecol. Evol. 1995, 10, 198–202. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.; McNicol, J.W.; Machray, G.C.; Doyle, J.J.; Tingey, S.V.; Rafalski, J.A. Hypervariable microsatellites provide a general source of polymorphic DNA markers for the chloroplast genome. Curr. Biol. 1995, 5, 1023–1029. [Google Scholar] [CrossRef]
- Huang, J.; Yang, X.; Zhang, C.; Yin, X.; Liu, S.; Li, X. Development of Chloroplast Microsatellite Markers and Analysis of Chloroplast Diversity in Chinese Jujube (Ziziphus jujuba Mill.) and Wild Jujube (Ziziphus acidojujuba Mill.). PLoS ONE 2015, 10, e0134519. [Google Scholar] [CrossRef]
- Kim, I.H. East Asian Nations’ Awareness on Ocean during the 16-18th Century Perceived from Transfer of Sweet Potatoes. J. Asia Pac. Stud. 2014, 21, 69–97. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. Available online: http://darwin.cirad.fr/darwin (accessed on 10 October 2019).
- Kamvar, Z.N.; Tabima, J.F.; Grunwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Laurie, S.M.; Calitz, F.J.; Adebola, P.O.; Lezar, A. Characterization and evaluation of South African sweet potato (Ipomoea batatas (L.) LAM) land races. S. Afr. J. Bot. 2013, 85, 10–16. [Google Scholar] [CrossRef]
- Desiderio, F.; Bitocchi, E.; Bellucci, E.; Rau, D.; Rodriguez, M.; Attene, G.; Papa, R.; Nanni, L. Chloroplast Microsatellite Diversity in Phaseolus vulgaris. Front. Plant Sci. 2012, 3, 312. [Google Scholar] [CrossRef]
- Singh, N.; Pal, A.K.; Roy, R.K.; Tamta, S.; Rana, T.S. Development of cpSSR markers for analysis of genetic diversity in Gladiolus cultivars. Plant Gene 2017, 10, 31–36. [Google Scholar] [CrossRef]
