Plant Protection by Benzoxazinoids—Recent Insights into Biosynthesis and Function


Abstract
1. Introduction
2. Structure of Benzoxazinoids and Chemical Properties
3. Distribution of Benzoxazinoids
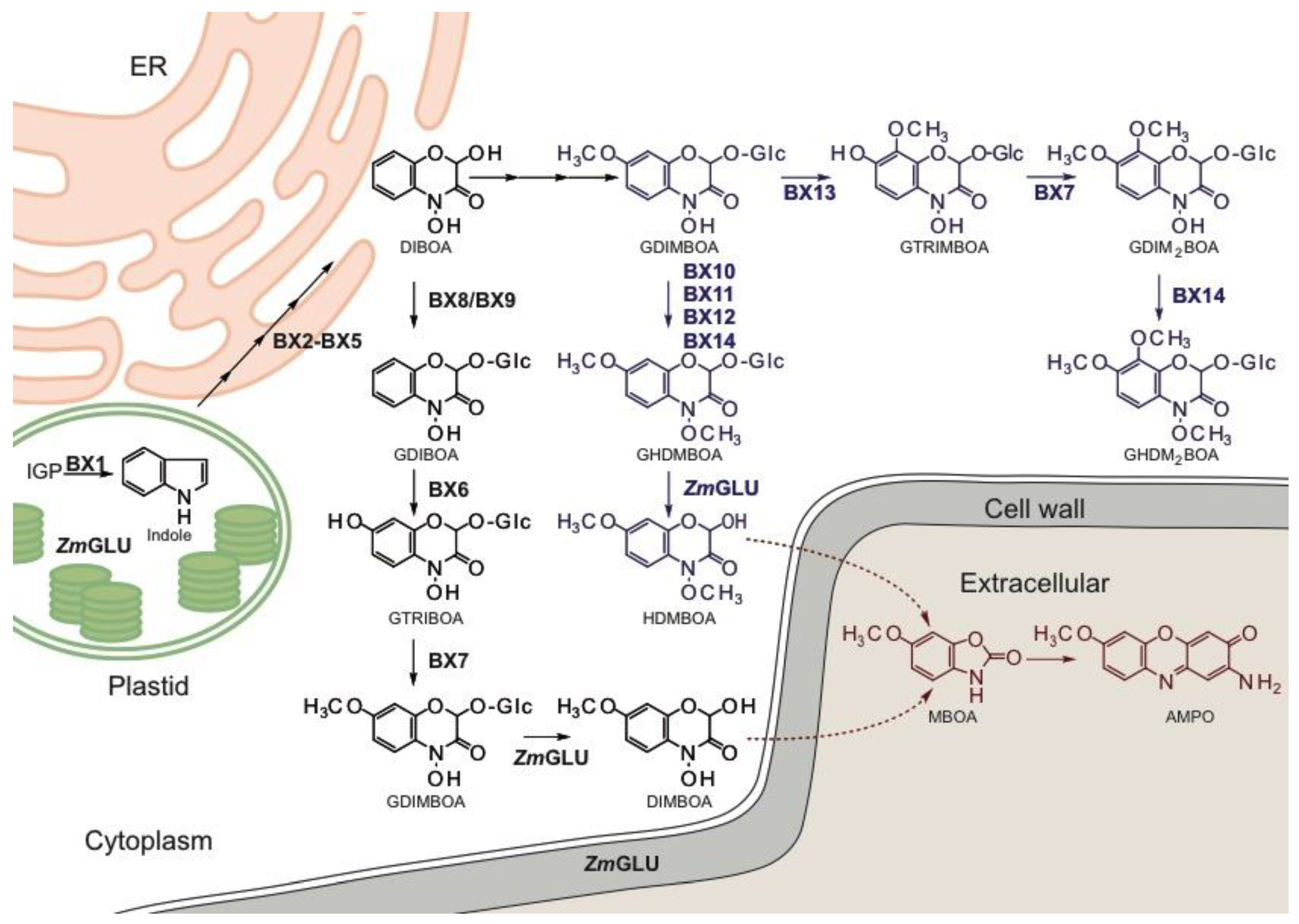
4. Biosynthesis
5. Biological Interaction
5.1. Human Health Clinical Studies, Detrimental and Health-Promoting Effects
5.2. Interactions with Microbes
5.3. Benzoxazinoids in the Soil: Allelopathy and the Chelating of Metals
5.4. Insects
5.4.1. Chewing Insects
5.4.2. Aphids
5.4.3. Specialist Insects
5.5. BXs as Signalling Components
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: Nature’s diversity and ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Strack, D. Phytochemistry meets genome analysis, and beyond. Phytochemistry 2003, 62, 815–816. [Google Scholar] [CrossRef]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Barry, D.; Darrah, L.L. Effect of research on commercial hybrid maize resistance to European corn borer (lepidoptera: Pyralidae). J. Econ. Entomol. 1991, 84, 1053–1059. [Google Scholar] [CrossRef]
- Grombacher, A.W.; Russell, W.A.; Guthrie, W.D. Resistance to first-generation European corn borer (lepidoptera: Pyralidae) and DIMBOA concentration in midwhorl leaves of the BS9 maize synthetic. J. Kansas Entomol. Soc. 1989, 62, 103–107. [Google Scholar]
- Klun, J.A.; Guthrie, W.D.; Hallauer, A.R.; Russell, W.A. Genetic nature of the concentration of 2,4-dihydroxy-7-methoxy 2h-l,4-benzoxazin-3(4h)-one and resistance to the European corn borer in a diallel set of eleven maize inbreds. Crop Sci. 1970, 10, 87–90. [Google Scholar] [CrossRef]
- Pentzold, S.; Zagrobelny, M.; Rook, F.; Bak, S. How insects overcome two-component plant chemical defence: Plant beta-glucosidases as the main target for herbivore adaptation. Biol. Rev. 2014, 89, 531–551. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Richter, A.; Jander, G. Beyond defense: Multiple functions of benzoxazinoids in maize metabolism. Plant Cell Physiol 2018. [Google Scholar] [CrossRef] [PubMed]
- Schulz, M.; Marocco, A.; Tabaglio, V.; Macias, F.A.; Molinillo, J.M. Benzoxazinoids in rye allelopathy—From discovery to application in sustainable weed control and organic farming. J. Chem. Ecol. 2013, 39, 154–174. [Google Scholar] [CrossRef] [PubMed]
- Wouters, F.C.; Blanchette, B.; Gershenzon, J.; Vassao, D.G. Plant defense and herbivore counter-defense: Benzoxazinoids and insect herbivores. Phytochem. Rev.: Proc. Phytochem. Soc. Eur. 2016, 15, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Hietala, P.K.; Virtanen, A.I. Precursors of benzoxazolinone in rye plants. II. Precursor I, the glucoside. Acta Chem. Scand. 1960, 14, 502–504. [Google Scholar] [CrossRef]
- Pérez, F.J.; Niemeyer, H.M. Reaction of DIMBOA with amines. Phytochemistry 1989, 28, 1831–1834. [Google Scholar] [CrossRef]
- Hamilton, R.H. A corn mutant deficient in 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3- one with an altered tolerance of atrazine. Weeds 1964, 12, 27–30. [Google Scholar] [CrossRef]
- Wenger, K.; Bigler, L.; Suter, M.J.; Schonenberger, R.; Gupta, S.K.; Schulin, R. Effect of corn root exudates on the degradation of atrazine and its chlorinated metabolites in soils. J. Environ. Qual. 2005, 34, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Shudo, K. Chemistry of biologically active benzoxazinoids. Phytochemistry 1996, 43, 551–559. [Google Scholar] [CrossRef]
- Hofmann, A.; Sicker, D. A formylating agent by dehydration of the natural product dimboa. J. Nat. Prod. 1999, 62, 1151–1153. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Sellars, J.D.; Kenwright, A.M.; Steel, P.G. The maize benzoxazinone dimboa reacts with glutathione and other thiols to form spirocyclic adducts. Phytochemistry 2012, 77, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Maresh, J.; Zhang, J.; Lynn, D.G. The innate immunity of maize and the dynamic chemical strategies regulating two-component signal transduction in agrobacterium tumefaciens. ACS Chem Biol 2006, 1, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, L.; Niemeyer, H.M.; Pérez, F.J. Reaction of DIMBOA, a resistance factor from cereals, with α-chymotrypsin. Phytochemistry 1990, 29, 1429–1432. [Google Scholar] [CrossRef]
- Cuevas, L.; Niemeyer, H.M. Effect of hydroxamic acids from cereals on aphid cholinesterases. Phytochemistry 1993, 34, 983–985. [Google Scholar] [CrossRef]
- Friebe, A.; Roth, U.; Kück, P.; Schnabl, H.; Schulz, M. Effects of 2,4-dihydroxy-1,4-benzoxazin-3-ones on the activity of plasma membrane H+-ATPase. Phytochemistry 1997, 44, 979–983. [Google Scholar] [CrossRef]
- Sicker, D.; Frey, M.; Schulz, M.; Gierl, A. Role of natural benzoxazinones in the survival strategy of plants. Int. Rev. Cytol. 2000, 198, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, H.M. Hydroxamic acids derived from 2-hydroxy-2h-1,4-benzoxazin-3(4h)-one: Key defense chemicals of cereals. J. Agric. Food Chem. 2009, 57, 1677–1696. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; McMullen, M.D.; Bauer, E.; Schon, C.C.; Gierl, A.; Frey, M. Prolonged expression of the bx1 signature enzyme is associated with a recombination hotspot in the benzoxazinoid gene cluster in Zea mays. J. Exp. Bot. 2015, 66, 3917–3930. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Holland, J.B.; McMullen, M.D.; Buckler, E.S. Genetic design and statistical power of nested association mapping in maize. Genetics 2008, 178, 539. [Google Scholar] [CrossRef] [PubMed]
- Hanhineva, K.; Rogachev, I.; Aura, A.M.; Aharoni, A.; Poutanen, K.; Mykkanen, H. Qualitative characterization of benzoxazinoid derivatives in whole grain rye and wheat by lc-ms metabolite profiling. J. Agric. Food Chem. 2011, 59, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Pekkinen, J.; Katina, K.; Poutanen, K.; Hanhineva, K. Glycosylated benzoxazinoids are degraded during fermentation of wheat bran. J. Agric. Food Chem. 2015, 63, 5943–5949. [Google Scholar] [CrossRef] [PubMed]
