A Novel Biostimulant, Belonging to Protein Hydrolysates, Mitigates Abiotic Stress Effects on Maize Seedlings Grown in Hydroponics

Abstract
1. Introduction
2. Materials and Methods
2.1. Maize Seedlings Growth
2.2. Set up of the Novel Biostimulant Concentration to be Supplied to Stressed Maize Plants
2.3. Stress Application and Morphological Analyses
2.4. RNA Extraction, and cDNA Synthesis
2.5. Real Time qPCR
2.6. Data Analysis
3. Results
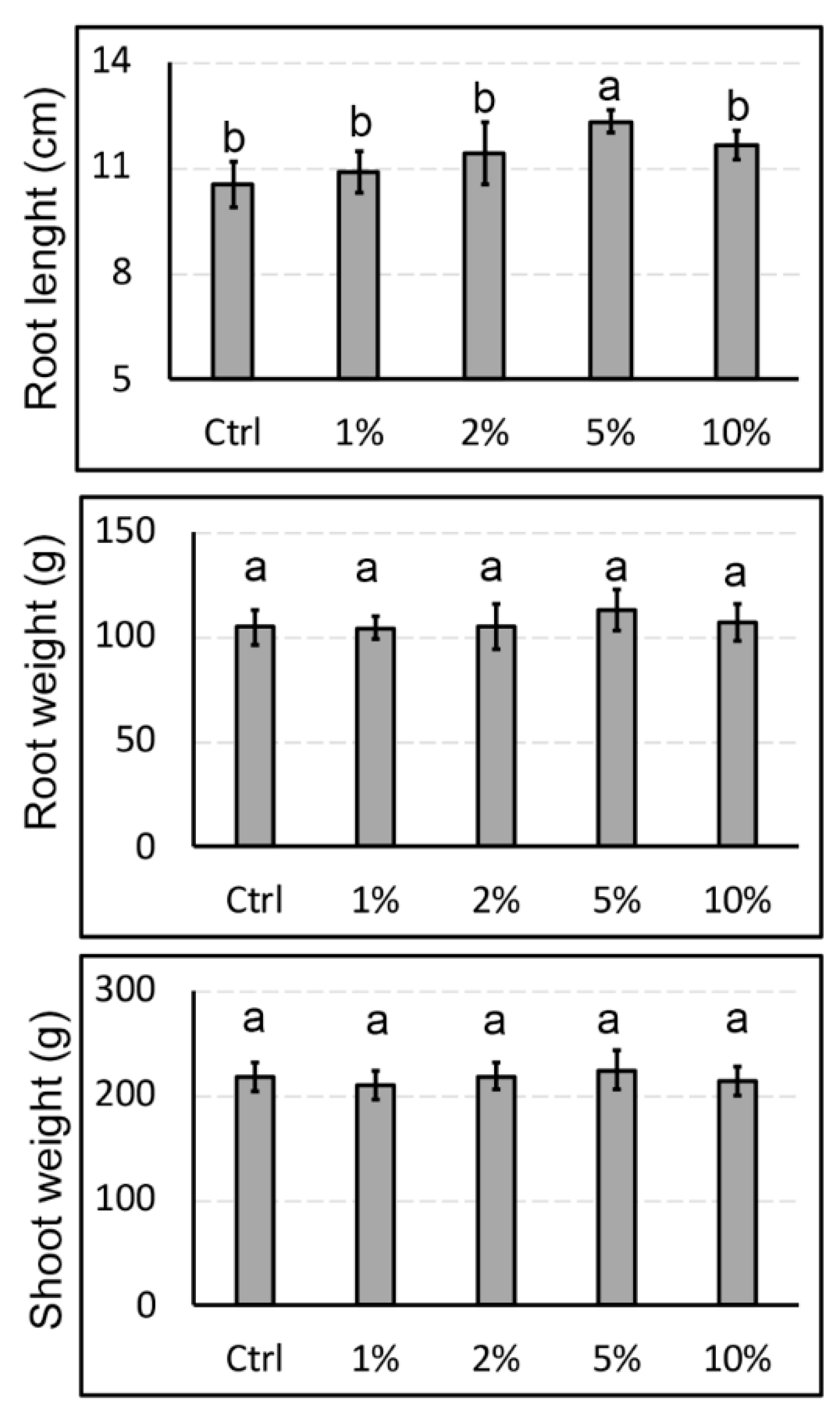
3.1. Choice of the Novel Biostimulant Concentration to be Used for Subsequent Treatments in Hydroponics
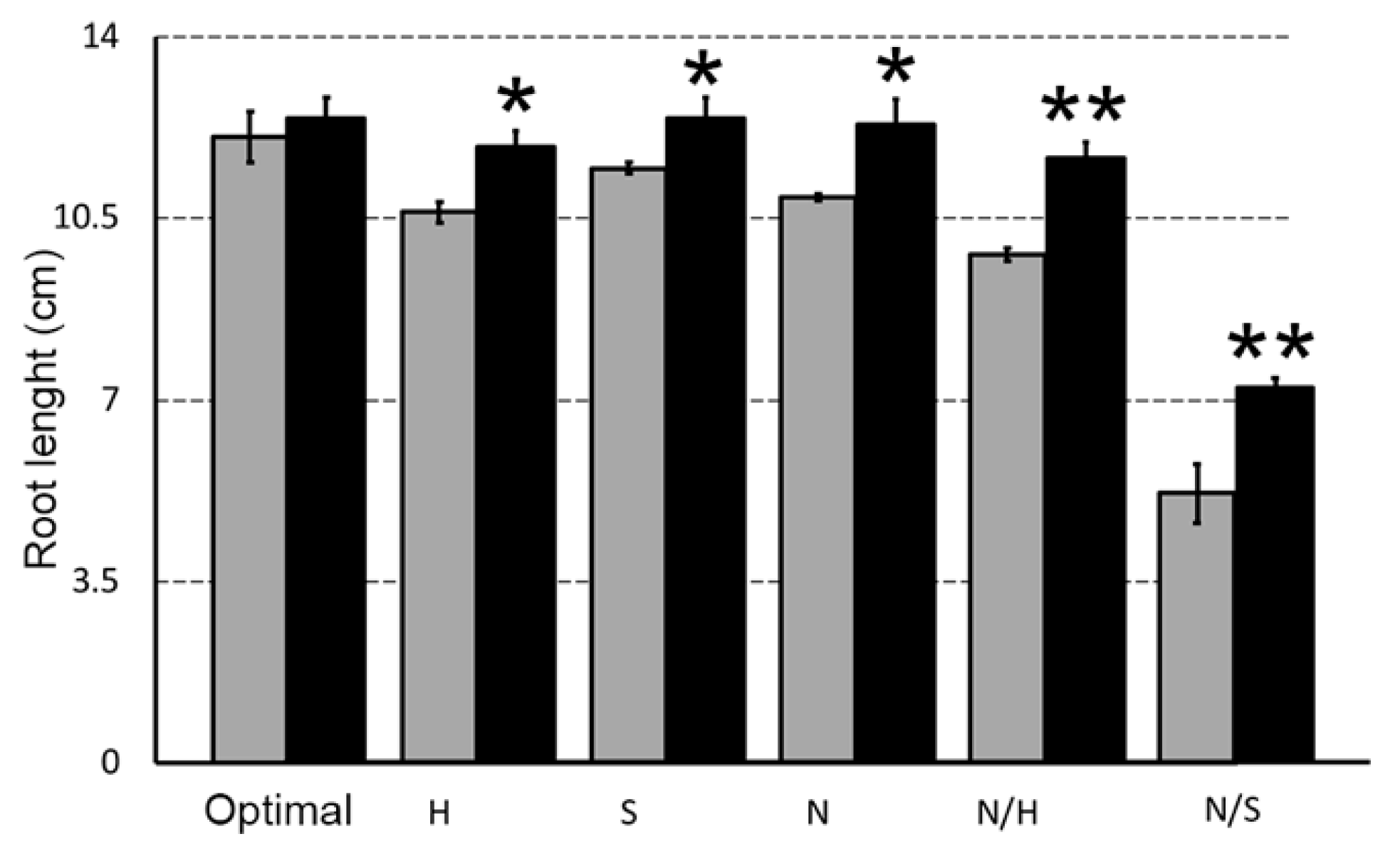