- The International Potato Center (CIP). Global Strategy for Ex-Situ Conservation of sweetpotato Genetic Resources. Sweetpotato Genetic Resources Conservation—CIP November 2007. Available online: http://www.croptrust.org/documents/web/SweetPotato-Strategy-FINAL-12Dec07.pdf (accessed on 10 October 2019).
- Kawa, N. Tropical Root and Tuber Crops: Cassava, Sweet Potato, Yams and Aroids by V. Lebot. Econ. Bot. 2010, 64, 86–87. [Google Scholar]
- Shiga, T.; Sakamoto, S.; Ando, T.; Ishikawa, H.; Kato, S.; Takemata, T.; Umehara, M. On a new sweet potato [Ipomoea batatas] cultivar “Beniazuma”. Bull. Nat. Agric. Res. Cent. 1985, 3, 73–84. [Google Scholar]
- Katayama, K.; Tamiya, S.; Kuranouchi, T.; Komaki, K.; Nakatani, M. New sweet potato cultivar “Quick Sweet”. Bull. Natl. Inst. Crop Sci. 2003, 3, 35–52. [Google Scholar]
- Ohara-Takada, A.; Kumagai, T.; Kuranouchi, T.; Nakamura, Y.; Fujita, T.; Nakatani, M.; Tamiya, S.; Katayama, K. ‘Aikomachi’, a new sweetpotato cultivar with good appearance and high confectionery quality. Bull. Naro Inst. Crop Sci. 2016, 16, 35–55. [Google Scholar]
- RDA. Plant Germplasm Research 20; RDA: Jeonju, Korea, 2007. [Google Scholar]
- Lin, K.H.; Lai, Y.C.; Chang, K.Y.; Chen, Y.F.; Hwang, S.Y.; Lo, H.F. Improving breeding efficiency for quality and yield of sweet potato. Bot. Stud. 2007, 48, 283–292. [Google Scholar]
- Mok, I.G.; Zhao, D.; Kwak, S.S. Genetic resources of sweetpotato for industrial use. J. Plant Biotechnol. 2009, 36, 202. [Google Scholar] [CrossRef]
- Kobayashi, M.; Miyazaki, T. Sweet potato breeding using wild related species. In Proceedings of the Fourth Symposium of the International Society for Tropical Root Crops, IDRC, Cali, Colombia, 1–7 August 1976; Volume 1, pp. 53–57. [Google Scholar]