- Schullehner, K.; Dick, R.; Vitzthum, F.; Schwab, W.; Brandt, W.; Frey, M.; Gierl, A. Benzoxazinoid biosynthesis in dicot plants. Phytochemistry 2008, 69, 2668–2677. [Google Scholar] [CrossRef] [PubMed]
- Bravo, H.R.; Copaja, S.V. Contents and morphological distribution of 2,4-dihydroxy-l,4-benzoxazin-3-one and 2-benzoxazolinone in Acanthus mollis in relation to protection from larvae of Pseudaletia impuncta. Ann. Appl. Biol. 2002, 140, 129–132. [Google Scholar] [CrossRef]
- Frey, M.; Chomet, P.; Glawischnig, E.; Stettner, C.; Grun, S.; Winklmair, A.; Eisenreich, W.; Bacher, A.; Meeley, R.B.; Briggs, S.P.; et al. Analysis of a chemical plant defense mechanism in grasses. Science 1997, 277, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Von Rad, U.; Huttl, R.; Lottspeich, F.; Gierl, A.; Frey, M. Two glucosyltransferases are involved in detoxification of benzoxazinoids in maize. Plant J. 2001, 28, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Jonczyk, R.; Schmidt, H.; Osterrieder, A.; Fiesselmann, A.; Schullehner, K.; Haslbeck, M.; Sicker, D.; Hofmann, D.; Yalpani, N.; Simmons, C.; et al. Elucidation of the final reactions of DIMBOA-glucoside biosynthesis in maize: Characterization of Bx6 and Bx7. Plant Physiol. 2008, 146, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Ishihara, A.; Imaishi, H.; Endo, T.; Ohkawa, H.; Iwamura, H. Molecular characterization and chromosomal localization of cytochrome P450 genes involved in the biosynthesis of cyclic hydroxamic acids in hexaploid wheat. Mol. Genet. Genom. 2002, 267, 210–217. [Google Scholar] [CrossRef]
- Nomura, T.; Ishihara, A.; Yanagita, R.C.; Endo, T.R.; Iwamura, H. Three genomes signalling differentially contribute to the biosynthesis of benzoxazinones in hexaploid wheat. Proc. Natl Acad. Sci. USA 2005, 102, 16490–16495. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Nasuda, S.; Kawaura, K.; Ogihara, Y.; Kato, N.; Sato, F.; Kojima, T.; Toyoda, A.; Iwamura, H.; Endo, T.R. Structures of the three homoeologous loci of wheat benzoxazinone biosynthetic genes TaBx3 and TaBx4 and characterization of their promoter sequences. Theor. Appl. Genet. 2008, 116, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Sue, M.; Nakamura, C.; Nomura, T. Dispersed benzoxazinone gene cluster: Molecular characterization and chromosomal localization of glucosyltransferase and glucosidase genes in wheat and rye. Plant Physiol. 2011, 157, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Bakera, B.; Makowska, B.; Groszyk, J.; Niziolek, M.; Orczyk, W.; Bolibok-Bragoszewska, H.; Hromada-Judycka, A.; Rakoczy-Trojanowska, M. Structural characteristics of ScBx genes controlling the biosynthesis of hydroxamic acids in rye (Secale cereale L.). J. Appl. Genet. 2015, 56, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Rakoczy-Trojanowska, M.; Orczyk, W.; Krajewski, P.; Bocianowski, J.; Stochmal, A.; Kowalczyk, M. Scbx gene based association analysis of hydroxamate content in rye (Secale cereale L.). J. Appl. Genet. 2017, 58, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tanwir, F.; Dionisio, G.; Adhikari, K.B.; Fomsgaard, I.S.; Gregersen, P.L. Biosynthesis and chemical transformation of benzoxazinoids in rye during seed germination and the identification of a rye Bx6-like gene. Phytochemistry 2017, 140, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Kriechbaumer, V.; Weigang, L.; Fießelmann, A.; Letzel, T.; Frey, M.; Gierl, A.; Glawischnig, E. Characterisation of the tryptophan synthase alpha subunit in maize. BMC Plant Biol. 2008, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Hyde, C.C.; Ahmed, S.A.; Padlan, E.A.; Miles, E.W.; Davies, D.R. Three-dimensional structure of the tryptophan synthase alpha 2 beta 2 multienzyme complex from Salmonella typhimurium. J. Biol. Chem. 1988, 263, 17857–17871. [Google Scholar] [PubMed]
- Spiteller, P.; Glawischnig, E.; Gierl, A.; Steglich, W. Studies on the biosynthesis of 2-hydroxy-1,4-benzoxazin-3-one (HBOA) from 3-hydroxyindolin-2-one in Zea mays. Phytochemistry 2001, 57, 373–376. [Google Scholar] [CrossRef]
- Oikawa, A.; Ishihara, A.; Tanaka, C.; Mori, N.; Tsuda, M.; Iwamura, H. Accumulation of HDMBOA-glc is induced by biotic stresses prior to the release of MBOA in maize leaves. Phytochemistry 2004, 65, 2995–3001. [Google Scholar] [CrossRef] [PubMed]
- Meihls, L.N.; Handrick, V.; Glauser, G.; Barbier, H.; Kaur, H.; Haribal, M.M.; Lipka, A.E.; Gershenzon, J.; Buckler, E.S.; Erb, M.; et al. Natural variation in maize aphid resistance is associated with 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one glucoside methyl- transferase activity. Plant Cell 2013, 25, 2341–2355. [Google Scholar] [CrossRef] [PubMed]
- Tzin, V.; Hojo, Y.; Strickler, S.R.; Bartsch, L.J.; Archer, C.M.; Ahern, K.R.; Zhou, S.; Christensen, S.A.; Galis, I.; Mueller, L.A.; et al. Rapid defense responses in maize leaves induced by Spodoptera exigua caterpillar feeding. J. Exp. Bot. 2017, 68, 4709–4723. [Google Scholar] [CrossRef] [PubMed]
