Pattern Recognition Receptors—Versatile Genetic Tools for Engineering Broad-Spectrum Disease Resistance in Crops


School of Life Sciences, Phytopathology, Technical University of Munich, Emil-Ramann-Str. 2, 85354 Freising-Weihenstephan, Germany
Agronomy 2018, 8(8), 134; https://doi.org/10.3390/agronomy8080134
Submission received: 1 July 2018
/
Revised: 28 July 2018
/
Accepted: 30 July 2018
/
Published: 1 August 2018
(This article belongs to the Special Issue Biotic and Abiotic Stress Responses in Crop Plants)
{kind=link}
{kind=link}
Abstract
:Infestations of crop plants with pathogens pose a major threat to global food supply. Exploiting plant defense mechanisms to produce disease-resistant crop varieties is an important strategy to control plant diseases in modern plant breeding and can greatly reduce the application of agrochemicals. The discovery of different types of immune receptors and a detailed understanding of their activation and regulation mechanisms in the last decades has paved the way for the deployment of these central plant immune components for genetic plant disease management. This review will focus on a particular class of immune sensors, termed pattern recognition receptors (PRRs), that activate a defense program termed pattern-triggered immunity (PTI) and outline their potential to provide broad-spectrum and potentially durable disease resistance in various crop species—simply by providing plants with enhanced capacities to detect invaders and to rapidly launch their natural defense program.
1. Introduction
Pathogens and pests of plants are a major problem in agricultural food production despite the application of plant protection chemicals [1]. Pre- and post-harvest diseases can cause significant losses in crop yield and impair crop quality. The emergence and global spreading of novel pathogens or pathogen races/strains capable of defeating existing, resistant crop cultivars, such as the wheat stem rust (Puccinia graminis f. sp. tritici) race Ug99 [2] or the kiwi fruit pathogen Pseudomonas syringae pv. actinidae [3], and the increasing resistance of many pathogen races/strains against available pesticides, illustrates the vulnerability of our current plant protection strategies and the looming risk of devastating disease outbreaks. Increasing the yield of high-quality plant products on the available arable farm land while reducing the amount of ecologically harmful agrochemicals, necessitates the development of future-oriented, sustainable agricultural production systems and effective but environment-friendly plant protection measures. In the last 20 years, the identification of a range of different molecular plant immune components has not only greatly advanced our mechanistic understanding of the plant immune system but also provided the conceptual framework to deploy these discoveries for genetic plant protection. Using selected examples, I will demonstrate in which ways PRRs and PTI can be deployed for disease resistance management in crop plants and illustrate future perspectives for molecular engineering of PRRs and PRR signaling.
2. Pattern-Triggered Immunity Forms a Robust Host Barrier to Invaders
Plants rely on genetically determined (innate) immunity to protect themselves from potentially harmful invaders such as pathogens, pests, and parasitic plants. Central to the plant immune system is a multi-layered surveillance system of various extra- and intracellular immune receptors that sense molecular features of the invader (non-self recognition) as well as perturbations of the cellular integrity provoked by the invader (altered-self recognition). Detection of such danger signals by plant immune sensors activates local and systemic defense mechanisms [4,5]. The capacity of a plant to detect invasion attempts early and to mount a defense response in time, i.e., before establishment and proliferation of the invader, largely depends on its repertoire of immune sensors capable of recognizing the invader.
2.1. Microbe-Associated Molecular Patterns (MAMPs)
Plants sense a variety of conserved microbial components with vital roles for microbial fitness as immune elicitors [4]. Because of their important functions, these components are usually conserved across microbial species and cannot readily be modulated to evade recognition without a fitness cost. Such immunogenic molecular structures are termed microbe- or pathogen-associated molecular patterns (MAMPs/PAMPs). MAMPs are chemically diverse, e.g. proteins, polysaccharides, lipids or composite molecules. Classical examples of MAMPs are microbial cell wall structures, such as chitin (fungi), beta-glucans (oomycetes), lipopolysaccharide or peptidoglycan (bacteria), or microbial proteins, such as bacterial flagellin or elongation factor thermo-unstable (EF-Tu, part of the cellular protein translation machinery) [6]. In case of larger molecular structures, a defined partial structure (epitope or pattern) of these molecules is generally sufficient to activate plant immunity. One of the best-studied examples is the flg22 epitope, which corresponds to a stretch of 22 amino acids from the highly conserved N-terminus of flagellin, a region which is important for assembly of the flagellin monomers into of a functional flagellum and for bacterial motility. A synthetic flg22 peptide is sufficient to activate the same defense program as the natural flagellin protein [7]. Similarly, the peptide elf18, which corresponds to the conserved N-terminus of EF-Tu, is sufficient to trigger plant immunity [8]. The use of synthetic peptide MAMPs has greatly expedited functional studies of these proteinaceous elicitors.
The isolation and identification of MAMPs is often a challenging task. While it is relatively easy to enrich fractions of microbial extracts that trigger PTI-like defense responses in plants, the identification of the causal molecular motif is demanding and requires sophisticated biochemical and analytical skills [6,9]. This is largely due to the fact, that MAMPs are not necessarily very abundant microbial components. To detect invaders timely, host plants evolved highly sensitive immune receptors that detect MAMPs at very low concentrations (nanomolar range) [9]. Thus, although novel analytical techniques have expedited the identification of MAMPs, these are usually not routine methods applicable for high-throughput identification of MAMPs from microbial populations.
Studies on proteinaceous MAMPs are at the forefront of research because they can be produced in large amounts and high purity through recombinant expression systems or chemical synthesis, can be easily genetically modified, and are amenable to population-wide genetic studies. Once a protein motif has been identified as MAMP epitope in a microbial species/race/strain, the DNA sequence of the respective gene can be easily analyzed in silico in many species. Such bioinformatic studies in combination with functional assays have revealed that MAMP epitopes are not as strictly conserved as was initially assumed. Although the protein sequences of MAMPs are overall conserved to maintain their function, the MAMP epitopes were found to be more diversified, presumably due to the selective pressure exerted on microbes to evade host immune sensing through these epitopes [10]. flg22 epitopes from different bacterial species, for example, are sensed with different efficacy in different plant species and some flg22 motifs are not detected at all in some plant species [7,11,12,13,14,15]. The specific genetic fingerprints resulting from the opposing (purifying versus diversifying) selection pressure on MAMPs can, in turn, be exploited to identify putative MAMP epitopes de novo by means of bioinformatic genome surveys of microbial populations [16]. In a proof-of-concept study several candidate MAMP epitopes identified by screening P. syringae and Xanthomonas campestris genome data were shown to elicit PTI in Arabidopsis. Hence, with the increasing availability of microbial genome data such in silico screenings will greatly speed up identification of proteinaceous MAMPs in the future and facilitate targeted screenings for MAMP epitopes in pathogen species of interest.
To date, numerous MAMPs from all microbial classes that are sensed by different plant species have been identified and the list is continuously growing (for a comprehensive summary see recent reviews [6,17,18]). In addition, immunogenic molecular patterns have also been enriched from nematodes, insects, and parasitic plants [6,18,19]. Taken together, this demonstrates the central role of MAMP sensing for plant defense against different types of invaders.