- Khoury, C.K.; Heider, B.; Castañeda-Álvarez, N.P.; Achicanoy, H.A.; Sosa, C.C.; Miller, R.E.; Scotland, R.W.; Wood, J.R.I.; Rossel, G.; Eserman, L.A.; et al. Distributions, ex situ conservation priorities, and genetic resource potential of crop wild relatives of sweetpotato [Ipomoea batatas (L.) Lam., I. series Batatas]. Front. Plant Sci. 2015, 6, 251. [Google Scholar] [CrossRef] [PubMed]
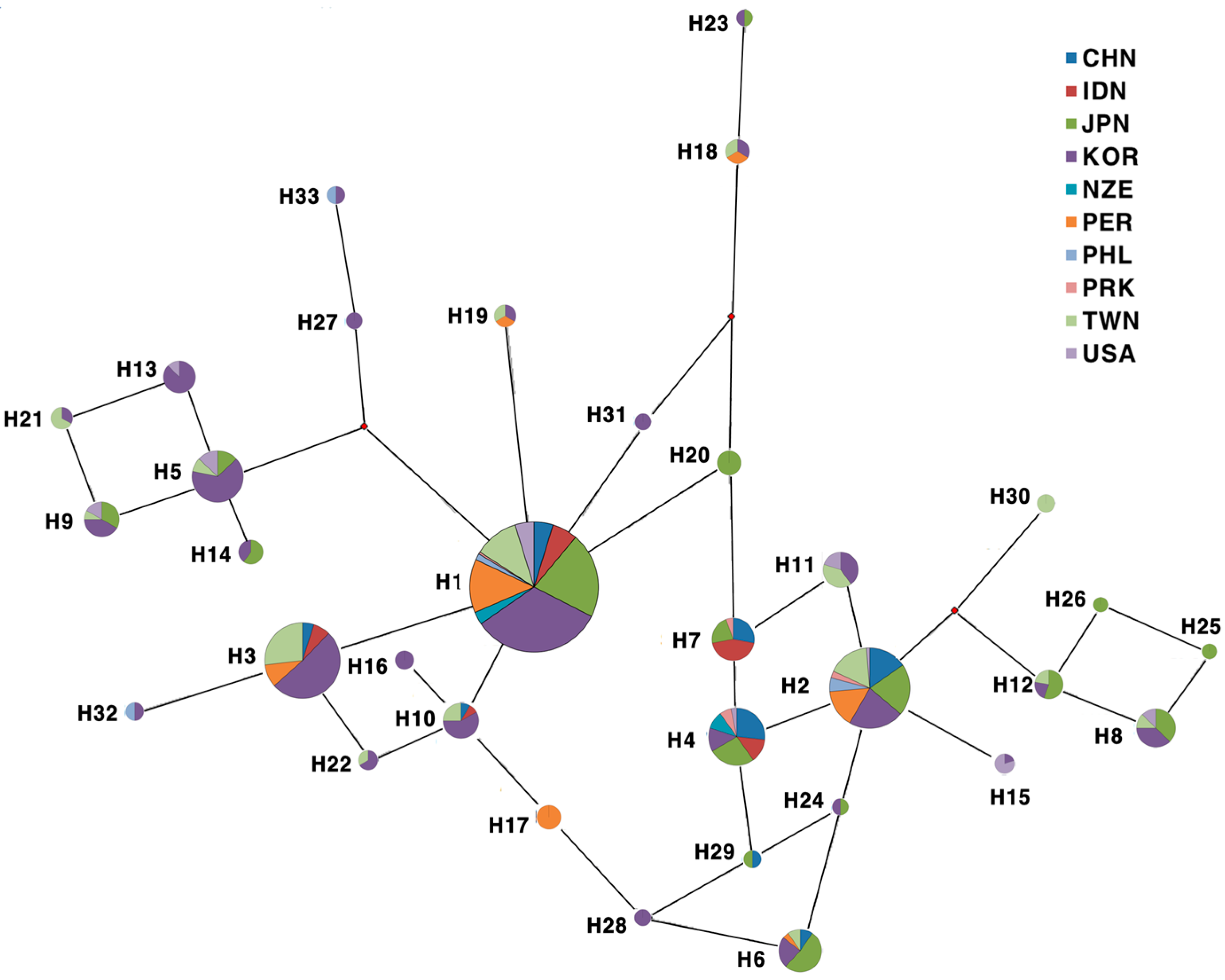
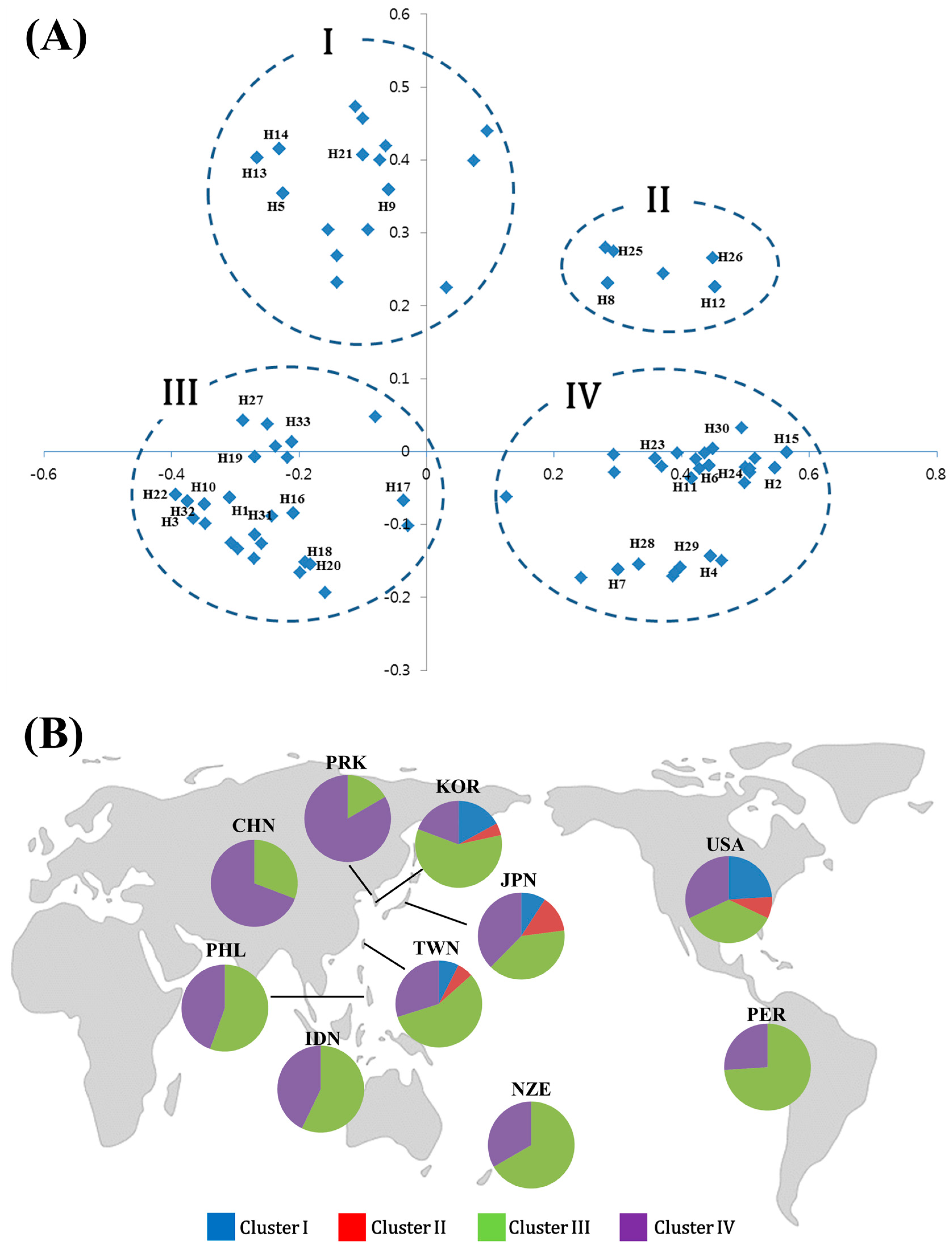

{kind=link}
{kind=link}
| Unknown | Breeding Line | Cultivar | Landrace | Total | |
|---|---|---|---|---|---|
| CHN 1 | 11 | 2 | 29 | 1 | 43 |
| IDN | 3 | 18 | 9 | 30 | |
| JPN | 15 | 22 | 79 | 7 | 123 |
| KOR | 125 | 48 | 17 | 190 | |
| NZE | 9 | 9 | |||
| PER | 50 | 50 | |||
| PHL | 1 | 8 | 9 | ||
| PRK | 2 | 3 | 1 | 6 | |
| TWN | 31 | 32 | 10 | 73 | |
| USA | 22 | 3 | 25 | ||
| Total | 60 | 184 | 276 | 38 | 558 |
| SSR Marker | Primer Sequences (5′→3′) | Dye |
|---|---|---|
| ccmp2 | F: GATCCCGGACGTAATCCTG | 6-FAM |
| R: ATCGTACCGAGGGTTCGAAT | ||
| NTCP18 | F: CTGTTCTTTCCATGACCCCTC | HEX |
| R: CCACCTAGCCAAGCCAGA | ||
| NTCP28 | F: TCCAATGGCTTTGGCTA | NED |
| R: AGAAACGAAGGAACCCAC | ||
| NTCP26 | F: GCAATTGCAATGGCTTCTTTA | 6-FAM |
| R: TTTATGTTCGGTGGAAATCACA | ||
| Ibcp5 | F: GCTCTCACGCTCAATTACTTA | HEX |
| R: ATGCTTAATTGACGACCTGT | ||
| Ibcp8 | F: AATAAGTACTTGGCCGTGAA | NED |
| R: CGATTCAAGTAGGCAAAGAG | ||
| Ibcp10 | F: ATATAAGGGGCCATTTTAGG | 6-FAM |
| R: ACGATAGAGGAGAAGGTTCC | ||
| ibcp4 | F: ATCCTGGACGTGAAGAATAA | HEX |
| R: GATGGCTGAGTGGACTAAAG | ||
| ibcp31 | F: AACGGATTTCTCCAATGTA | NED |
| R: ACCTCACCGTTTCAGAAGTA |