3.2. Biostimulant Effects on Root Length in the Presence of Different Stress
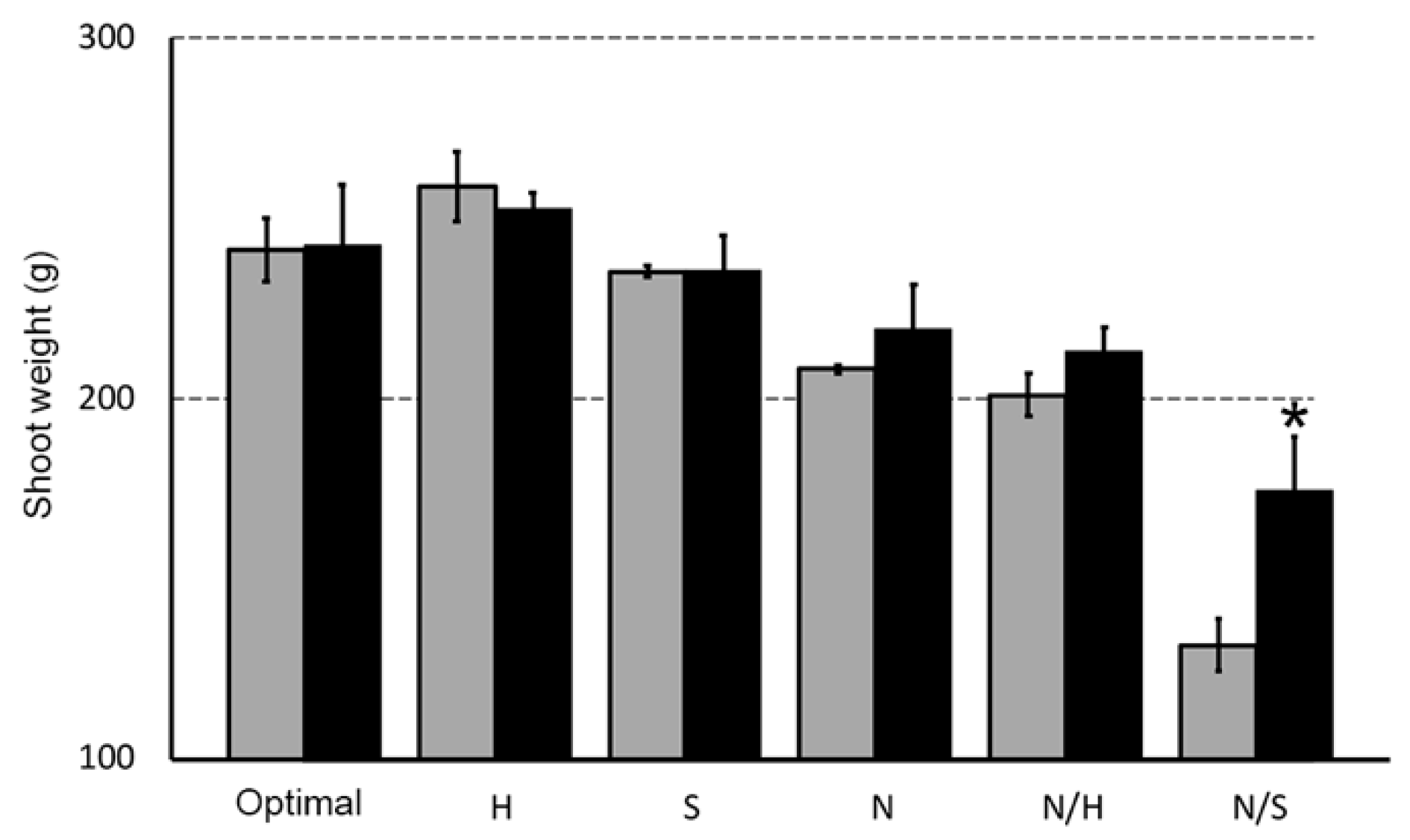
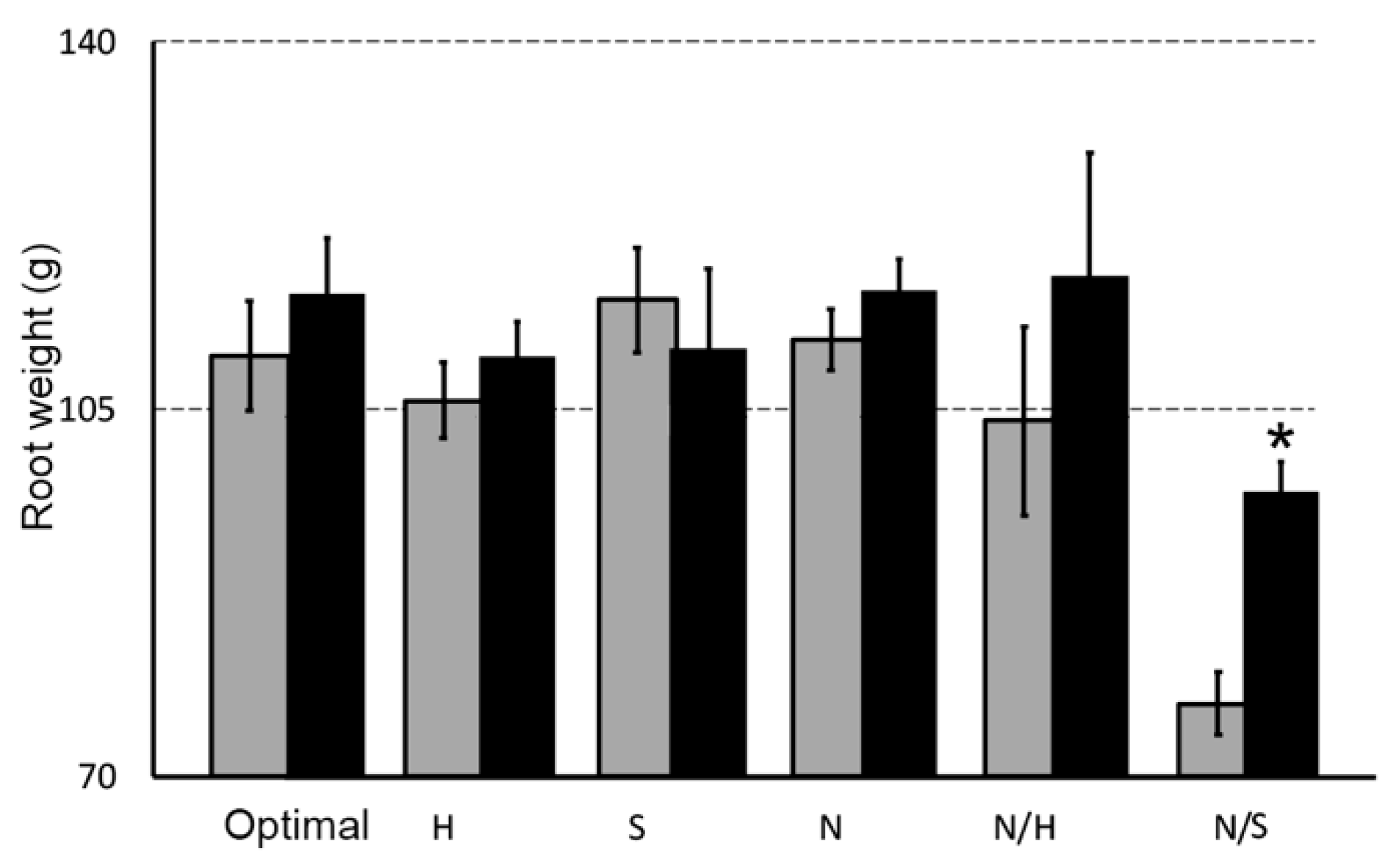
3.3. Biostimulant Effects on Root and Shoot Weight in the Presence of Different Stress
3.4. Biostimulant Regulation of Gene Expression
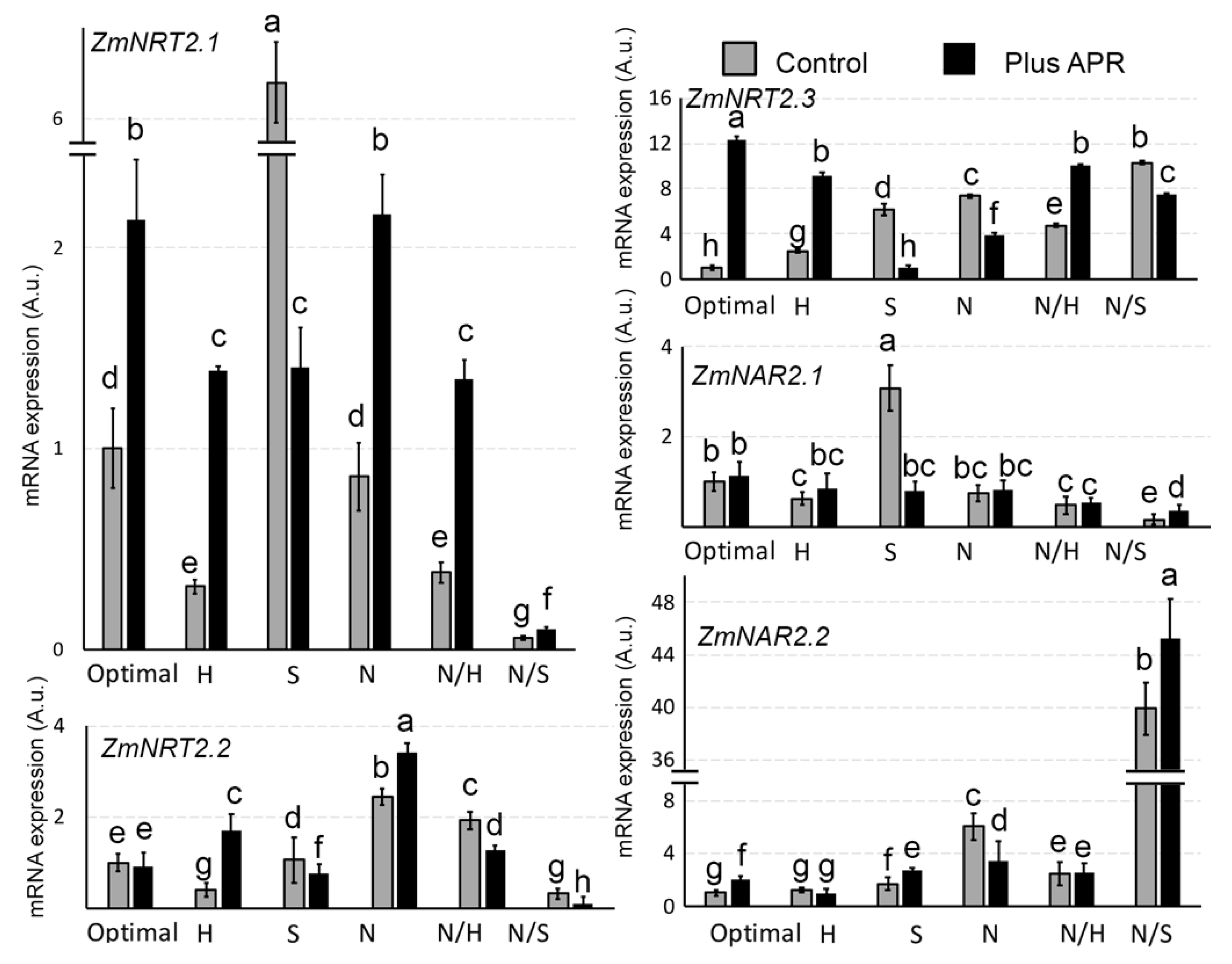
3.5. Biostimulant Effects on ZmNRT2 Genes Expression