2.2. Damage-Associated Molecular Patterns (DAMPs)
Invaders are not only sensed by the plant immune system via their own molecular components (exogeneous elicitors) but usually also provoke the release of plant-derived signals characteristic of infection, called endogenous elicitors or damage-associated molecular patterns (DAMPs) [4]. Typical examples of DAMPs are fragments of cell wall components generated during attack by microbial cell wall-degrading enzymes, e.g., oligogalacturonides (OGs, derived from pectin) [20] or cutin monomers [21], and intracellular plant components released into the extracellular space upon cell lysis, such as extracellular ATP (eATP) [22], extracellular NAD (eNAD) [23], or intracellular proteins (e.g., Arabidopsis HMGB3) [24]. Like MAMPs, DAMPs are sensed by cell surface-resident PRRs and activate typical PTI signaling and defense responses [4].
In addition, plants release various endogenous peptide hormones into the apoplast upon wounding or pathogen attack. Because these peptides trigger typical PTI defense responses via PRR-like receptors, they are also classified as DAMPs. The systemin peptide, for instance, is produced upon wounding in tomato and contributes to plant defense against herbivorous insects [25]. Two peptide families from Arabidopsis, termed AtPEPs and AtPIPs, that are produced upon biotic stress and wounding, apparently play an important role in the amplification of PTI [26,27,28]. Peptides of the PEP family are found in various monocot and dicot plant species [29,30], suggesting that amplification of PTI by endogenous peptide hormones is a common strategy in plant immunity.
2.3. Pattern Recognition Receptors (PRRs)
MAMPs and DAMPs are detected by immune receptors localized at the host cell surface, called PRRs (Figure 1) [4]. PRRs are typically single-span transmembrane or membrane-anchored proteins with structurally diverse extracellular domains, such as leucine-rich repeat (LRR), Lysin-motif (LysM) or lectin domains, that bind MAMP/DAMP epitopes with high specificity and sensitivity [18]. Receptor-like kinase (RLK)-type PRRs possess a cytosolic kinase domain, which usually functions in intracellular signal transduction (Figure 1). Receptor-like protein (RLP)-type PRRs that lack active signaling domains and RLK-type PRRs with non-functional kinase domains require signaling-competent protein partners [18,31]. In general, however, PRRs (RLP-type as well as RLK-type) do not function alone but are part of multi-protein complexes where they engage with co-receptors, signaling partners and regulatory proteins that fine-tune PRR activation/deactivation for appropriate signaling output (Figure 1) [31]. Among the best-studied PRRs to date are the flg22 receptor FLS2 and the elf18 receptor EFR, both of which are LRR-type RLKs [8,32]. Many other LRR-type RLKs and RLPs also detect proteinaceous MAMPs, e.g., RLP23 senses the nlp20 epitope of necrosis- and ethylene-inducing peptide 1-like proteins (NLPs) which show a remarkably broad distribution in bacteria, fungi as well as oomycetes [33,34]. Whereas binding of a MAMP or DAMP to its respective PRR is highly specific, other receptor complex components are commonly shared by several PRRs (Figure 1) [31]. Various LRR-type PRRs (e.g., FLS2 and EFR) interact with the LRR-RLK BAK1 in a ligand-dependent manner [35]. LRR-RLP-type PRRs constitutively associate with the signaling adapter LRR-RLK SOBIR and also recruit BAK1 upon ligand binding [36]. The LysM-RLK CERK1 similarly interacts with different LysM-type PRRs [31].
Individual plant species apparently harbor numerous PRRs that sense different classes of invaders in a partially redundant manner. In this way, each invader is sensed by several PRRs. Arabidopsis, for instance, senses at least seven MAMPs from Pseudomonas bacteria through distinct PRRs [37]. Presumably, the diversity and functional redundancy of PRRs is central to the remarkable robustness of the PTI system. Some PRRs are broadly distributed across the plant kingdom (evolutionary old, e.g., FLS2) whereas others are specific to individual plant families or species (evolutionary young, e.g., EFR) [4]. As a consequence, different plant species have different but partially overlapping sets of PRRs. Just as invaders continuously evolve to evade MAMP sensing, plants continuously adapt their PRRs and evolve new PRRs that facilitate detection of novel MAMPs or of other epitopes within a MAMP. Tomato, for instance, has a second flagellin receptor, FLS3, that detects an epitope (termed flgII-28) distinct from flg22 (sensed by FLS2) [38]. Similarly, rice can sense another EF-Tu epitope (EFa50) distinct from elf18 which is sensed in Brassicaceae [39]. Thus, the genetic diversity of plants provides a rich source of PRRs not only for a multitude of different MAMPs from various kinds of invaders but also for distinct MAMP epitopes.
2.4. Pattern-Triggered Immunity (PTI)
Detection of a MAMP or DAMP by its respective PRR is highly specific. However, downstream of PRRs, signaling pathways converge on common signaling hubs, often already at the receptor complex level, because certain co-receptors (e.g., BAK1, SOBIR, and CERK1) can act in multiple receptor complexes (Figure 1) [4,5,31]. Therefore, PRRs generally activate a quite stereotypic defense program referred to as PTI. Typical PTI signaling and defense responses are, for example, the depolarization of the plasma membrane, an increase in the cytosolic concentration of the secondary messenger Ca2+, activation of different protein kinases, production of reactive oxygen species (ROS), induction of defense-related genes, cell wall fortifications, and production of antimicrobial enzymes and secondary metabolites as well as defense-related plant hormones (Figure 1) [4,31]. PTI is not restricted to the site of infection but establishes in a systemic manner, resulting in an increased resistance of distal, uninfected parts of the plant to secondary infection with the same or unrelated pathogens [4,40]. Plant immune components including PRRs are systemically upregulated upon elicitor sensing, which further enables the plant to respond faster and stronger to a subsequent pathogen attack [4].
Although PTI is a temperate immune response that usually does not culminate in a hypersensitive response (HR, a programmed cell death reaction), it nevertheless constitutes a robust host barrier effective against commensals and non-adapted pathogens and maintains a basal level of immunity during infection with adapted pathogens [5]. The vital role of PTI is illustrated by the following observations: (i) Loss of individual PRRs renders plants more susceptible to infection with microbes harboring the respective MAMP [5,41,42]; (ii) Treatment of plants with MAMPs to activate PTI (which depends on the presence of a functional, matching PRR) results in enhanced resistance to subsequent local or systemic infection [5,40,41,42]; (iii) For successful host colonization pathogens need to overcome PTI by evading MAMP sensing or by releasing effectors that subvert PTI through interfering with MAMP/DAMP detection or downstream signaling and defense responses (adapted pathogens). If the effector repertoire of the pathogen sufficiently suppresses host immunity, the pathogen can establish itself in the host and cause disease (effector-triggered susceptibility, ETS) [5,43].
MAMPs are not specific to pathogens. Accordingly, commensal and beneficial microbes are also sensed by PRRs unless they have adapted their MAMP epitopes to evade detection [44,45]. Indeed, it is now evident that PTI restricts not only the growth of pathogenic microbes but also controls colonization of plant tissues with commensal and beneficial microbes [44,45,46,47]. Presumably, the diversity and functional redundancy of PRRs in combination with signal amplification through feedback-induction of genes encoding PRRs and PTI signaling components and diverse plant hormones is key to the apparent robustness of the PTI system. Indeed, breaching of host immunity against natural commensal bacteria requires inactivation of multiple PRRs simultaneously or of co-receptors shared by several PRRs [46]. In conclusion, the PRR class of immune sensors and PTI form a robust protective barrier against various types of plant colonizers and are central to plant health.