- Handrick, V.; Robert, C.A.M.; Ahern, K.R.; Zhou, S.; Machado, R.A.R.; Maag, D.; Glauser, G.; Fernandez-Penny, F.E.; Chandran, J.N.; Rodgers-Melnik, E.; et al. Biosynthesis of 8-O-methylated benzoxazinoid defense compounds in maize. Plant Cell 2016, 28, 1682–1700. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.; Schullehner, K.; Dick, R.; Fiesselmann, A.; Gierl, A. Benzoxazinoid biosynthesis, a model for evolution of secondary metabolic pathways in plants. Phytochemistry 2009, 70, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Oka, R.; Zicola, J.; Weber, B.; Anderson, S.N.; Hodgman, C.; Gent, J.I.; Wesselink, J.J.; Springer, N.M.; Hoefsloot, H.C.J.; Turck, F.; et al. Genome-wide mapping of transcriptional enhancer candidates using DNA and chromatin features in maize. Genome Biol. 2017, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Butron, A.; Chen, Y.C.; Rottinghaus, G.E.; McMullen, M.D. Genetic variation at Bx1 controls DIMBOA content in maize. Theor. Appl. Genet. 2010, 120, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Nutzmann, H.W.; Huang, A.; Osbourn, A. Plant metabolic clusters—From genetics to genomics. New Phytol. 2016, 211, 771–789. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Osbourn, A. Computational genomic identification and functional reconstitution of plant natural product biosynthetic pathways. Nat. Prod. Rep. 2016, 33, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Wisecaver, J.H.; Borowsky, A.T.; Tzin, V.; Jander, G.; Kliebenstein, D.J.; Rokas, A. A global coexpression network approach for connecting genes to specialized metabolic pathways in plants. Plant Cell 2017, 29, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Tzin, V.; Fernandez-Pozo, N.; Richter, A.; Schmelz, E.A.; Schoettner, M.; Schäfer, M.; Ahern, K.R.; Meihls, L.N.; Kaur, H.; Huffaker, A.; et al. Dynamic maize responses to aphid feeding are revealed by a time series of transcriptomic and metabolomic assays. Plant Physiol. 2015, 169, 1727–1743. [Google Scholar] [CrossRef] [PubMed]
- Esen, A. Purification and partial characterization of maize (Zea mays L.) beta-glucosidase. Plant Physiol. 1992, 98, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Czjzek, M.; Cicek, M.; Zamboni, V.; Burmeister, W.P.; Bevan, D.R.; Henrissat, B.; Esen, A. Crystal structure of a monocotyledon (Maize ZmGLU1) beta-glucosidase and a model of its complex with p-nitrophenyl beta-d-thioglucoside. Biochem. J. 2001, 354, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Nikus, J.; Daniel, G.; Jonsson, L.M. Subcellular localization of beta-glucosidase in rye, maize and wheat seedlings. Physiol. Plant. 2001, 111, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Sue, M.; Yamazaki, K.; Yajima, S.; Nomura, T.; Matsukawa, T.; Iwamura, H.; Miyamoto, T. Molecular and structural characterization of hexameric beta-d-glucosidases in wheat and rye. Plant Physiol. 2006, 141, 1237–1247. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huffaker, A.; Dafoe, N.J.; Schmelz, E.A. Zmpep1, an ortholog of Arabidopsis elicitor peptide 1, regulates maize innate immunity and enhances disease resistance. Plant Physiol. 2011, 155, 1325–1338. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Pearce, G.; Veyrat, N.; Erb, M.; Turlings, T.C.J.; Sartor, R.; Shen, Z.; Briggs, S.P.; Vaughan, M.M.; Alborn, H.T.; et al. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. Proc. Nat.l Acad. Sci. 2013, 110, 5707. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, A.; Ishihara, A.; Hasegawa, M.; Kodama, O.; Iwamura, H. Induced accumulation of 2-hydroxy-4,7-dimethoxy-1,4-benzoxazin-3-one glucoside (HDMBOA-glc) in maize leaves. Phytochemistry 2001, 56, 669–675. [Google Scholar] [CrossRef]
- Oikawa, A.; Ishihara, A.; Iwamura, H. Induction of HDMBOA-glc accumulation and DIMBOA-glc 4-O-methyltransferase by jasmonic acid in poaceous plants. Phytochemistry 2002, 61, 331–337. [Google Scholar] [CrossRef]
- Dick, R.; Rattei, T.; Haslbeck, M.; Schwab, W.; Gierl, A.; Frey, M. Comparative analysis of benzoxazinoid biosynthesis in monocots and dicots: Independent recruitment of stabilization and activation functions. Plant Cell 2012, 24, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Hannemann, L.; Lucaciu, C.R.; Sharma, S.; Rattei, T.; Mayer, K.F.X.; Gierl, A.; Frey, M. A promiscuous beta-glucosidase is involved in benzoxazinoid deglycosylation in Lamium galeobdolon. Phytochemistry 2018. Paper under review. [Google Scholar]
- Pihlava, J.M.; Kurtelius, T. Determination of benzoxazinoids in wheat and rye beers by HPLC-DAD and UPLC-QTOF MS. Food chem. 2016, 204, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, K.B.; Tanwir, F.; Gregersen, P.L.; Steffensen, S.K.; Jensen, B.M.; Poulsen, L.K.; Nielsen, C.H.; Hoyer, S.; Borre, M.; Fomsgaard, I.S. Benzoxazinoids: Cereal phytochemicals with putative therapeutic and health-protecting properties. Mol. Nutr. Food Res. 2015, 59, 1324–1338. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.H.; Bandurski, R.S.; Reusch, W.H. Isolation and characterization of a cyclic hydroxamate from Zea mays. Cereal Chem. 1962, 39, 107–113. [Google Scholar]
- Hashimoto, Y.; Shudo, K.; Okamoto, T.; Nagao, M.; Takahashi, Y.; Sugimura, T. Mutagenicities of 4-hydroxy-1,4-benzoxazinones naturally occurring in maize plants and of related compounds. Mutat. Res. 1979, 66, 191–194. [Google Scholar] [CrossRef]
- Arroyo, E.; Chinchilla, N.; Molinillo, J.M.; Macias, F.A.; Astola, A.; Ortiz, M.; Valdivia, M.M. Aneugenic effects of benzoxazinones in cultured human cells. Mutat. Res. 2010, 695, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, C.A.; Nersesyan, A.; Kopp, B.; Schauberger, D.; Darroudi, F.; Grummt, T.; Krupitza, G.; Kundi, M.; Schulte-Hermann, R.; Knasmueller, S. Dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA) and 2,4-dihydroxy-1,4-benzoxazin-3-one (DIBOA), two naturally occurring benzoxazinones contained in sprouts of gramineae are potent aneugens in human-derived liver cells (hepg2). Cancer Lett. 2007, 246, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, S.K.; Pedersen, H.A.; Adhikari, K.B.; Laursen, B.B.; Jensen, C.; Hoyer, S.; Borre, M.; Pedersen, H.H.; Borre, M.; Edwards, D.; et al. Benzoxazinoids in prostate cancer patients after a rye-intensive diet: Methods and initial results. J. Agric. Food Chem. 2016, 64, 8235–8245. [Google Scholar] [CrossRef] [PubMed]