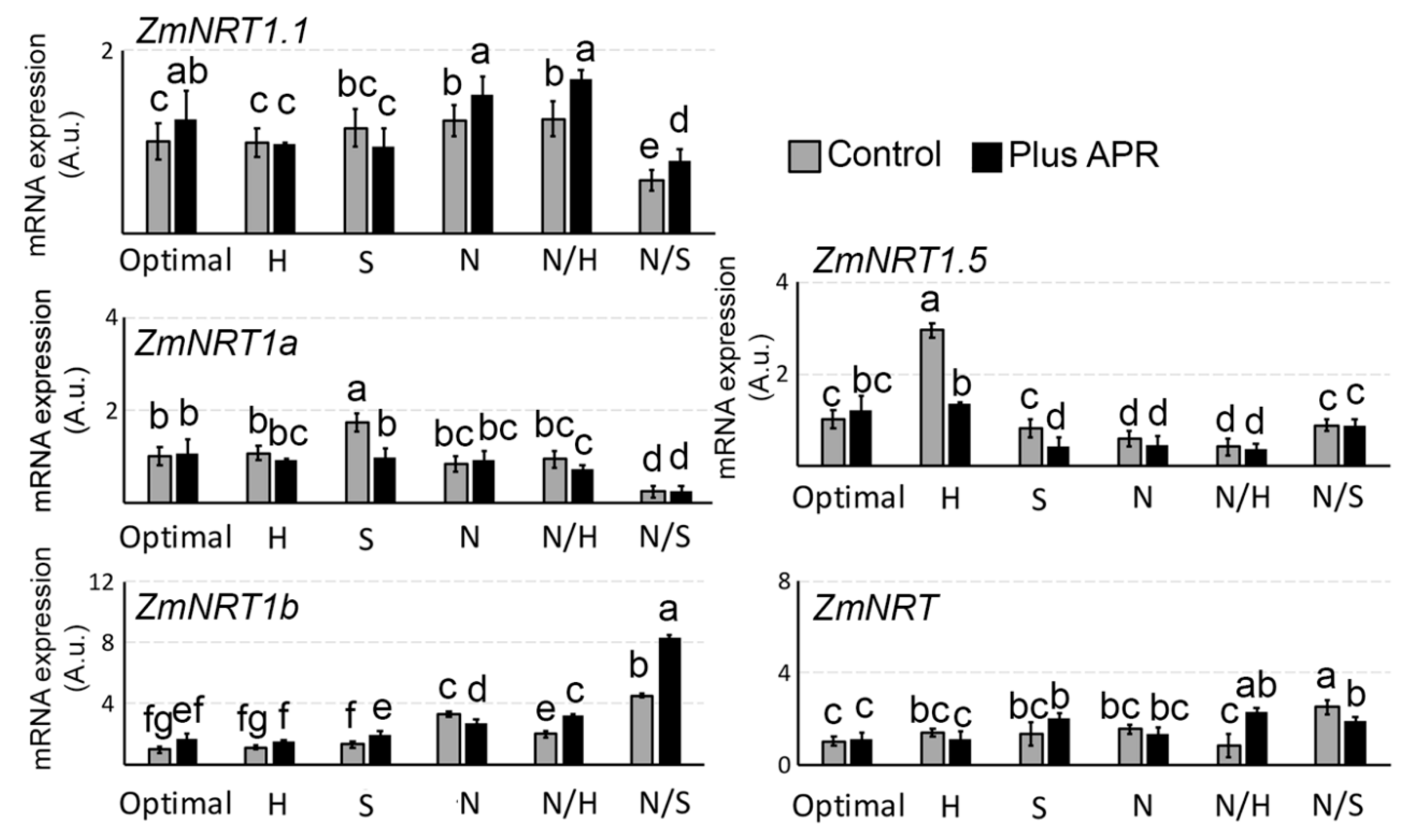
3.6. Biostimulant Effects on NRT1 Genes Expression
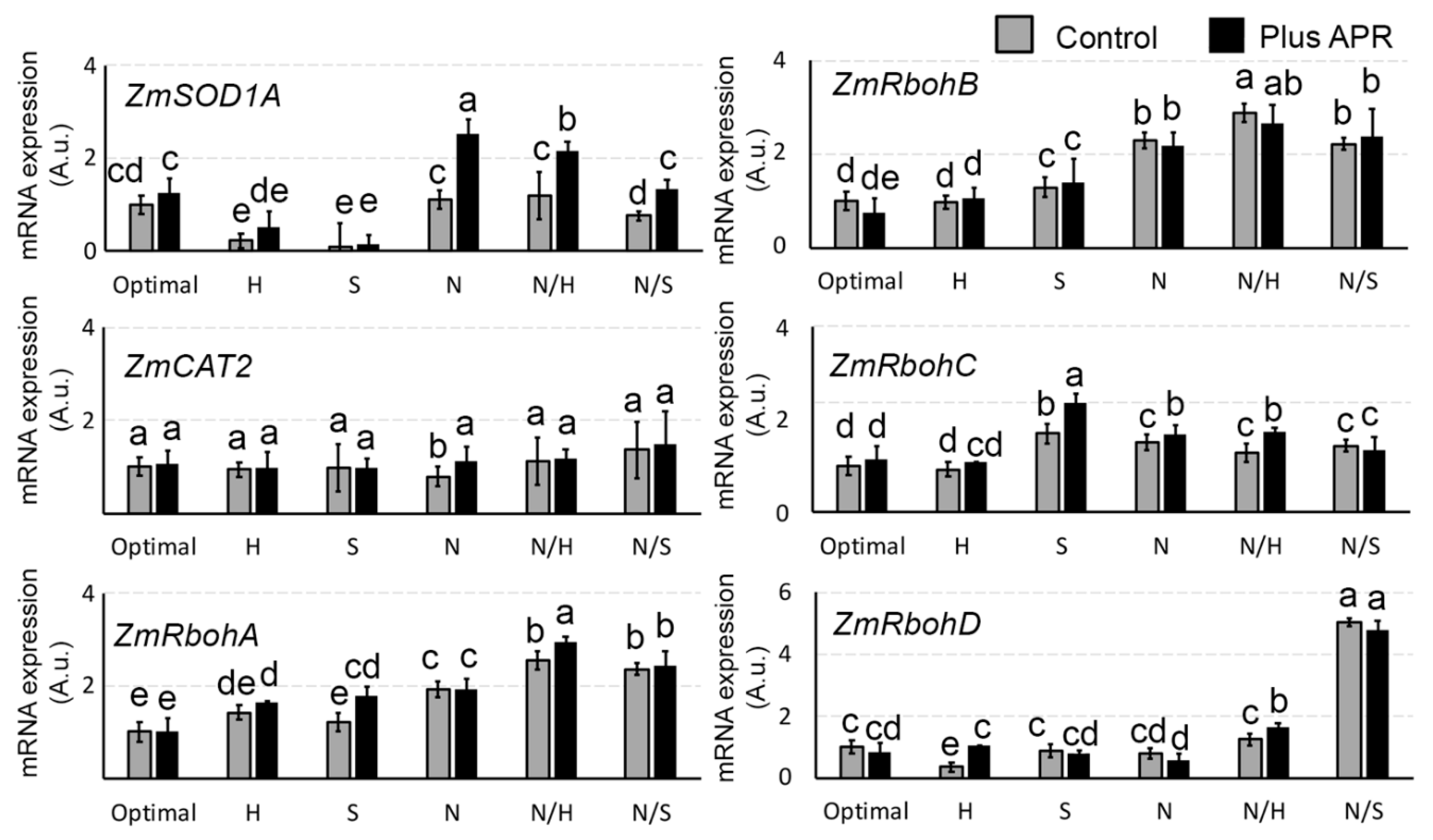
3.7. Biostimulant Effects on ROS Genes Expression
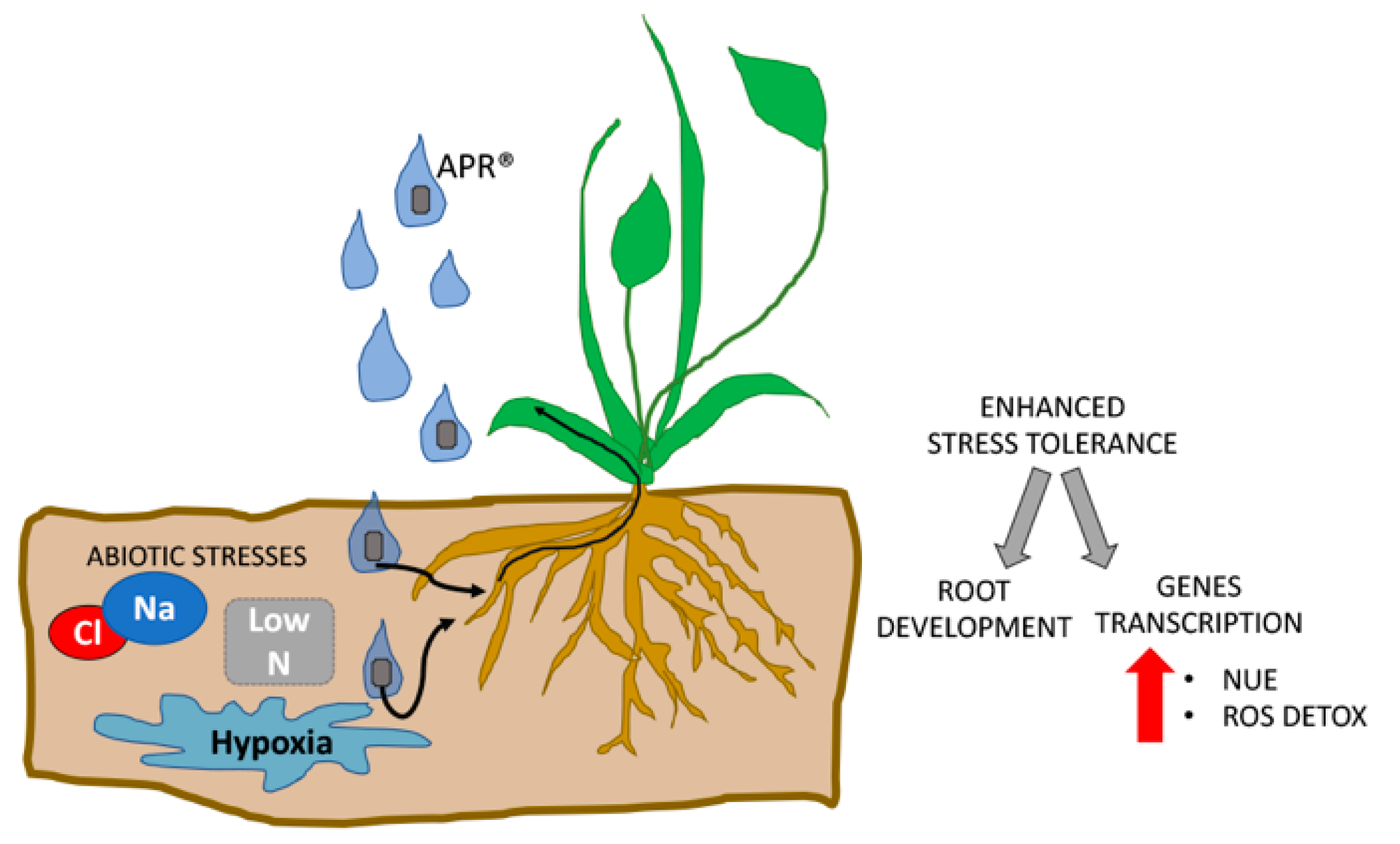
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UN Department of Economic and Social Affairs. Population Division. 2017. World Population Prospects: The 2017 Revision. Key Findings and Advance Tables. Working Paper No. ESA/P/WP/248. Available online: https://www.compassion.com/multimedia/world-population-prospects.pdf (accessed on 7 January 2019).
- UNPD Department of Economic and Social Affairs. World Urbanization Prospects: The 2014 Revision. United Nations Development Programme; 2014: Food and Agriculture Organization of the United Nations (FAO). 2014. The Future of the Food and Agriculture: Trends and Challenges. FAO: Rome, Italy, 2017. Available online: http://www.fao.org/publications/fofa/en/ (accessed on 2 July 2018).