| SSR marker | Na 1 | Ne | I | h | uh |
|---|---|---|---|---|---|
| Ibcp10 | 3 | 2.216 | 0.933 | 0.549 | 0.550 |
| Ibcp31 | 1 | monomorphic | |||
| Ibcp4 | 3 | 2.050 | 0.829 | 0.512 | 0.513 |
| Ibcp5 | 2 | 1.266 | 0.365 | 0.210 | 0.210 |
| Ibcp8 | 2 | 1.918 | 0.672 | 0.479 | 0.480 |
| NTCP18 | 2 | 1.174 | 0.280 | 0.148 | 0.148 |
| NTCP26 | 4 | 2.675 | 1.123 | 0.626 | 0.627 |
| NTCP28 | 2 | 1.812 | 0.640 | 0.448 | 0.449 |
| ccmp2 | 3 | 2.072 | 0.839 | 0.517 | 0.518 |
| Mean | 2.4 ± 0.3 | 1.798 ± 0.18 | 0.631 ± 0.12 | 0.388 ± 0.07 | 0.388 ± 0.07 |
| Countries | Na 1 | Ne | I | h | uh |
|---|---|---|---|---|---|
| JPN | 2.22 ± 0.28 | 1.75 ± 0.16 | 0.61 ± 0.10 | 0.39 ± 0.06 | 0.39 ± 0.06 |
| KOR | 2.11 ± 0.20 | 1.64 ± 0.14 | 0.54 ± 0.10 | 0.35± 0.07 | 0.35 ± 0.07 |
| TWN | 2.11 ± 0.20 | 1.77 ± 0.18 | 0.56 ± 0.12 | 0.37 ± 0.08 | 0.38 ± 0.08 |
| USA | 2.11 ± 0.20 | 1.51 ± 0.08 | 0.51 ± 0.08 | 0.31 ± 0.05 | 0.33 ± 0.05 |
| CHN | 2.11 ± 0.20 | 1.45 ± 0.12 | 0.44 ± 0.10 | 0.27 ± 0.06 | 0.28 ± 0.07 |
| PHL | 1.67 ± 0.17 | 1.41 ± 0.13 | 0.36 ± 0.10 | 0.24 ± 0.07 | 0.27 ± 0.08 |
| PER | 1.89 ± 0.20 | 1.56 ± 0.16 | 0.44 ± 0.11 | 0.30 ± 0.08 | 0.30 ± 0.08 |
| PRK | 1.44 ± 0.18 | 1.25 ± 0.12 | 0.23 ± 0.09 | 0.15 ± 0.06 | 0.19 ± 0.08 |
| IDN | 1.78 ± 0.22 | 1.55 ± 0.17 | 0.42 ± 0.12 | 0.29 ± 0.08 | 0.30 ± 0.08 |
| NZE | 1.67 ± 0.24 | 1.30 ± 0.10 | 0.31 ± 0.10 | 0.19 ± 0.06 | 0.22 ± 0.07 |
| Mean | 1.91 ± 0.07 | 1.52 ± 0.04 | 0.44 ± 0.03 | 0.29 ± 0.02 | 0.30 ± 0.02 |
| KOR | JPN | TWN | USA | CHN | PER | IDN | PRK | PHL | NZE | |
|---|---|---|---|---|---|---|---|---|---|---|
| N 1 | 190 | 123 | 73 | 25 | 43 | 50 | 30 | 6 | 9 | 9 |
| H1 | 62 | 40 | 21 | 9 | 9 | 25 | 12 | 1 | 3 | 6 |