3. Prospects of Deploying PTI for Broad-Spectrum Disease Resistance Engineering in Crops
It has been known for decades that microbial and plant-derived elicitors (now usually called MAMPs and DAMPs) trigger defense responses in diverse plant species [48,49,50,51]. However, only with the molecular identification of PRRs it was possible to unravel the perception and cellular signaling mechanisms in sufficient detail to eventually exploit this central layer of the plant immune system for genetic plant protection in a purposive manner.
3.1. Pattern- versus Effector-Triggered Immunity
Because of the conserved nature and broad occurrence of MAMPs in different microbes, sensing of MAMPs to activate PTI enables the host to detect and efficiently control a wide range of microbes [4]. Adapted pathogens employ effectors to dampen PTI and to modulate host cell metabolism for their own needs. Plants, in turn, evolved immune sensors, called resistance (R) proteins that detect microbial effectors directly or indirectly by monitoring the effector targets, thus turning effectors into avirulence factors. This results in effector-triggered immunity (ETI), which includes overall similar defense responses like PTI but usually develops faster and in a stronger fashion and is mostly accompanied by an HR [5]. Since effector genes are typically specific to certain pathogen races, while R genes are limited to certain plant cultivars, this form of resistance is called race-specific or gene-for-gene resistance. In a co-evolutionary arms-race with their hosts, pathogens may be able to overcome R-gene mediated ETI relatively easy [5,52]. Mostly, recognized effectors are not essential and/or functional redundant allowing microbes to evade recognition by diversifying or even losing them. Alternatively, microbes may produce novel effectors that e.g. suppress the same or other PTI components or R protein signaling [5,52]. To date, plant resistance breeding widely relies on race-specific resistance mechanisms, largely because effectors and R genes typically result in complete or near-complete resistance. The selective pressure exerted on pathogen populations by ETI, however, may result in rapid resistance breaching in the field by the appearance of novel pathogen races/strains [52]. PTI, by contrast, enables defense against a broad spectrum of pathogens and presumably is more durable because of the evolutionary constraints on MAMP modulation and the quantitative resistance conferred by PRRs which reduces the selective pressure on the pathogen.
3.2. PRR Transfer Between Plants Species
One feature makes PRRs particularly attractive targets for genetic engineering of plant immunity: signaling networks downstream of PRRs are sufficiently conserved (Figure 1), not only within but also between plant families and even between monocot and dicot classes, to facilitate functional transfer of PRRs between them [53,54,55]. Apparently, PRRs can plug into the existing signaling network by engaging with conserved interacting signaling partners (e.g., SOBIR, BAK1 or CERK1) (Figure 2) [54,55]. Because PRRs as the ligand binding components of PTI receptor complexes determine the epitope specificity, introducing a PRR with a novel epitope specificity can confer recognition of this epitope onto previously insensitive plant species. In this way, plants can be equipped with additional MAMP sensing capacities to enhance disease resistance to pathogen classes harboring the respective MAMPs (Figure 2). These can either be PRRs for MAMPs not yet sensed by the recipient plant or for additional epitopes within a given MAMP (e.g., flgII-28/FLS3). A breakthrough study utilizing the Brassicaceae-specific EFR receptor has proven the general feasibility of this strategy [53], which was subsequently shown for multiple PRRs in various plant species (summarized in [6,56]). EFR-transgenic tomato (Solanum lycopersicum) and Nicotiana benthamiana plants gained immune responsiveness to the elf18 epitope and were more resistant to several major bacterial pathogens carrying this epitope including Ralstonia solanacearum, P. syringae, Xanthomonas perforans, and Agrobacterium tumefaciens [53]. EFR was subsequently also successfully transferred into monocots, such as wheat (Triticum aestivum) and rice (Oryza sativa) [55,57]. Similarly, transfer of the rice PRR XA21, which detects the RaxX protein from Xanthomonas oryzae pv. oryzae, to banana (Musa sp.) and sweet orange (Citrus sinensis) confers increased resistance against different Xanthomonas spp. [58,59] and expression of the PRR ELR from a wild potato species renders cultivated potato (Solanum tuberosum) more resistant to the oomycete pathogen Phytophthora infestans [60]. Taken together, these results demonstrate that addition of only a single PRR can already significantly enhance resistance of plants to adapted pathogens. Given that these pathogens are equipped with an arsenal of effectors to dampen PTI, this is quite astonishing and suggests that increased pathogen sensing provides a substantial advantage to the plant, possibly because of a stronger and/or faster onset of defense reactions.
Often, the isolation of MAMPs is the bottleneck in identifying PRRs [6,9]. However, it is not necessary to structurally identify a MAMP but sufficient to enrich it to a degree that facilitates genetic screenings for the respective PRR [61,62,63]. Furthermore, the identification of various PRRs for chemically diverse types of MAMPs has revealed that plant PRRs are typically RLKs or RLPs. Thus, these protein families can now also be systematically tested for a putative role in plant immunity, for instance through screening for altered resistance against the pathogen of interest in loss- or gain-of-function mutants or natural accessions, or through heterologous expression in suitable model plants (e.g., Arabidopsis thaliana or N. benthamiana) [64,65,66,67,68,69].
A major concern of introducing novel PRRs into plant species is that this might negatively affect beneficial interactions of these plant species with their natural microbiota and/or symbiotic interactions, such as legume-rhizobia or mycorrhizal interactions. Transgenic Medicago plants expressing EFR from Arabidopsis, however, are apparently not defective in symbiosis and are more resistant to the root pathogen R. solanacearum. Although the EFR transgenic plants showed a delay in nodule formation the final extent of nodulation and nitrogen fixation of the EFR transgenics was comparable to the wild-type plants [70]. Potentially, adapted symbionts have evolved efficient strategies to evade or suppress host PTI at various levels.
While to date mostly strong, constitutive promoters (e.g., viral promoters such as the 35S promoter of cauliflower mosaic virus, plant ubiquitin or actin promoters) are used to drive heterologous PRR expression in plants because of their universal functionality in diverse plant species and different plant tissues, this can result in unwanted side effects on general plant performance. Overexpression of RLKs, for instance, may trigger their activation in the absence of ligand and lead to growth defects because of constitutive activation of PTI or interference with developmental signaling [71]. This can be overcome by using plant promoters from e.g. endogenous PRR genes that drive PRR expression in plant tissues preferentially targeted by the pathogen of interest (e.g., root) or at sites of pathogen entry (e.g. stomata guard cells) and are strongly induced above the basal level upon infection [72,73].