- Rosenzweig, C.; Elliott, J.; Deryng, D.; Ruane, A.C.; Müller, C.; Arneth, A.; Boote, K.J.; Folberth, C.; Glotter, M.; Khabarov, N.; et al. Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc. Natl. Acad. Sci. USA 2014, 111, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Myers, S. Planetary health: Protecting human health on a rapidly changing planet. Lancet 2017, 390, 860–2868. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Tringe, S.G. Building the crops of tomorrow: Advantages of symbiont-based approaches to improving abiotic stress tolerance. Front. Microbiol. 2014, 5, 283. [Google Scholar] [CrossRef] [PubMed]
- Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. Responses to abiotic stresses. In Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1158–1203. [Google Scholar]
- Grover, A.; Kapoor, A.; Lakshmi, O.S.; Agarwal, S.; Sahi, C.; Agarwal, K.; Agarwal, M.; Dubey, H. Understanding molecular alphabets of the plant abiotic stress responses. Curr. Sci. 2001, 80, 206–216. [Google Scholar]
- Singhal, P.; Jan, A.T.; Azam, M.; Haq, Q.M.R. Plant abiotic stress: A prospective strategy of exploiting promoters as alternative to overcome the escalating burden. Front. Life Sci. 2016, 1, 52–63. [Google Scholar] [CrossRef]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Chem. 2018, 19, 6–26. [Google Scholar] [CrossRef]
- Antoniou, C.; Savvides, A.; Christou, A.; Fotopoulos, V. Unravelling chemical priming machinery in plants: The role of reactive oxygen-nitrogen-sulfur species in abiotic stress tolerance enhancement. Curr. Opin. Biotechnol. 2016, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- du Jardin, P. Plant biostimulants: Definition; concept; main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- da Silva, R.R. Bacterial and fungal proteolytic enzymes: Production; catalysis and potential applications. Appl. Biochem. Biotechnol. 2017, 183, 1–19. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.R. Comment on mRNA-Sequencing Analysis Reveals Transcriptional Changes in Root of Maize Seedlings Treated with Two Increasing Concentrations of a New Biostimulant. J. Agric. Food. Chem. 2018, 66, 2061–2062. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Francioso, O.; Ferrari, E.; Schiavon, M.; Nardi, S. Spectroscopic-Chemical Fingerprint and Biostimulant Activity of a Protein-Based Product in Solid Form. Molecules 2018, 23, E1031. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, S.; Manoli, A.; Ravazzolo, L.; Franceschi, C.; Quaggiotti, S. mRNA-Sequencing Analysis Reveals Transcriptional Changes in Root of Maize Seedlings Treated with Two Increasing Concentrations of a New Biostimulant. J. Agric. Food. Chem. 2017, 65, 9956–9969. [Google Scholar] [CrossRef]
- Trevisan, S.; Manoli, A.; Begheldo, M.; Nonis, A.; Enna, M.; Vaccaro, S.; Caporale, G.; Ruperti, B.; Quaggiotti, S. Transcriptome analysis reveals coordinated spatiotemporal regulation of hemoglobin and nitrate reductase in response to nitrate in maize roots. New Phytol. 2011, 192, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, S.; Begheldo, M.; Nonis, A.; Quaggiotti, S. The miRNA-mediated post-transcriptional regulation of maize response to nitrate. Plant Signal. Behav. 2012, 7, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, S.; Manoli, A.; Ravazzolo, L.; Botton, A.; Pivato, M.; Masi, A.; Quaggiotti, S. Nitrate sensing by the maize root apex transition zone: A merged transcriptomic and proteomic survey. J. Exp. Bot. 2015, 66, 3699–3715. [Google Scholar] [CrossRef]
- Trevisan, S.; Trentin, A.R.; Ghisi, R.; Masi, A.; Quaggiotti, S. Nitrate affects transcriptional regulation of UPBEAT1 and ROS localisation in roots of Zea mays L. Physiol. Plant. 2018. [Google Scholar] [CrossRef] [PubMed]
- Quaggiotti, S.; Ruperti, B.; Borsa, P.; Destro, T.; Malagoli, M. Expression of a putative high-affinity NO3- transporter and of an H+-ATPase in relation to whole plant nitrate transport physiology in two maize genotypes differently responsive to low nitrogen availability. J. Exp. Bot. 2003, 54, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects; resistance mechanisms; and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Henry, C.; Bledsoe, S.; Griffiths, C.A.; Paul, M.J.; Kollman, A.; Sakr, S.; Lagrimini, M. Differential role for trehalose metabolism in salt stressed maize. Plant Physiol. 2015, 169, 1072–1089. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Mühling, K.H.; Kutschera, U.; Geilfus, C.M. Salinity Stiffens the Epidermal Cell Walls of Salt-Stressed Maize Leaves: Is the Epidermis Growth-Restricting? PLoS ONE 2015, 10, e0118406. [Google Scholar] [CrossRef]
- Manoli, A.; Sturaro, A.; Trevisan, S.; Quaggiotti, S.; Nonis, A. Evaluation of candidate reference genes for qPCR in maize. J. Plant Physiol. 2012, 169, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Manoli, A.; Begheldo, M.; Genre, A.; Lanfranco, L.; Trevisan, S.; Quaggiotti, S. NO homeostasis is a key regulator of early nitrate perception and root elongation in maize. J. Exp. Bot. 2014, 65, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Nolan, T. Pitfalls of quantitative real-time reverse-transcription polymerase chain reaction. J. Biomol. Tech. 2004, 15, 155–166. [Google Scholar] [PubMed]
- Schaafsma, G. Safety of protein hydrolysates; fractions thereof and bioactive peptides in human nutrition. Eur. J. Clin. Nutr. 2009, 63, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- du Jardin, P. The Science of Plant Biostimulants—A bibliographic analysis. Ad hoc Study Report to the European Commission DG ENTR 2012. Available online: https://orbi.uliege.be/handle/2268/169257 (accessed on 7 January 2019).