- Whitney, N.J.; Mortimore, C.G. Effect of 6-methoxybenzoxazolinone on the growth of Xanthomonas stewartii (erw. Smith) dowson and its presence in sweet corn (Zea mays var. Saccharata bailey). Nature 1961, 189, 596. [Google Scholar] [CrossRef]
- Corcuera, L.J.; Woodward, M.D.; Helgeson, J.P.; Kelman, A.; Upper, C.D. 2,4-dihydroxy-7-methoxy-2h-1,4-benzoxazin-3(4h)-one, an inhibitor from Zea mays with differential activity against soft rotting erwinia species. Plant Physiol. 1978, 61, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Sahi, S.V.; Chilton, M.D.; Chilton, W.S. Corn metabolites affect growth and virulence of Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA 1990, 87, 3879–3883. [Google Scholar] [CrossRef] [PubMed]
- Rasmann, S.; Turlings, T.C. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Neal, A.L.; Ahmad, S.; Gordon-Weeks, R.; Ton, J. Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS ONE 2012, 7, e35498. [Google Scholar] [CrossRef] [PubMed]
- Neal, A.L.; Ton, J. Systemic defense priming by Pseudomonas putida KT2440 in maize depends on benzoxazinoid exudation from the roots. Plant Signal. Behav. 2013, 8, e22655. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, A.I.; Hietala, P.K. 2(3)-benzoxazolinone, an anti-Fusarium factor in rye seedlings. Acta Chem. Scand. 1955, 9, 1543–1544. [Google Scholar] [CrossRef]
- Glenn, A.E.; Gold, S.E.; Bacon, C.W. Fdb1 and fdb2, Fusarium verticillioides loci necessary for detoxification of preformed antimicrobials from corn. Mol. Plant Microbe Interact. 2002, 15, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Etzerodt, T.; Maeda, K.; Nakajima, Y.; Laursen, B.; Fomsgaard, I.S.; Kimura, M. 2,4-dihydroxy-7-methoxy-2h-1,4-benzoxazin-3(4h)-one (DIMBOA) inhibits trichothecene production by Fusarium graminearum through suppression of tri6 expression. Int. J. Food Microbiol. 2015, 214, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.; Kohn, L.M. Host-synthesized secondary compounds influence the in vitro interactions between fungal endophytes of maize. Appl. Environ. Microbiol. 2008, 74, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Glenn, A.E.; Bacon, C.W. Fdb2 encodes a member of the arylamine n-acetyltransferase family and is necessary for biotransformation of benzoxazolinones by Fusarium verticillioides. J. Appl. Microbiol. 2009, 107, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Glenn, A.E.; Hinton, D.M.; Yates, I.E.; Bacon, C.W. Detoxification of corn antimicrobial compounds as the basis for isolating Fusarium verticillioides and some other Fusarium species from corn. Appl. Environ. Microbiol. 2001, 67, 2973–2981. [Google Scholar] [CrossRef] [PubMed]
- Ridenour, J.B.; Bluhm, B.H. The novel fungal-specific gene fug1 has a role in pathogenicity and fumonisin biosynthesis in Fusarium verticillioides. Mol. Plant Pathol. 2017, 18, 513–528. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.; Kohn, L.M. Evidence for alteration of fungal endophyte community assembly by host defense compounds. New Phytol. 2009, 182, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Bravo, H.R.; Lazo, W. Antialgal and antifungal activity of natural hydroxamic acids and related compounds. J. Agricl. Food Chem. 1996, 44, 1569–1571. [Google Scholar] [CrossRef]
- Bravo, H.R.; Copaja, S.V.; Lazo, W. Antimicrobial activity of natural 2-benzoxazolinones and related derivatives. J. Agricl. Food Chem. 1997, 45, 3255–3257. [Google Scholar] [CrossRef]
- Glensk, M.; Gajda, B.; Franiczek, R.; Krzyzanowska, B.; Biskup, I.; Wlodarczyk, M. In vitro evaluation of the antioxidant and antimicrobial activity of DIMBOA [2,4-dihydroxy-7-methoxy-2h-1,4-benzoxazin-3(4h)-one]. Nat. Prod. Res. 2016, 30, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Veyrat, N.; Gordon-Weeks, R.; Zhang, Y.; Martin, J.; Smart, L.; Glauser, G.; Erb, M.; Flors, V.; Frey, M.; et al. Benzoxazinoid metabolites regulate innate immunity against aphids and fungi in maize. Plant Physiol. 2011, 157, 317–327. [Google Scholar] [CrossRef] [PubMed]
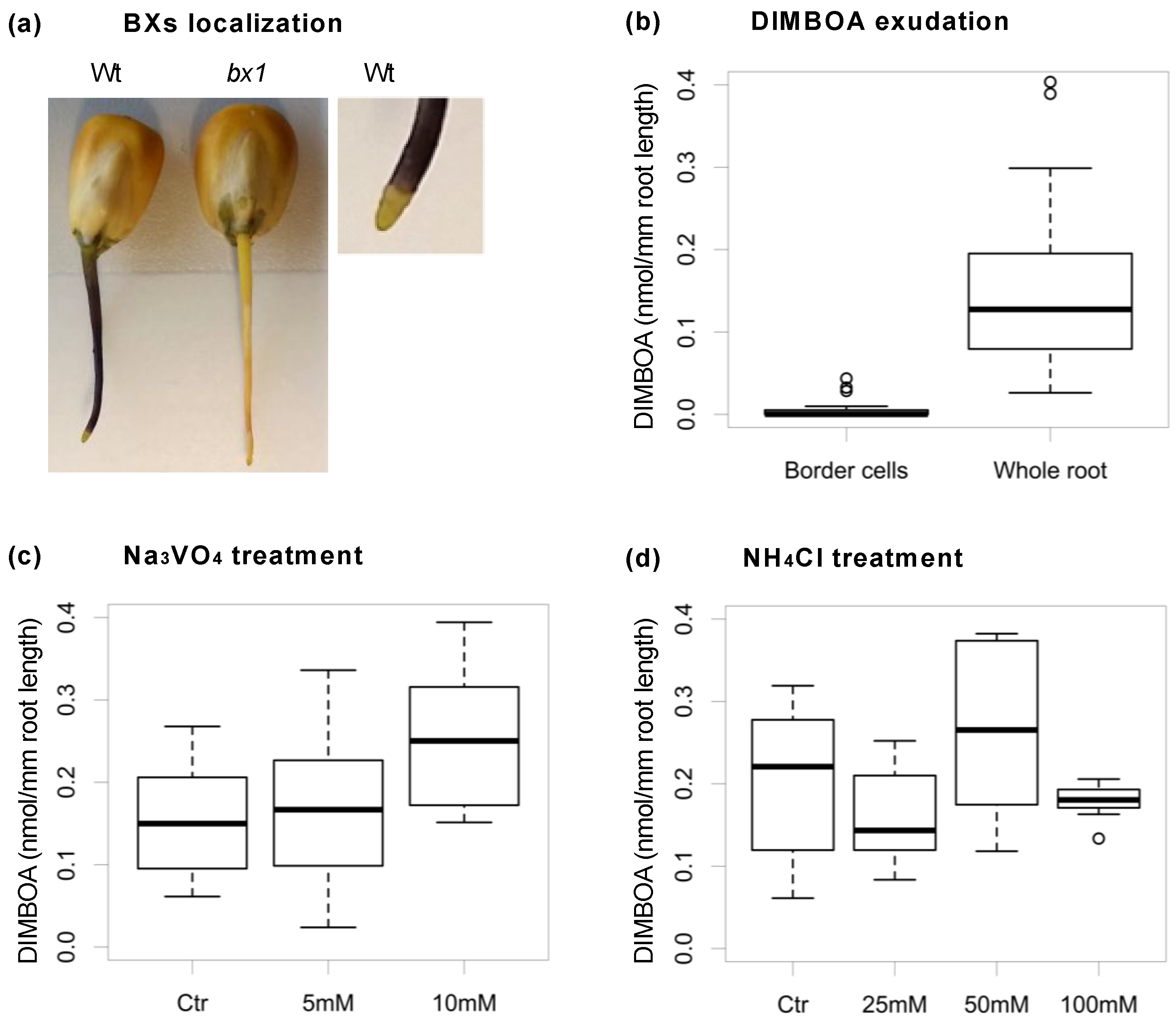
- Niculaes, C.; Frey, M. (Technical University of Munich, Freising, Germany) Application of transport inhibitors: Surface sterilized seeds were left overnight in tubes containing 50 mL of the inhibitor solution in water for imbibition. The bottom of plastic containers was covered with sponges and 50 mL of the inhibitor solution added. The sponges were pricked with straight steel pins. On each pin an imbibed seed was mounted. Seedlings were grown in the closed containers in the dark at 28 °C for 3 days. Collection of the exudates was done by washing the roots with 100 l water. Inhibitors applied were sodium orthovanadate, potassium cyanide, sodium azide, gramicidin, ammonium chloride, verapamil, glibenclamide and nifedipine. All inhibitors were purchased from Sigma-Aldrich Germany, now Merck KGaA, Darmstadt, Germany. The maize inbred line B73 was used in the experiment. 2016
- Cannesan, M.A.; Gangneux, C.; Lanoue, A.; Giron, D.; Laval, K.; Hawes, M.; Driouich, A.; Vicré-Gibouin, M. Association between border cell responses and localized root infection by pathogenic Aphanomyces euteiches. Ann. Bot. 2011, 108, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.A.; Veyrat, N.; Glauser, G.; Marti, G.; Doyen, G.R.; Villard, N.; Gaillard, M.D.; Kollner, T.G.; Giron, D.; Body, M.; et al. A specialist root herbivore exploits defensive metabolites to locate nutritious tissues. Ecol. Lett. 2012, 15, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Frey, M. (Technical University of Munich, Freising, Germany). The maize inbred line B73 was crossed with the bx1 mutant line. The bx1 homozygous mutant was selected in the F2 progeny. After six backcross generations the homozygous bx1 mutant line was selected and propagated by selfing. 2018.