The growing interest in PTI in recent years already led to the identification of numerous PRRs from different model and wild plant species and there is more to come (summarized in [6,17]). Natural diversity provides plant breeders with a versatile genetic tool box for crop improvement. Relevant PRR genes can be introduced, for instance, from wild relatives through classical breeding strategies. However, this is usually a lengthy process, bears the risk of co-segregation of unfavorable traits (linkage drag), and is not applicable to all crop species (e.g., banana, which is sterile) [56,74]. Alternatively, modern genetic engineering tools facilitate direct transfer of PRR genes across plant families beyond the constraints of sexual compatibility with the advantage that PRR genes can be quickly introduced into elite crop varieties as single traits (Figure 2). Additionally, this allows to utilize virtually any plant species as source of PRR genes. In conclusion, PRR transfer has great potential for conferring broad-spectrum and potentially durable resistance traits onto crop plants.
3.3. PRR Engineering
During co-evolution with host plants, pathogens modulate MAMP epitopes to evade host immunity whereas plants adapt their PRRs [16]. Hence, there is a natural diversity of PRRs that recognize slightly different variants of a given MAMP epitope. Ecotype collections of plant species harboring a particular PRR are thus a rich source of PRR variants with enhanced sensing capacities for these epitope variants (Figure 2) [75,76]. Additionally, in vitro evolution of PRRs can be applied to produce PRRs with altered ligand specificities [77]. With the increasing availability of PRR ectodomain structures and computational modelling tools ligand binding sites of PRRs can be modified in a directed manner to perceive a desired epitope variant or may eventually even be designed de novo (Figure 2). Potentially, ligand binding sites can be engineered at the native gene locus through CRISPR/Cas-mediated genome editing.
Some PRRs may not be able to integrate optimally with the endogenous signaling adapters upon transfer in more distantly related plant species. In such cases, full signaling competence can be restored by exchanging the transmembrane and/or intracellular signaling domain with a related PRR from the recipient species (Figure 2) [54,62]. Such chimeric PRRs have been shown to be fully functional and can be further exploited to combine different ligand binding specificities with different downstream signaling capacities (Figure 2). Some PRRs naturally induce a stronger immune response including an HR, such as the rice RLKs XA21 [78]. Sensing of chitin fragments by the rice PRR CEBiP, by contrast, does not result in HR [79]. A chimeric receptor combining the chitin-sensing ectodomain from CEBiP with the intracellular signaling domain of Xa21 activates HR upon chitin sensing and enhances resistance to the rice pathogen Magnaporthae oryzae, a pathogen that does not naturally activate Xa21 signaling [80].
3.4. Exploiting DAMP Signalling
Transfer and engineering of PRRs sensing DAMPs are also possible given that the respective DAMP is produced in the recipient species during infection with the pathogen of interest. Expression of the Arabidopsis eATP receptor DORN1 (also known as P2K1) in potato, for instance, enhances resistance to P. infestans [22,81]. Alternatively, plants can be engineered to produce the desired DAMP upon infection with diverse pathogens, for instance, by introducing a microbial enzyme producing the DAMP under control of a pathogen-responsive promoter. In this way, any pathogen that induces transcription of the transgene will trigger PTI independent of the ability to produce this DAMP itself. A promising example is the production of elicitor-active OG fragments through the balanced action of a microbial polygalacturonase (PG) and a plant PG inhibitor. Pathogen-inducible expression of this OG factory in Arabidopsis increased resistance to Botrytis cinerea, Pectobacterium carotovorum and P. syringae infections [82]. Such pathogen-responsive in situ production of DAMP signals in combination with providing the respective PRR could be a generally applicable strategy to boost PTI in any plant species and to confer resistance to a wide range of pathogens.
5. Conclusions
Most PRRs known to date have been identified from model species such as Arabidopsis, rice and recently also increasingly from tomato and potato [6,17]. These rather limited studies already illustrate the enormous resource of plant immune sensors available in different plant species that just await discovery. With novel genetic and computational tools at hand we can now exploit this natural genetic tool box for providing plants with expanded capacities to sense any kind of undesired invaders and strengthen their natural defense to resist them. Moreover, a mechanistic understanding of PRR function enables us to accelerate PRR evolution artificially, to modify PRRs in a targeted manner or even design novel PRRs.
Although PTI is generally more difficult for pathogens to overcome than ETI, pathogens will, albeit presumably on a longer time scale, eventually succeed. Hence, the deployment of single PRRs still poses the risk that pathogen populations eventually adapt their MAMP epitopes or their effector repertoire, particularly if the selection pressure on pathogen populations is constantly high. To deploy PRR engineering in a durable manner, combination of several PRR and R genes (stacking or pyramiding) appears to be a promising strategy as pathogens are unlikely to overcome several immune receptors at the same time. Possibly, resistant varieties equipped with different PRR/R gene combinations can be used in alternation to lower the selective pressure of the individual components on pathogen populations. If utilized in a thoughtful manner, these genetic tools have great power for developing truly durable and sustainable plant disease management practices.