| H2 | 16 | 15 | 12 | 1 | 11 | 11 | 2 | 4 | ||
| H3 | 21 | 11 | 2 | 4 | 3 | |||||
| H4 | 4 | 8 | 1 | 8 | 4 | 2 | 3 | |||
| H5 | 15 | 3 | 2 | 3 | ||||||
| H6 | 5 | 11 | 2 | 2 | 1 | |||||
| H7 | 4 | 5 | 8 | 1 | ||||||
| H8 | 6 | 6 | 2 | 2 | ||||||
| H9 | 5 | 4 | 1 | 2 | ||||||
| H10 | 7 | 3 | 1 | 1 | ||||||
| H11 | 4 | 4 | 2 | |||||||
| H12 | 2 | 5 | 2 | |||||||
| H13 | 7 | 1 | ||||||||
| H14 | 2 | 3 | ||||||||
| H15 | 1 | 4 | ||||||||
| H16 | 4 | |||||||||
| H17 | 3 | |||||||||
| H18 | 1 | 1 | 1 | |||||||
| H19 | 1 | 1 | 1 | |||||||
| H20 | 3 | |||||||||
| H21 | 1 | 2 | ||||||||
| H22 | 2 | 1 | ||||||||
| H23 | 1 | 1 | ||||||||
| H24 | 1 | 1 | ||||||||
| H25 | 2 | |||||||||
| H26 | 2 | |||||||||
| H27 | 2 | |||||||||
| H28 | 2 | |||||||||
| H29 | 1 | 1 | ||||||||
| H30 | 2 | |||||||||
| H31 | 2 | |||||||||
| H32 | 1 | 1 | ||||||||
| H33 | 1 | 1 | ||||||||
| PH | 14 | 14 | 6 | 4 | 4 | 2 | ||||
| H | 41 | 31 | 22 | 5 | 13 | 12 | 5 | 8 | 3 | 10 |
| DH | 0.216 | 0.252 | 0.301 | 0.833 | 0.302 | 0.24 | 0.556 | 0.267 | 0.333 | 0.40 |
| Source of Variation | df | SS | MS | Est. Var. | % | PhiPT | Sig. | Nm |
|---|---|---|---|---|---|---|---|---|
| Whole data set | ||||||||
| Among Populations | 9 | 120.09 | 13.34 | 0.236 | 12% | 0.125 | <0.001 | 3.513 |
| Within Populations | 548 | 908.38 | 1.66 | 1.658 | 88% | |||
| Total | 557 | 1028.47 | 1.894 | 100% | ||||
| I | ||||||||
| Among Populations | 3 | 2.73 | 0.91 | 0.046 | 9% | 0.093 | <0.05 | 4.870 |
| Within Populations | 47 | 21.00 | 0.45 | 0.447 | 91% | |||
| Total | 50 | 23.73 | 0.493 | 100% | ||||
| II | ||||||||
| Among Populations | 3 | 1.94 | 0.65 | 0.051 | 13% | 0.134 | ns | 3.244 |