- Putnam, A.R.; DeFrank, J. Use of phytotoxic plant residues for selective weed control. Crop Prot. 1983, 2, 173–181. [Google Scholar] [CrossRef]
- Belz, R.G.; Hurle, K. Differential exudation of two benzoxazinoids-one of the determining factors for seedling allelopathy of triticeae species. J. Agric. Food Chem. 2005, 53, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Molinillo, J.M.G. Optimization of benzoxazinones as natural herbicide models by lipophilicity enhancement. J. Agric. Food Chem. 2006, 54, 9357–9365. [Google Scholar] [CrossRef] [PubMed]
- Fomsgaard, I.S.; Spliid, N.H.; Felding, G. Leaching of pesticides through normal-tillage and low-tillage soil--a lysimeter study. I. Isoproturon. J. Environ. Sci. Heal. B 2003, 38, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Oliveros-Bastidas, A.; Marin, D.; Castellano, D.; Simonet, A.M.; Molinillo, J.M. Degradation Studies on Benzoxazinoids. Soil Degradation Dynamics of 2,4-Dihydroxy-7-methoxy-(2H)-1,4-benzoxazin-3(4H)-one (DIMBOA) and Its Degradation Products, Phytotoxic Allelochemicals from Gramineae. J. Agric. Food Chem. 2004, 52, 6402–6413. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Oliveros-Bastidas, A.; Marín, D.; Chinchilla, N.; Castellano, D.; Molinillo, J.M.G. Evidence for an allelopathic interaction between rye and wild oats. J. Agric. Food Chem. 2014, 62, 9450–9457. [Google Scholar] [CrossRef] [PubMed]
- Sahi, S.V.; Anderson, C.E.; Chilton, W.S. The corn wound metabolite DIMBOA causes cell death in tobacco and corn. Plant Sci. 1995, 108, 31–40. [Google Scholar] [CrossRef]
- Baerson, S.R.; Sánchez-Moreiras, A.; Pedrol-Bonjoch, N.; Schulz, M.; Kagan, I.A.; Agarwal, A.K.; Reigosa, M.J.; Duke, S.O. Detoxification and transcriptome response in Arabidopsis seedlings exposed to the allelochemical benzoxazolin-2(3h)-one. J. Agric. Food Chem. 2005, 280, 21867–21881. [Google Scholar] [CrossRef]
- Hofmann, D.; Knop, M.; Hao, H.; Hennig, L.; Sicker, D.; Schulz, M. Glucosides from MBOA and BOA detoxification by Zea mays and Portulaca oleracea. J. Nat. Prod. 2006, 69, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Schulz, M.; Filary, B.; Kuhn, S.; Colby, T.; Harzen, A.; Schmidt, J.; Sicker, D.; Hennig, L.; Hofmann, D.; Disko, U.; et al. Benzoxazolinone detoxification by N-glucosylation: The multi-compartment-network of Zea mays L. Plant Signal. Behav. 2016, 11, e1119962. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Venturelli, S.; Belz, R.G.; Kamper, A.; Berger, A.; von Horn, K.; Wegner, A.; Bocker, A.; Zabulon, G.; Langenecker, T.; Kohlbacher, O.; et al. Plants release precursors of histone deacetylase inhibitors to suppress growth of competitors. Plant Cell 2015, 27, 3175–3189. [Google Scholar] [CrossRef] [PubMed]
- Petho, M. Possible role of cyclic hydroxamic acids in the iron uptake by grasses. Acta Agron. Hung. 1993, 42, 203–214. [Google Scholar]
- Petho, M. Physiological role of the cyclic hydroxamic acids. In Proceedings of the 7th Hungarian Congress on Plant Physiology, Szeged, Hungary, 24–27.06.2002; Petho M., Ed. Acta Biol.(Szeged) 2002, 46, 175–176. [Google Scholar]
- Poschenrieder, C.; Tolra, R.P.; Barcelo, J. A role for cyclic hydroxamates in aluminium resistance in maize? J. Inorg. Biochem. 2005, 99, 1830–1836. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.T.; Famoso, A.N.; Zhao, K.; Shaff, J.E.; Craft, E.J.; Bustamante, C.D.; McCouch, S.R.; Aneshansley, D.J.; Kochian, L.V. High-throughput two-dimensional root system phenotyping platform facilitates genetic analysis of root growth and development. Plant Cell Environ. 2013, 36, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, C.T.; Simoes, C.C.; Pastina, M.M.; Maron, L.G.; Magalhaes, J.V.; Vasconcellos, R.C.; Guimaraes, L.J.; Lana, U.G.; Tinoco, C.F.; Noda, R.W.; et al. Genetic dissection of al tolerance qtls in the maize genome by high density snp scan. BMC Genom. 2014, 15, 153. [Google Scholar] [CrossRef] [PubMed]
- Klun, J.A.; Tipton, C.L.; Brindley, T.A. 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA), an active agent in the resistance of maize to the European corn borer123. J. Econ. Entomol. 1967, 60, 1529–1533. [Google Scholar] [CrossRef]
- Köhler, A.; Maag, D.; Veyrat, N.; Glauser, G.; Wolfender, J.L.; Turlings, T.C.J.; Erb, M. Within-plant distribution of 1,4-benzoxazin-3-ones contributes to herbivore niche differentiation in maize. Plant Cell Environ. 2015, 38, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Barry, D.; Alfaro, D.; Darrah, L.L. Relation of European corn borer (Lepidoptera: Pyralidae) leaf-feeding resistance and DIMBOA content in maize. Environ. Entomol. 1994, 23, 177–182. [Google Scholar] [CrossRef]
- Campos, F.; Atkinson, J.; Arnason, J.T.; Philogéne, B.J.R.; Morand, P.; Werstiuk, N.H.; Timmins, G. Toxicity and toxicokinetics of 6-methoxybenzoxazolinone (MBOA) in the european corn borer, Ostrinia nubilalis (Hübner). J. Chem. Ecol. 1988, 14, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.; Atkinson, J.; Arnason, J.T.; Philogène, B.J.R.; Morand, P.; Werstiuk, N.H.; Timmins, G. Toxicokinetics of 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA) in the european corn borer, Ostrinia nubilalis (Hübner). J. Chem. Ecol. 1989, 15, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Maag, D.; Dalvit, C.; Thevenet, D.; Köhler, A.; Wouters, F.C.; Vassão, D.G.; Gershenzon, J.; Wolfender, J.-L.; Turlings, T.C.J.; Erb, M.; et al. 3-β-d-glucopyranosyl-6-methoxy-2-benzoxazolinone (MBOA-N-glc) is an insect detoxification product of maize 1,4-benzoxazin-3-ones. Phytochemistry 2014, 102, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kojima, W.; Fujii, T.; Suwa, M.; Miyazawa, M.; Ishikawa, Y. Physiological adaptation of the asian corn borer Ostrinia furnacalis to chemical defenses of its host plant, maize. J. Insect Physiol. 2010, 56, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Liang, X.; Zhu, X. The role of DIMBOA on the feeding of Asian corn borer, Ostrinia furnacalis (guenée) (Lep., Pyralidae). J. Appl. Entomol. 1999, 123, 49–53. [Google Scholar] [CrossRef]