Funding
Research in the Ranf lab is funded by the German Research Foundation through Collaborative Research Centre SFB924/TP-B10 and Emmy Noether program RA2541/1.
Acknowledgments
I thank Martin Stegmann for critical reading of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Oerke, E. Crop Losses to Animal Pests, Plant Pathogens, and Weeds. In Encyclopedia of Pest Management, Volume II; CRC Press: Boca Raton, FL, USA, 2009; pp. 116–120. [Google Scholar]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Govindan, V. The Emergence of Ug99 Races of the Stem Rust Fungus is a Threat to World Wheat Production. Annu. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Scortichini, M.; Marcelletti, S.; Ferrante, P.; Petriccione, M.; Firrao, G. Pseudomonas syringae pv. actinidiae: A re-emerging, multi-faceted, pandemic pathogen. Mol. Plant Pathol. 2012, 13, 631–640. [Google Scholar] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutrot, F.; Zipfel, C. Function, Discovery, and Exploitation of Plant Pattern Recognition Receptors for Broad-Spectrum Disease Resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Felix, G.; Duran, J.D.; Volko, S.; Boller, T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999, 18, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipfel, C.; Kunze, G.; Chinchilla, D.; Caniard, A.; Jones, J.D.G.; Boller, T.; Felix, G. Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell 2006, 125, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Ranf, S.; Scheel, D.; Lee, J. Challenges in the identification of microbe-associated molecular patterns in plant and animal innate immunity: A case study with bacterial lipopolysaccharide. Mol. Plant Pathol. 2016, 17, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Lewis, J.; Yan, S.; Liu, H.; Clarke, C.R.; Campanile, F.; Almeida, N.F.; Studholme, D.J.; Lindeberg, M.; Schneider, D.; et al. The plant pathogen Pseudomonas syringae pv. tomato is genetically monomorphic and under strong selection to evade tomato immunity. PLoS Pathog. 2011, 7, e1002130. [Google Scholar]
- Gómez-Gómez, L.; Felix, G.; Boller, T. A single locus determines sensitivity to bacterial flagellin in Arabidopsis thaliana. Plant J. 1999, 18, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Pfund, C.; Tans-Kersten, J.; Dunning, F.M.; Alonso, J.M.; Ecker, J.R.; Allen, C.; Bent, A.F. Flagellin is not a major defense elicitor in Ralstonia solanacearum cells or extracts applied to Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2004, 17, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Sun, W. Within-species flagellin polymorphism in Xanthomonas campestris pv. campestris and its impact on elicitation of Arabidopsis FLAGELLIN SENSING2-dependent defenses. Plant Cell 2006, 18, 764–779. [Google Scholar] [CrossRef] [PubMed]
- Clarke, C.R.; Chinchilla, D.; Hind, S.R.; Taguchi, F.; Miki, R.; Ichinose, Y.; Martin, G.B.; Leman, S.; Felix, G.; Vinatzer, B.A. Allelic variation in two distinct Pseudomonas syringae flagellin epitopes modulates the strength of plant immune responses but not bacterial motility. New Phytol. 2013, 200, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sun, Z.; Wang, H.; Liu, L.; Lu, F.; Yang, J.; Zhang, M.; Zhang, S.; Guo, Z.; Bent, A.F.; et al. Rice OsFLS2-mediated perception of bacterial flagellins is evaded by Xanthomonas oryzae pvs. oryzae and oryzicola. Mol. Plant 2015, 8, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- McCann, H.C.; Nahal, H.; Thakur, S.; Guttman, D.S. Identification of innate immunity elicitors using molecular signatures of natural selection. Proc. Natl. Acad. Sci. USA 2012, 109, 4215–4220. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P.-I.; Yasuda, S. Pattern recognition receptors and signaling in plant-microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranf, S. Sensing of molecular patterns through cell surface immune receptors. Curr. Opin. Plant Biol. 2017, 38, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Manosalva, P.; Manohar, M.; von Reuss, S.H.; Chen, S.; Koch, A.; Kaplan, F.; Choe, A.; Micikas, R.J.; Wang, X.; Kogel, K.-H.; et al. Conserved nematode signalling molecules elicit plant defenses and pathogen resistance. Nat. Commun. 2015, 6, 7795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, M.G.; Darvill, A.G.; Albersheim, P. Host-Pathogen Interactions: XIX. The endogenous elicitor, a fragment of a plant cell wall polysaccharide that elicits phytoalexin accumulation in soybeans. Plant Physiol. 1981, 68, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, P.; Felix, G.; Buchala, A.; Muller, C.; Métraux, J.-P. Perception of free cutin monomers by plant cells. Plant J. 1996, 10, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Tanaka, K.; Cao, Y.; Qi, Y.; Qiu, J.; Liang, Y.; Lee, S.Y.; Stacey, G. Identification of a plant receptor for extracellular ATP. Science 2014, 343, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhou, M.; Zhang, X.; Yao, J.; Zhang, Y.; Mou, Z. A lectin receptor kinase as a potential sensor for extracellular nicotinamide adenine dinucleotide in Arabidopsis thaliana. eLife 2017, 6, 267. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Manohar, M.; Manosalva, P.; Tian, M.; Moreau, M.; Klessig, D.F. Activation of Plant Innate Immunity by Extracellular High Mobility Group Box 3 and Its Inhibition by Salicylic Acid. PLoS Pathog. 2016, 12, e1005518. [Google Scholar] [CrossRef] [PubMed]