- Kasparkova, V.; Kolomaznik, K.; Burketova, L.; Sasek, V.; Simek, L. Characterization of low-molecular weight collagen hydrolysates prepared by combination of enzymatic and acid hydrolysis. J. Am. Leather Chem. Assoc. 2009, 104, 46–51. [Google Scholar]
- Pecha, J.; Fürst, T.; Kolomaznik, K.; Friebrova, V.; Svoboda, P. Protein biostimulant foliar uptake modeling: The impact of climatic conditions. AIChE J. 2012, 58, 2010–2019. [Google Scholar] [CrossRef]
- Baglieri, A.; Cadili, V.; Monterumici, C.M.; Gennari, M.; Tabasso, S.; Montoneri, E.; Nardi, S.; Negre, M. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production; chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 96, 28–38. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil. 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Life Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress; the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Malamy, J.E. Intrinsic and environmental response path- ways that regulate root system architecture. Plant Cell Environ. 2005, 28, 67–77. [Google Scholar] [CrossRef]
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant root growth; architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar] [CrossRef]
- Kong, X.; Liu, G.; Liu, J.; Ding, Z. The Root Transition Zone: A Hot Spot for Signal Crosstalk. Trends Plant Sci. 2018, 23, 403–409. [Google Scholar] [CrossRef]
- Alarcon, M.V.; Salguero, J. Transition zone cells reach G2 phase before initiating elongation in maize root apex. Biol. Open 2017, 6, 909–913. [Google Scholar] [CrossRef]
- Yang, Z.B.; Geng, X.; He, C.; Zhang, F.; Wang, R.; Horst, W.J.; Ding, Z. TAA1-regulated local auxin biosynthesis in the root-apex transition zone mediates the aluminum-induced inhibition of root growth in Arabidopsis. Plant Cell 2014, 26, 2889–2904. [Google Scholar] [CrossRef]
- Baluska, F.; Volkmann, D.; Barlow, P.W. Specialized zones of development in roots: View from the cellular level. Plant Physiol. 1996, 112, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Baluska, F.; Mancuso, S.; Volkmann, D.; Barlow, P.W. Root apex transition zone: A signalling-response nexus in the root. Trends Plant Sci. 2010, 15, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Baluska, F.; Mancuso, S. Root apex transition zone as oscillatory zone. Front. Plant Sci. 2013, 4, 354. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A Vegetal Biopolymer-Based Biostimulant Promoted Root Growth in Melon While Triggering Brassinosteroids and Stress-Related Compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein Hydrolysate Stimulates Growth in Tomato Coupled With N-Dependent Gene Expression Involved in N Assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef]
- Plett, D.C.; Holtham, L.R.; Okamoto, M.; Garnett, T.P. Nitrate uptake and its regulation in relation to improving nitrogen use efficiency in cereals. Semin. Cell Dev. Biol. 2018, 74, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xia, M.X.; Chen, J.; Yuan, R.; Deng, F.N.; Shen, F.F. Gene Expression Characteristics and Regulation Mechanisms of Superoxide Dismutase and Its Physiological Roles in Plants under Stress. Biochemistry 2016, 81, 465–480. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maize GDB Accession ID | Code | Description | |
|---|---|---|---|
| HATS | Zm00001d054057 | ZmNRT2.1 | High affinity nitrate transporter |
| Zm00001d054060 | ZmNRT2.2 | High affinity nitrate transporter | |
| Zm00001d014976 | ZmNRT2.3 | High affinity nitrate transporter | |
| Zm00001d017095 | ZmNAR2.1 | High affinity nitrate transporter | |
| Zm00001d003287 | ZmNAR2.2 | High affinity nitrate transporter | |
| LATS | Zm00001d024587 | ZmNRT1.1 | Nitrate transporter |
| Zm00001d029932 | ZmNRT1a | Dual-affinity nitrate transporter | |
| Zm00001d036941 | ZmNRT1b | Nitrate transporter | |
| Zm00001d017666 | ZmNRT1.5 | Nitrate transporter | |
| Zm00001d007785 | ZmNRT | Nitrate and chloride transporter | |
| ROS | Zm00001d042961 | ZmRbohA | Respiratory burst oxidase protein A |
| Zm00001d043543 | ZmRbohB | Respiratory burst oxidase protein B | |
| Zm00001d038762 | ZmRbohC | Respiratory burst oxidase protein C | |
| Zm00001d052653 | ZmRbohD | Respiratory burst oxidase protein D | |
| Zm00001d031908 | ZmSOD | Superoxide dismutase [Cu-Zn] | |
| Zm00001d027511 | ZmCAT2 | Catalase 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevisan, S.; Manoli, A.; Quaggiotti, S. A Novel Biostimulant, Belonging to Protein Hydrolysates, Mitigates Abiotic Stress Effects on Maize Seedlings Grown in Hydroponics. Agronomy 2019, 9, 28. https://doi.org/10.3390/agronomy9010028
Trevisan S, Manoli A, Quaggiotti S. A Novel Biostimulant, Belonging to Protein Hydrolysates, Mitigates Abiotic Stress Effects on Maize Seedlings Grown in Hydroponics. Agronomy. 2019; 9(1):28. https://doi.org/10.3390/agronomy9010028
Chicago/Turabian StyleTrevisan, Sara, Alessandro Manoli, and Silvia Quaggiotti. 2019. "A Novel Biostimulant, Belonging to Protein Hydrolysates, Mitigates Abiotic Stress Effects on Maize Seedlings Grown in Hydroponics" Agronomy 9, no. 1: 28. https://doi.org/10.3390/agronomy9010028
APA StyleTrevisan, S., Manoli, A., & Quaggiotti, S. (2019). A Novel Biostimulant, Belonging to Protein Hydrolysates, Mitigates Abiotic Stress Effects on Maize Seedlings Grown in Hydroponics. Agronomy, 9(1), 28. https://doi.org/10.3390/agronomy9010028