- Glauser, G.; Marti, G.; Villard, N.; Doyen, G.A.; Wolfender, J.L.; Turlings, T.C.J.; Erb, M. Induction and detoxification of maize 1,4-benzoxazin-3-ones by insect herbivores. Plant J. 2011, 68, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Rostás, M. The effects of 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one on two species of Spodoptera and the growth of Setosphaeria turcica in vitro. J. Pest Sci. 2006, 80, 35–41. [Google Scholar] [CrossRef]
- Manuwoto, S.; Mark Scriber, J. Consumption and utilization of three maize genotypes by the southern armyworm. J. Econo. Entomol. 1982, 75, 163–167. [Google Scholar] [CrossRef]
- Ortego, F.; Castanera, P.; Ruiz, M. Effect of DIMBOA on growth and digestive physiology of Sesamia nonagrioides (Lepidoptera: Noctuidae) larvae. J. Insect Physiol. 1998, 44, 95–101. [Google Scholar] [CrossRef]
- Hedin, P.A.; Davis, F.M.; Williams, W.P. 2-hydroxy-4,7-dimethoxy-1,4-benzoxazin-3-one (N-O-Me-DIMBOA), a possible toxic factor in corn to the southwestern corn borer. J. Chem. Ecol. 1993, 19, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Alouw, J.C.; Miller, N.J. Effects of benzoxazinoids on specialist and generalist Diabrotica species. J. Appl. Entomol. 2015, 139, 424–431. [Google Scholar] [CrossRef]
- Bjostad, L.B.; Hibbard, B.E. 6-methoxy-2-benzoxazolinone: A semiochemical for host location by western corn rootworm larvae. J. Chem. Ecol. 1992, 18, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.S.; Arnason, J.T.; Philogène, B.J.R.; Lambert, J.D.H.; Atkinson, J.; Morand, P. Role of 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA) in the resistance of maize to western corn rootworm, Diabrotica virgifera virgifera (Leconte) (Coleoptera: Chrysomelidae). Can. Entomol. 1990, 122, 1177–1186. [Google Scholar] [CrossRef]
- Rogers, C.D.; Evans, K.A. Wheat bulb fly (Delia coarctata, fallen, Diptera: Anthomyiidae) larval response to hydroxamic acid constituents of host-plant root exudates. Bull. Entomol. Res. 2013, 103, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Argandoña, V.H.; Luza, J.G.; Niemeyer, H.M.; Corcuera, L.J. Role of hydroxamic acids in the resistance of cereals to aphids. Phytochemistry 1980, 19, 1665–1668. [Google Scholar] [CrossRef]
- Cambier, V.; Hance, T.; De Hoffmann, E. Effects of 1,4-benzoxazin-3-one derivatives from maize on survival and fecundity of Metopolophium dirhodum (Walker) on artificial diet. J. Chem. Ecol. 2001, 27, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.M. Effect of 6-methoxybenzoxazolin-2-one (MBOA) on the reproduction rate of the grain aphid (Sitobion avenae F.). J. Agric. Food Chem. 2006, 54, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Argandoña, V.H.; Niemeyer, H.M.; Corcuera, L.J. Effect of content and distribution of hydroxamic acids in wheat on infestation by the aphid Schizaphis graminum. Phytochemistry 1981, 20, 673–676. [Google Scholar] [CrossRef]
- Corcuera, L.J.; Argandoña, V.H.; Niemeyer, H.M. Effect of cyclic hydroxamic acids from cereals on aphids. In Proceedings of the Chemistry and Biology of Hydroxamic Acids—1st International Symposium, Dayton, Ohio, USA, May 1981; Kehl, H., Ed.; Karger: Basel, Switzerland, 1982; pp. 111–118. [Google Scholar]
- Bohidar, K.; Wratten, S.D.; Niemeyer, H.M. Effects of hydroxamic acids on the resistance of wheat to the aphid Sitobion avenae. Ann. Appl. Biol. 1986, 109, 193–198. [Google Scholar] [CrossRef]
- Leszczynski, B.; Dixon, A.F.G. Resistance of cereals to aphids: The interaction between hydroxamic acids and glutathione s-transferases in the grain aphid Sitobion avenae (F.) (Hom., Aphididae). J. Appl. Entomol. 1992, 113, 61–67. [Google Scholar] [CrossRef]
- Leszczynski, B.; Matok, H.; Dixon, A.F.G. Resistance of cereals to aphids: The interaction between hydroxamic acids and UDP-glucose transferases in the aphid Sitobion avenue (Homoptera: Aphididae). J. Chem. Ecol. 1992, 18, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, H.M.; Pesel, E.; Franke, S.; Francke, W. Ingestion of the benzoxazinone DIMBOA from wheat plants by aphids. Phytochemistry 1989, 28, 2307–2310. [Google Scholar] [CrossRef]
- Betsiashvili, M.; Ahern, K.R.; Jander, G. Additive effects of two quantitative trait loci that confer Rhopalosiphum maidis (corn leaf aphid) resistance in maize inbred line Mo17. J. Exp. Bot. 2015, 66, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Bing, J.W.; Guthrie, W.D.; Dicke, F.F.; Obryckp, J.J. Relation of corn leaf aphid (Homoptera: Aphididae) colonization to DIMBOA content in maize inbred lines. J. Econ. Entomol. 1990, 83, 1626–1632. [Google Scholar] [CrossRef]
- Couture, R.M.; Routley, D.G.; Dunn, G.M. Role of cyclic hydroxamic acids in monogenic resistance of maize to Helminthosporium turcicum. Physiol. Plant Pathol. 1971, 1, 515–521. [Google Scholar] [CrossRef]
- Long, B.J.; Dunn, G.M.; Routley, D.G. Relationship of hydroxamate concentration in maize and field reaction to Helminthosporium turcicum. Crop Sci. 1978, 18, 573–575. [Google Scholar] [CrossRef]
- Kostandi, S.F.; Koraiem, Y.S.; Kamara, A.; Omar, M.A. Effect of phenols in host-pathogen interaction of maize (Zea mays L.). Cephalosporium maydis system. Agrochimica 1981, 367–375. [Google Scholar]
- Elnaghy, M.A.; Shaw, M. Correlation between resistance to stem rust and the concentration of a glucoside in wheat. Nature 1966, 210, 417. [Google Scholar] [CrossRef]
- Zheng, Y.Q.; Zhao, Y.; Dong, F.S.; Yao, J.R.; Hurle, K. Relationship of DIMBOA content in wheat seedlings and its resistance to plant pathogens. Allelopathy J. 2005, 15, 137–143. [Google Scholar]
- Lyons, P.C.; Nicholson, R.L. Evidence that cyclic hydroxamate concentrations are not related to resistance of corn leaves to anthracnose. Can. J. Plant Pathol. 1989, 11, 215–220. [Google Scholar] [CrossRef]
- Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Castellano, D.; Simonet, A.M.; Molinillo, J.M.G. Structure−activity relationships (SAR) studies of benzoxazinones, their degradation products and analogues. Phytotoxicity on standard target species (STS). J. Agric. Food Chem. 2005, 53, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.P.; Putnam, A.R.; Burke, B.A.; Aasen, A.J. Isolation and characterization of allelochemicals in rye herbage. Phytochemistry 1987, 26, 1385–1390. [Google Scholar] [CrossRef]