- Pearce, G.; Strydom, D.; Johnson, S.; Ryan, C.A. A polypeptide from tomato leaves induces wound-inducible proteinase inhibitor proteins. Science 1991, 253, 895–897. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Wang, X.; Chen, D.; Yang, X.; Wang, M.; Turrà, D.; Di Pietro, A.; Zhang, W. The Secreted Peptide PIP1 Amplifies Immunity through Receptor-Like Kinase 7. PLoS Pathog. 2014, 10, e1004331-15. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Ryan, C.A. Endogenous peptide defense signals in Arabidopsis differentially amplify signaling for the innate immune response. Proc. Natl. Acad. Sci. USA 2007, 104, 10732–10736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Yamashita-Yamada, M.; Hirase, T.; Fujiwara, T.; Tsuda, K.; Hiruma, K.; Saijo, Y. Danger peptide receptor signaling in plants ensures basal immunity upon pathogen-induced depletion of BAK1. EMBO J. 2016, 35, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Pearce, G.; Veyrat, N.; Erb, M.; Turlings, T.C.J.; Sartor, R.; Shen, Z.; Briggs, S.P.; Vaughan, M.M.; Alborn, H.T.; et al. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. Proc. Natl. Acad. Sci. USA 2013, 110, 5707–5712. [Google Scholar] [CrossRef] [PubMed]
- Lori, M.; van Verk, M.C.; Hander, T.; Schatowitz, H.; Klauser, D.; Flury, P.; Gehring, C.A.; Boller, T.; Bartels, S. Evolutionary divergence of the plant elicitor peptides (Peps) and their receptors: Interfamily incompatibility of perception but compatibility of downstream signalling. J. Exp. Bot. 2015, 66, 5315–5325. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Oome, S.; Raaymakers, T.M.; Cabral, A.; Samwel, S.; Böhm, H.; Albert, I.; Nürnberger, T.; Van den Ackerveken, G. Nep1-like proteins from three kingdoms of life act as a microbe-associated molecular pattern in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 16955–16960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhm, H.; Albert, I.; Oome, S.; Raaymakers, T.M.; Van den Ackerveken, G.; Nürnberger, T. A Conserved Peptide Pattern from a Widespread Microbial Virulence Factor Triggers Pattern-Induced Immunity in Arabidopsis. PLoS Pathog. 2014, 10, e1004491. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, D.; Shan, L.; He, P.; de Vries, S.; Kemmerling, B. One for all: The receptor-associated kinase BAK1. Trends Plant Sci. 2009, 14, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Gust, A.A.; Felix, G. Receptor like proteins associate with SOBIR1-type of adaptors to form bimolecular receptor kinases. Curr. Opin. Plant Biol. 2014, 21, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Brunner, F.; Nürnberger, T. Identification of immunogenic microbial patterns takes the fast lane. Proc. Natl. Acad. Sci. USA 2012, 109, 4029–4030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hind, S.R.; Strickler, S.R.; Boyle, P.C.; Dunham, D.M.; Bao, Z.; O’Doherty, I.M.; Baccile, J.A.; Hoki, J.S.; Viox, E.G.; Clarke, C.R.; et al. Tomato receptor FLAGELLIN-SENSING 3 binds flgII-28 and activates the plant immune system. Nat. Plants 2016, 2, 16128. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Inagaki, H.; Takai, R.; Hirai, H.; Che, F.-S. Two distinct EF-Tu epitopes induce immune responses in rice and Arabidopsis. Mol. Plant-Microbe Interact. 2014, 27, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Mishina, T.E.; Zeier, J. Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in Arabidopsis. Plant J. 2007, 50, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Robatzek, S.; Navarro, L.; Oakeley, E.J.; Jones, J.D.G.; Felix, G.; Boller, T. Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 2004, 428, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Ranf, S.; Gisch, N.; Schäffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Sánchez-Carballo, P.M.; Zähringer, U.; Hückelhoven, R.; Lee, J.; et al. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-Pathogen Effectors: Cellular Probes Interfering with Plant Defenses in Spatial and Temporal Manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Spaepen, S.; Garrido-Oter, R.; Schulze-Lefert, P. Interplay between Innate Immunity and the Plant Microbiota. Annu. Rev. Phytopathol. 2017, 55, 565–589. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Oldroyd, G.E.D. Plant signalling in symbiosis and immunity. Nat. Publ. Group 2017, 543, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.-F.; Nomura, K.; Aung, K.; Velásquez, A.C.; Yao, J.; Boutrot, F.; Chang, J.H.; Zipfel, C.; He, S.Y. Bacteria establish an aqueous living space in plants crucial for virulence. Nature 2016, 539, 524–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Halane, M.K.; Gassmann, W.; Stacey, G. The Role of Plant Innate Immunity in the Legume-Rhizobium Symbiosis. Annu. Rev. Plant Biol. 2017, 68, 535–561. [Google Scholar] [CrossRef] [PubMed]
- Albersheim, P.; Valent, B.S. Host-pathogen interactions in plants. Plants, when exposed to oligosaccharides of fungal origin, defend themselves by accumulating antibiotics. J. Cell Biol. 1978, 78, 627–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somssich, I.E.; Science, K. Pathogen defence in plants—A paradigm of biological complexity. Trends Plant Sci. 1998, 3, 86–90. [Google Scholar] [CrossRef]