| Within Populations | 25 | 8.33 | 0.33 | 0.333 | 87% | |||
| Total | 28 | 10.28 | 0.385 | 100% | ||||
| III | ||||||||
| Among Populations | 9 | 42.48 | 4.72 | 0.273 | 36% | 0.361 | <0.001 | 0.885 |
| Within Populations | 156 | 75.40 | 0.48 | 0.483 | 64% | |||
| Total | 165 | 117.89 | 0.756 | 100% | ||||
| IV | ||||||||
| Among Populations | 8 | 5.96 | 0.74 | 0.015 | 4% | 0.042 | <0.05 | 11.370 |
| Within Populations | 258 | 89.72 | 0.35 | 0.348 | 96% | |||
| Total | 266 | 95.68 | 0.363 | 100% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.J.; Lee, G.-A.; Lee, J.-R.; Sebastin, R.; Shin, M.-J.; Cho, G.-T.; Hyun, D.Y. Genetic Diversity of Sweet Potato (Ipomoea batatas L. Lam) Germplasms Collected Worldwide Using Chloroplast SSR Markers. Agronomy 2019, 9, 752. https://doi.org/10.3390/agronomy9110752
Lee KJ, Lee G-A, Lee J-R, Sebastin R, Shin M-J, Cho G-T, Hyun DY. Genetic Diversity of Sweet Potato (Ipomoea batatas L. Lam) Germplasms Collected Worldwide Using Chloroplast SSR Markers. Agronomy. 2019; 9(11):752. https://doi.org/10.3390/agronomy9110752
Chicago/Turabian StyleLee, Kyung Jun, Gi-An Lee, Jung-Ro Lee, Raveendar Sebastin, Myoung-Jae Shin, Gyu-Taek Cho, and Do Yoon Hyun. 2019. "Genetic Diversity of Sweet Potato (Ipomoea batatas L. Lam) Germplasms Collected Worldwide Using Chloroplast SSR Markers" Agronomy 9, no. 11: 752. https://doi.org/10.3390/agronomy9110752
APA StyleLee, K. J., Lee, G.-A., Lee, J.-R., Sebastin, R., Shin, M.-J., Cho, G.-T., & Hyun, D. Y. (2019). Genetic Diversity of Sweet Potato (Ipomoea batatas L. Lam) Germplasms Collected Worldwide Using Chloroplast SSR Markers. Agronomy, 9(11), 752. https://doi.org/10.3390/agronomy9110752