- Chiapusio, G.; Pellissier, F.; Gallet, C. Uptake and translocation of phytochemical 2-benzoxazolinone (BOA) in radish seeds and seedlings. J. Exp. Bot. 2004, 55, 1587–1592. [Google Scholar] [CrossRef] [PubMed]
- Burgos, N.R.; Talbert, R.E. Differential activity of allelochemicals from Secale cereale in seedling bioassays. Weed Sci. 2000, 48, 302–310. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Setia, N.; Kohli, R.K. Effects of 2-benzoxazolinone on the germination, early growth and morphogenetic response of mung bean (Phaseolus aureus). Ann. Appl. Biol. 2005, 147, 267–274. [Google Scholar] [CrossRef]
- Tabaglio, V.; Gavazzi, C.; Schulz, M.; Marocco, A. Alternative weed control using the allelopathic effect of natural benzoxazinoids from rye mulch. Agron. Sustain. Dev. 2008, 28, 397–401. [Google Scholar] [CrossRef]
- Schulz, M.; Marocco, A.; Tabaglio, V. BOA detoxification of four summer weeds during germination and seedling growth. J. Chem. Ecol. 2012, 38, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.I.; Reigosa, M.J. Allelochemical stress inhibits growth, leaf water relations, PSII photochemistry, non-photochemical fluorescence quenching, and heat energy dissipation in three C3 perennial species. J. Exp. Bot. 2011, 62, 4533–4545. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Castellano, D.; Simonet, A.M.; Molinillo, J.M.G. Structure−activity relationship (SAR) studies of benzoxazinones, their degradation products, and analogues. Phytotoxicity on problematic weeds Avena fatua l. and Lolium rigidum gaud. J. Agric. Food Chem. 2006, 54, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Chinchilla, N.; Varela, R.M.; Oliveros-Bastidas, A.; Marín, D.; Molinillo, J.M.G. Structure−activity relationship studies of benzoxazinones and related compounds. Phytotoxicity on Echinochloa crus-galli (L.) p. Beauv. J. Agric. Food Chem. 2005, 53, 4373–4380. [Google Scholar] [CrossRef]
- Barnes, J.P.; Putnam, A.R. Role of benzoxazinones in allelopathy by rye (Secale cereale l.). J. Chem. Ecol. 1987, 13, 889–906. [Google Scholar] [CrossRef] [PubMed]
- Dafoe, N.J.; Huffaker, A.; Vaughan, M.M.; Duehl, A.J.; Teal, P.E.; Schmelz, E.A. Rapidly induced chemical defenses in maize stems and their effects on short-term growth of ostrinia nubilalis. J. Chem. Ecol. 2011, 37, 984. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Guo, J.; He, K.; Bai, S.; Zhang, T.; Zhao, J.; Wang, Z. Physiological responses induced by Ostrinia furnacalis (Lepidoptera: Crambidae) feeding in maize and their effects on o. Furnacalis performance. J. Econ. Entomol. 2017, 110, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Maag, D.; Kohler, A.; Robert, C.A.; Frey, M.; Wolfender, J.L.; Turlings, T.C.; Glauser, G.; Erb, M. Highly localized and persistent induction of Bx1-dependent herbivore resistance factors in maize. Plant J. 2016, 88, 976–991. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, H.; Zhuang, H.; Zhao, C.; Xu, Y.; Wu, S.; Qi, J.; Li, J.; Hettenhausen, C.; Wu, J. Transcriptomics and alternative splicing analyses reveal large differences between maize lines B73 and Mo17 in response to aphid Rhopalosiphum padi infestation. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Argandoña, V.H.; Corcuera Luis, J.; Niemeyer Hermann, M.; Campbell Bruce, C. Toxicity and feeding deterrency of hydroxamic acids from gramineae in synthetic diets against the greenbug, Schizaphis graminum. Entomol. Exp. Appl. 1983, 34, 134–138. [Google Scholar] [CrossRef]
- Givovich, A.; Morse, S.; Cerda, H.; Niemeyer, H.M.; Wratten, S.D.; Edwards, P.J. Hydroxamic acid glucosides in honeydew of aphids feeding on wheat. J. Chem. Ecol. 1992, 18, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Flint-Garcia, S.A.; Thuillet, A.C.; Yu, J.; Pressoir, G.; Romero, S.M.; Mitchell, S.E.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler, E.S. Maize association population: A high-resolution platform for quantitative trait locus dissection. Plant J. 2005, 44, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Q.; Wu, Y.; Lemmon, Z.H.; Xu, G.; Huang, C.; Liang, Y.; Xu, D.; Li, D.; Doebley, J.F.; et al. Genome-wide analysis of transcriptional variability in a large maize-teosinte population. Mol. Plant 2018, 11, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.A.M.; Zhang, X.; Machado, R.A.R.; Schirmer, S.; Lori, M.; Mateo, P.; Erb, M.; Gershenzon, J. Sequestration and activation of plant toxins protect the western corn rootworm from enemies at multiple trophic levels. eLife 2017, 6, e29307. [Google Scholar] [CrossRef] [PubMed]
- Sasai, H.; Ishida, M.; Murakami, K.; Tadokoro, N.; Ishihara, A.; Nishida, R.; Mori, N. Species-specific glucosylation of DIMBOA in larvae of the rice armyworm. Biosci. Biotechnol. Biochem. 2009, 73, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Clay, N.K.; Adio, A.M.; Denoux, C.; Jander, G.; Ausubel, F.M. Glucosinolate metabolites required for an Arabidopsis innate immune response. Science 2009, 323, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Romero Navarro, J.A.; Willcox, M.; Burgueno, J.; Romay, C.; Swarts, K.; Trachsel, S.; Preciado, E.; Terron, A.; Delgado, H.V.; Vidal, V.; et al. A study of allelic diversity underlying flowering-time adaptation in maize landraces. Nat. Genet. 2017, 49, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.M.; Jepsen, H.S.K.; Halkier, B.A.; Kliebenstein, D.J.; Burow, M. Natural variation in cross-talk between glucosinolates and onset of flowering in Arabidopsis. Front. Plant Sci. 2015, 6, 697. [Google Scholar] [CrossRef] [PubMed]
- Venis, M.A.; Watson, P.J. Naturally occuring modifiers of auxin-receptor interaction in corn: Identification as benzoxazolinones. Planta 1978, 142, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Togo, S.; Urashima, M.; Mizutani, J.; Kosemura, S.; Yamamura, S. An auxin-inhibiting substance from light-grown maize shoots. Phytochemistry 1992, 31, 3673–3676. [Google Scholar] [CrossRef]