- Ayers, A.R.; Valent, B.; Ebel, J.; Albersheim, P. Host-Pathogen Interactions: XI. Composition and Structure of Wall-released Elicitor Fractions. Plant Physiol. 1976, 57, 766–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A.; Fuller, K.W. Characterization of components from culture filtrates of Botrytis cinerea which stimulate phaseollin biosynthesis in Phaseolus vulgaris cell suspension cultures. Physiol. Plant Pathol. 1977, 11, 287–296. [Google Scholar] [CrossRef]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the Plant Immune System from Dissection to Deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacombe, S.E.V.; Rougon-Cardoso, A.; Sherwood, E.; Peeters, N.; Dahlbeck, D.; Van Esse, H.P.; Smoker, M.; Rallapalli, G.; Thomma, B.P.H.J.; Staskawicz, B.; et al. Interfamily transfer of a plant pattern-recognition receptor confers broad-spectrum bacterial resistance. Nat. Biotechnol. 2010, 28, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Holton, N.; Nekrasov, V.; Ronald, P.C.; Zipfel, C. The Phylogenetically-Related Pattern Recognition Receptors EFR and XA21 Recruit Similar Immune Signaling Components in Monocots and Dicots. PLoS Pathog. 2015, 11, e1004602-22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwessinger, B.; Bahar, O.; Thomas, N.; Holton, N.; Nekrasov, V.; Ruan, D.; Canlas, P.E.; Daudi, A.; Petzold, C.J.; Singan, V.R.; et al. Transgenic Expression of the Dicotyledonous Pattern Recognition Receptor EFR in Rice Leads to Ligand-Dependent Activation of Defense Responses. PLoS Pathog. 2015, 11, e1004809. [Google Scholar]
- Rodriguez-Moreno, L.; Song, Y.; Thomma, B.P. Transfer and engineering of immune receptors to improve recognition capacities in crops. Curr. Opin. Plant Biol. 2017, 38, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Schoonbeek, H.-J.; Wang, H.-H.; Stefanato, F.L.; Craze, M.; Bowden, S.; Wallington, E.; Zipfel, C.; Ridout, C.J. Arabidopsis EF-Tu receptor enhances bacterial disease resistance in transgenic wheat. New Phytol. 2015, 206, 606–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, J.N.; Lorenzen, J.; Bahar, O.; Ronald, P.; Tripathi, L. Transgenic expression of the rice Xa21 pattern-recognition receptor in banana (Musa sp.) confers resistance to Xanthomonas campestris pv. musacearum. Plant Biotechnol. J. 2014, 12, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.A.; Murata, M.M.; El-Shamy, H.A.; Graham, J.H.; Grosser, J.W. Enhanced resistance to citrus canker in transgenic mandarin expressing Xa21 from rice. Transgenic Res. 2018, 27, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Verzaux, E.; Chaparro-Garcia, A.; Bijsterbosch, G.; Keizer, L.C.P.; Zhou, J.; Liebrand, T.W.H.; Xie, C.; Govers, F.; Robatzek, S.; et al. Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato. Nat. Plants 2015, 1, 15034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Fraiture, M.; Kolb, D.; Loffelhardt, B.; Desaki, Y.; Boutrot, F.F.G.; Tor, M.; Zipfel, C.; Gust, A.A.; Brunner, F. Arabidopsis RECEPTOR-LIKE PROTEIN30 and Receptor-Like Kinase SUPPRESSOR OF BIR1-1/EVERSHED Mediate Innate Immunity to Necrotrophic Fungi. Plant Cell 2013, 25, 4227–4241. [Google Scholar] [CrossRef] [PubMed]
- Jehle, A.K.; Lipschis, M.; Albert, M.; Fallahzadeh-Mamaghani, V.; Fürst, U.; Mueller, K.; Felix, G. The receptor-like protein ReMAX of Arabidopsis detects the microbe-associated molecular pattern eMax from Xanthomonas. Plant Cell 2013, 25, 2330–2340. [Google Scholar] [CrossRef] [PubMed]
- Hegenauer, V.; Fürst, U.; Kaiser, B.; Smoker, M.; Zipfel, C.; Felix, G.; Stahl, M.; Albert, M. Detection of the plant parasite Cuscuta reflexa by a tomato cell surface receptor. Science 2016, 353, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Mott, G.A.; Thakur, S.; Smakowska, E.; Wang, P.W.; Belkhadir, Y.; Desveaux, D.; Guttman, D.S. Genomic screens identify a new phytobacterial microbe-associated molecular pattern and the cognate Arabidopsis receptor-like kinase that mediates its immune elicitation. Genome Biol. 2016, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nsibo, D.L.; Juhar, H.M.; Govers, F.; Bouwmeester, K. Ectopic expression of Arabidopsis L-type lectin receptor kinase genes LecRK-I.9 and LecRK-IX.1 in Nicotiana benthamiana confers Phytophthora resistance. Plant Cell Rep. 2016, 35, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Delteil, A.; Gobbato, E.; Cayrol, B.; Estevan, J.; Michel-Romiti, C.; Dievart, A.; Kroj, T.; Morel, J.-B. Several wall-associated kinases participate positively and negatively in basal defense against rice blast fungus. BMC Plant Biol. 2016, 16, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wu, H.; Chen, H.; Liu, Y.; He, J.; Kang, H.; Sun, Z.; Pan, G.; Wang, Q.; Hu, J.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2015, 33, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Weide, R.; Govers, F.; Bouwmeester, K. L-type lectin receptor kinases in Nicotiana benthamiana and tomato and their role in Phytophthora resistance. J. Exp. Bot. 2015, 66, 6731–6743. [Google Scholar] [CrossRef] [PubMed]
- Bourdais, G.; Burdiak, P.; Gauthier, A.; Nitsch, L.; Salojärvi, J.; Rayapuram, C.; Idänheimo, N.; Hunter, K.; Kimura, S.; Merilo, E.; et al. On behalf of the CRK Consortium. Large-Scale Phenomics Identifies Primary and Fine-Tuning Roles for CRKs in Responses Related to Oxidative Stress. PLoS Genet. 2015, 11, e1005373. [Google Scholar] [CrossRef] [PubMed]
- Pfeilmeier, S.; George, J.; Morel, A.; Roy, S.; Smoker, M.; Stansfeld, L.; Downie, A.; Peeters, N.; Malone, J.; Zipfel, C. Heterologous expression of the immune receptor EFR in Medicago truncatula reduces pathogenic infection, but not rhizobial symbiosis. bioRxiv 2017. [Google Scholar] [CrossRef]