- Park, W.J.; Schäfer, A.; Prinsen, E.; van Onckelen, H.; Kang, B.G.; Hertel, R. Auxin-induced elongation of short maize coleoptile segments is supported by 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one. Planta 2001, 213, 92–100. [Google Scholar] [CrossRef] [PubMed]
- González, L.F.; Rojas, M.C. Role of wall peroxidases in oat growth inhibition by DIMBOA. Phytochemistry 1999, 50, 931–937. [Google Scholar] [CrossRef]
- Jabeen, R.; Yamada, K.; Shigemori, H.; Hasegawa, T.; Hara, M.; Kuboi, T.; Hasegawa, K. Induction of ß-glucosidase activity in maize coleoptiles by blue light illumination. J. Plant Physiol. 2006, 163, 538–545. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target Organism | Benzoxazinoids | |||
|---|---|---|---|---|
| Core Benzoxazinones | Induced BXs * | BX Metabolites ** | ||
| Chewing insects | ||||
| Leaf-feeders | [59] | induce increase in GHDMBOA | ||
| Generalists | ||||
| Ostrinia nubilalis (Hübner, 1796) | [110,111,112,113] | DIBOA, DIMBOA | DIM2BOA | MBOA |
| Ostrinia furnacalis (Guenée, 1854) | [114,115] | DIMBOA | HDMBOA | MBOA |
| Ostrinia latipennis (Warren, 1892) | [114] | |||
| Ostrinia scapulalis(Walker, 1859) | [115] | |||
| Spodoptera littoralis (Boisduval, 1833) | [113,116] | DIMBOA | HDMBOA deterring | |
| Spodoptera exigua (Hübner 1808) | [117] | DIMBOA | ||
| Spodoptera eridania (Stoll, 1781) | [118] | # | ||
| Sesamia nonagrioides (Lefèbvre, 1827) | [119] | DIMBOA | ||
| Diatraea grandiosella (Dyar, 1911) | [120] | MBOA | ||
| Specialists | ||||
| Spodoptera frugiperda (J. E. Smith, 1797) | [10,113,116] | DIMBOA | HDMBOA deterring | MBOA |
| Rootworms | ||||
| Generalists | ||||
| Diabrotica balteata (LeConte, 1865) | [91] | DIMBOA | DIM2BOA | HMBOA |
| D.undecimpunctata howardi (Barber, 1947) | [121] | # | ||
| Specialists | ||||
| Diabrotica virgifera virgifera (LeConte, 1858) | [122,123] | DIBOA, DIMBOA | DIM2BOA | |
| [91] | ||||
| [121] | ||||
| Delia coarctata (Fallén, 1825) | [124] | DIMBOA | MBOA | |
| Aphids | ||||
| Metopolophium dirhodium (Walker, 1849) | [125,126,127] | DIMBOA | GHDMBOA | MBOA |
| Schizaphis graminum (Rondani, 1852) | [125,128,129] | DIMBOA | ||
| Sitobion avena (Fabricius, 1775) | [130,131,132,133] | # | ||
| Rhopalosiphum padi (Linnaeus, 1758) | [88] | # | ||
| Rhopalosiphum maidis (Fitch, 1856) | [134,135] | DIMBOA | GDMHBOA | ? |
| Fungi | [58] | induce increase in GHDMBOA | ||
| Helminthosporium turcicum Pass. | [136,137] | DIMBOA | ||
| Cephalosporium maydis Samra, Sabet & Hing. | [138] | |||
| Puccinia graminis Pers. | [139,140] | |||
| Bipolaris maydis (Y. Nisik. & C. Miyake) Shoemaker | [43] | MBOA | ||
| Curvularia lunata R.R. Nelson & Haasis | [43] | MBOA | ||
| Alternaria alternata (Fr.) Keissl. (Fr.) | [43] | MBOA | ||
| Colletotrichum graminicola D.J. Politis | [141] | |||
| Ustilago zeae (Link) Unger | [23] | |||
| Fusarium verticillioides (Sacc.) Nirenberg | [23] | increased fungal toxin production | ||
| Gibberella zeae (Schwein.) Petch | [83] | reduced fungal toxin production | ||
| Plants | ||||
| Dicots | ||||
| Lepidium sativum L. (Brassicales) **** | [142] | DIBOA, DIMBOA | APO | |
| Lepidium sativum L. (Brassicales) **** | [143] | DIBOA | BOA | |
| Raphanus sativus L. (Brassicales) **** | [144] | BOA | ||
| Solanum lycopersicum L. (Solanales) **** | [142] | DIBOA, DIMBOA | BOA, MBOA, APO | |
| Cucumis melo L. (Cucurbitales) **** | [145] | DIBOA | BOA | |
| Cucumis sativus L. (Cucurbitales) **** | [145] | ## | ||
| Cucurbita pepo L.(Cucurbitales) **** | [145] | ## | ||
| Lactuca sativa L. (Asterales) **** | [145] | DIBOA | BOA | |
| Vicia faba L. (Fabales) **** | [21] | DIBOA | ||
| Vigna radiata (L.) R.Wilczek (Fabales) **** | [146] | BOA | ||
| Portulaca oleraceae L.(Caryophylales) | [147] | ## | ||
| Arabidopsis thaliana (L.) Heyhn (Brassicales) | [31] | DIBOA, DIMBOA | ||
| Abutilon theophrasti Medik. (Malvales) | [145] | ## | ||
| Amaranthus palmeri S. Watson (Caryophyllales) | [145] | DIBOA | BOA | |
| Amaranthus retroflexus L. (Caryophyllales | [145] | ## | ||
| Chenopodium album L. (Caryophyllales) | [93,148] | ## | BOA | |
| Ipomoea hederacea Jacq. (Solanales) | [145] | ## | ||
| Ipomoea lacunosa L. (Solanales) | [145] | ## | ||
| Sesbania exaltata (Raf.) Rydb. (Fabales) | [145] | ## | ||
| Senna obtusifolia (L.) H.S.Irwin & Barneby (Fabales) | [145] | ## | ||
| Rumex acetosa L. (Rumex acetosa) | [149] | BOA | ||
| Sida spinosa L. (Malvales) | [145] | ## | ||
| Monocots | ||||
| Allium cepa L. (Aspargales) **** | [142] | DIBOA, DIMBOA | MBOA, APO | |
| Zea mays L. (Poales) **** | [145] | DIBOA | BOA | |
| Triticum aestivum L. (Poales) **** | [142] | DIBOA, DIMBOA | APO | |
| Avena sativa L. (Poales) **** | [21] | DIMBOA | ||
| Avena fatua L. (Poales) | [95,150] | DIBOA, DIMBOA | BOA, MBOA, APO | |
| Lolium rigidum Gaudin (Poales) | [95,150] | DIBOA, DIMBOA | BOA, MBOA, APO | |
| Lolium perenne L. (Poales) | [95,150] | BOA | ||
| Dactylis glomerata L. (Poales) | [149] | BOA | ||
| Digitaria sanguinalis (L.) Scop. (Poales) | [145] | ## | ||
| Echinochloa crus-galli (L.) P.Beauv. (Poales) | [143,151] | DIBOA, DIMBOA | APO, BOA | |
| Eleusine indica (L.) Gaertn. (Poales) | [145] | DIBOA | BOA | |
| Panicum miliaceum L. (Poales) | [152] | DIBOA | BOA | |

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niculaes, C.; Abramov, A.; Hannemann, L.; Frey, M. Plant Protection by Benzoxazinoids—Recent Insights into Biosynthesis and Function. Agronomy 2018, 8, 143. https://doi.org/10.3390/agronomy8080143
Niculaes C, Abramov A, Hannemann L, Frey M. Plant Protection by Benzoxazinoids—Recent Insights into Biosynthesis and Function. Agronomy. 2018; 8(8):143. https://doi.org/10.3390/agronomy8080143
Chicago/Turabian StyleNiculaes, Claudiu, Aleksej Abramov, Laura Hannemann, and Monika Frey. 2018. "Plant Protection by Benzoxazinoids—Recent Insights into Biosynthesis and Function" Agronomy 8, no. 8: 143. https://doi.org/10.3390/agronomy8080143
APA StyleNiculaes, C., Abramov, A., Hannemann, L., & Frey, M. (2018). Plant Protection by Benzoxazinoids—Recent Insights into Biosynthesis and Function. Agronomy, 8(8), 143. https://doi.org/10.3390/agronomy8080143