- Kim, S.Y.; Shang, Y.; Joo, S.-H.; Kim, S.-K.; Nam, K.H. Overexpression of BAK1 causes salicylic acid accumulation and deregulation of cell death control genes. Biochem. Biophys. Res. Commun. 2017, 484, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Wyrsch, I.; Strutt, J.; Wimalasekera, R.; Webb, A.; Boller, T.; Robatzek, S. Expression patterns of FLAGELLIN SENSING 2 map to bacterial entry sites in plant shoots and roots. J. Exp. Bot. 2014, 65, 6487–6498. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, C.; Robatzek, S. Plants and pathogens: Putting infection strategies and defence mechanisms on the map. Curr. Opin. Plant Biol. 2012, 15, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Gust, A.A.; Brunner, F.D.R.; Nürnberger, T. Biotechnological concepts for improving plant innate immunity. Curr. Opin. Biotechnol. 2010, 21, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Trdá, L.; Fernandez, O.; Boutrot, F.; Héloir, M.-C.; Kelloniemi, J.; Daire, X.; Adrian, M.; Clément, C.; Zipfel, C.; Dorey, S.; et al. The grapevine flagellin receptor VvFLS2 differentially recognizes flagellin-derived epitopes from the endophytic growth-promoting bacterium Burkholderia phytofirmans and plant pathogenic bacteria. New Phytol. 2013, 201, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Febres, V.J.; Jones, J.B.; Moore, G.A. A survey of FLS2 genes from multiple citrus species identifies candidates for enhancing disease resistance to Xanthomonas citri ssp. citri. Hortic. Res. 2016, 3, 80. [Google Scholar] [CrossRef] [PubMed]
- Helft, L.; Thompson, M.; Bent, A.F. Directed Evolution of FLS2 towards Novel Flagellin Peptide Recognition. PLoS ONE 2016, 11, e0157155. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Kouzai, Y.; Kaku, H.; Shibuya, N.; Minami, E.; Nishizawa, Y. Perception of the chitin oligosaccharides contributes to disease resistance to blast fungus Magnaporthe oryzae in rice. Plant J. 2010, 64, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Kouzai, Y.; Kaku, H.; Shibuya, N.; Minami, E.; Nishizawa, Y. Enhancement of MAMP signaling by chimeric receptors improves disease resistance in plants. Plant Signal. Behav. 2011, 6, 449–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouwmeester, K.; Han, M.; Blanco-Portales, R.; Song, W.; Weide, R.; Guo, L.-Y.; van der Vossen, E.A.G.; Govers, F. The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to Phytophthora infestans in Solanaceous plants. Plant Biotechnol. J. 2014, 12, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Pontiggia, D.; Raggi, S.; Cheng, Z.; Scaloni, F.; Ferrari, S.; Ausubel, F.M.; Cervone, F.; De Lorenzo, G. Plant immunity triggered by engineered in vivo release of oligogalacturonides, damage-associated molecular patterns. Proc. Natl. Acad. Sci. USA 2015, 112, 5533–5538. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Scheme of typical MAMP and DAMP-activated immune responses in plant cells. MAMPs and DAMPs (star symbols) are sensed by specific host PRRs localized at the cell surface. Signaling pathways downstream of PRRs converge on common signaling components, for instance co-receptors (CoR) or signaling HUBS. PTI responses include for example fluxes of various ions—including the secondary messenger Ca2+—across the plasma membrane, production of ROS by RBOH-type oxidases, activation of calcium-dependent and mitogen-activated protein kinases (CDPKs and MAPKs), gene expression changes, production of antimicrobial compounds, and fortification of the plant cell wall.
Figure 1.
Scheme of typical MAMP and DAMP-activated immune responses in plant cells. MAMPs and DAMPs (star symbols) are sensed by specific host PRRs localized at the cell surface. Signaling pathways downstream of PRRs converge on common signaling components, for instance co-receptors (CoR) or signaling HUBS. PTI responses include for example fluxes of various ions—including the secondary messenger Ca2+—across the plasma membrane, production of ROS by RBOH-type oxidases, activation of calcium-dependent and mitogen-activated protein kinases (CDPKs and MAPKs), gene expression changes, production of antimicrobial compounds, and fortification of the plant cell wall.
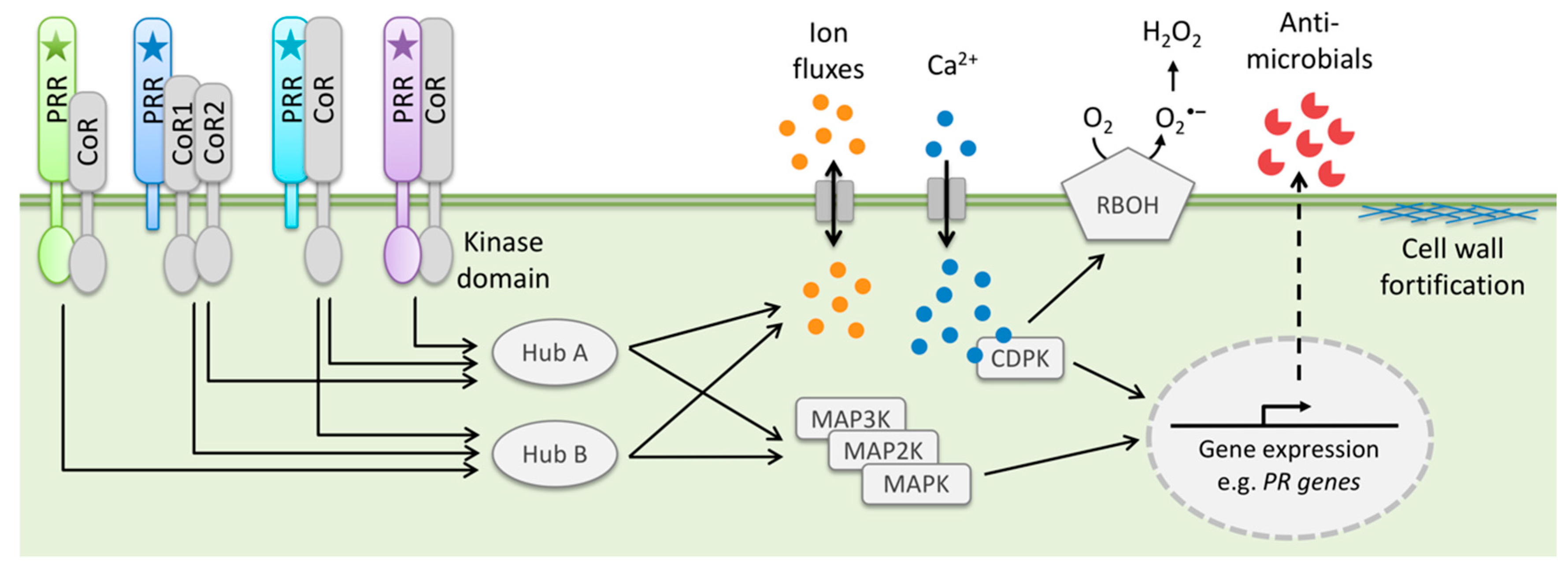
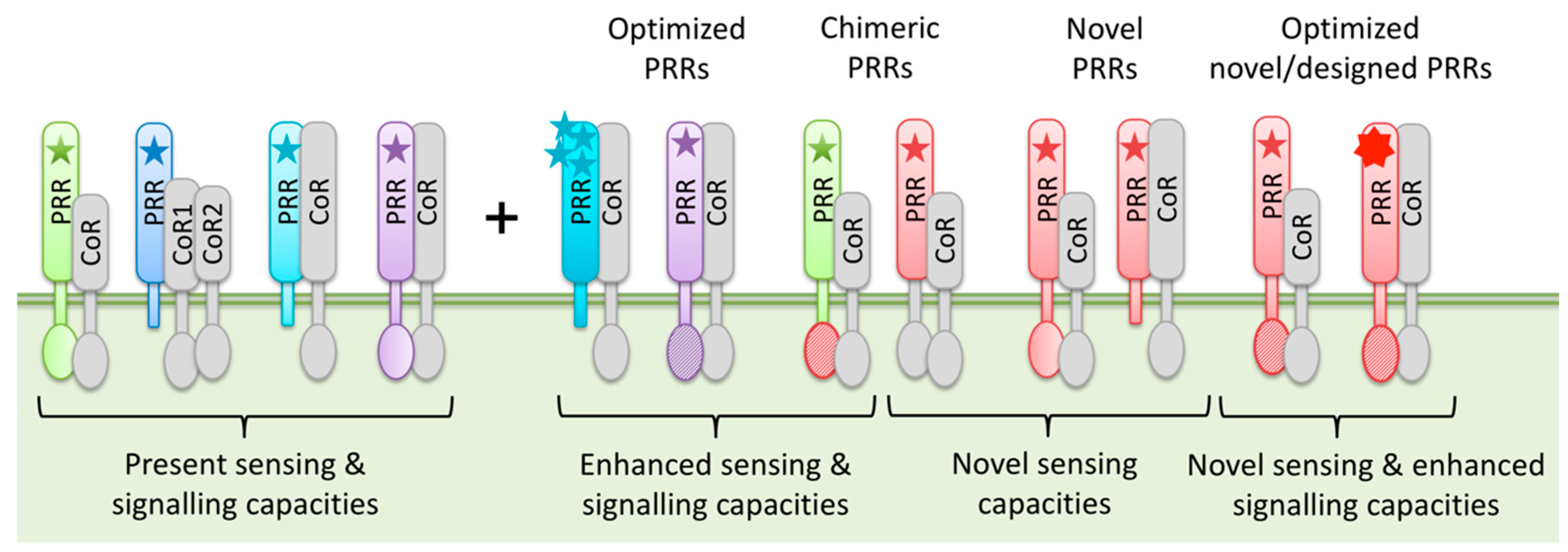
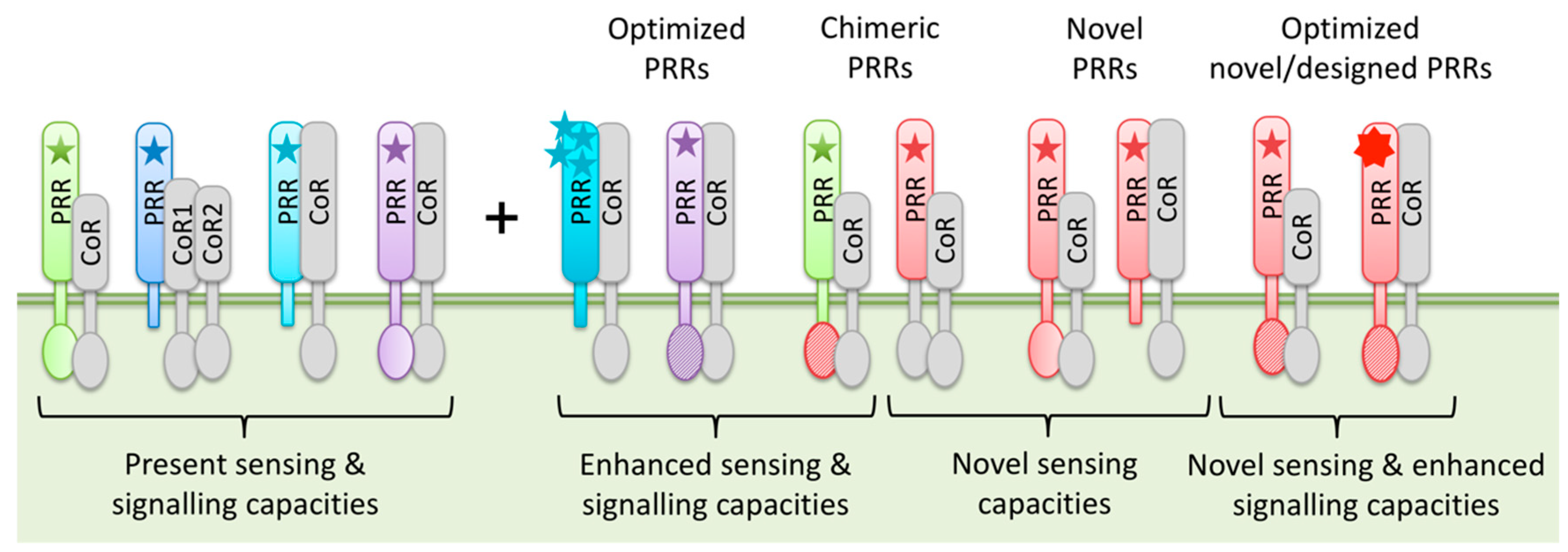
Figure 2.
Schematic representation of potential PRR transfer and engineering options. PRR-mediated sensing of MAMPs/DAMPs (star symbols) and PRR signaling capacities can be enhanced by optimization of endogenous PRRs or introduction of novel, chimeric or even “designer” PRRs. This equips plants with additional MAMP/DAMP sensing and improved signaling capacities and results in enhanced PTI against a broad spectrum of invaders.
Figure 2.
Schematic representation of potential PRR transfer and engineering options. PRR-mediated sensing of MAMPs/DAMPs (star symbols) and PRR signaling capacities can be enhanced by optimization of endogenous PRRs or introduction of novel, chimeric or even “designer” PRRs. This equips plants with additional MAMP/DAMP sensing and improved signaling capacities and results in enhanced PTI against a broad spectrum of invaders.
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ranf, S. Pattern Recognition Receptors—Versatile Genetic Tools for Engineering Broad-Spectrum Disease Resistance in Crops. Agronomy 2018, 8, 134. https://doi.org/10.3390/agronomy8080134
AMA Style
Ranf S. Pattern Recognition Receptors—Versatile Genetic Tools for Engineering Broad-Spectrum Disease Resistance in Crops. Agronomy. 2018; 8(8):134. https://doi.org/10.3390/agronomy8080134
Chicago/Turabian StyleRanf, Stefanie. 2018. "Pattern Recognition Receptors—Versatile Genetic Tools for Engineering Broad-Spectrum Disease Resistance in Crops" Agronomy 8, no. 8: 134. https://doi.org/10.3390/agronomy8080134
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.