Seed Germination in Relation to Total Sugar and Starch in Endosperm Mutant of Sweet Corn Genotypes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Laboratory Experiment
2.2.1. Seed Quality
2.2.2. Dry Weight of Seed Parts
2.2.3. Chemical Analyses
2.3. Statistical Analysis
3. Results
3.1. Seed Quality
3.2. Weights of Pericarp, Embryo, and Endosperm, Total Sugar Content, and Starch Content
3.3. Correlations among Traits Related to Seed Quality and Carbohydrate Characters
3.4. Application in Seed Production
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lertrat, K.; Pulam, T. Breeding for increased sweetness in sweet corn. Int. J. Plant Breed. 2017, 1, 27–30. [Google Scholar]
- Creech, R.G. Genetic control of carbohydrate synthesis in maize endosperm. Genetics 1965, 52, 1175–1186. [Google Scholar] [PubMed]
- Styer, R.C.; Cantliffe, D.J.; Hannah, L.C. Differential seed and seedling vigor in shrunken-2 compared to three other genotypes of corn at various stages of development. J. Am. Soc. Hortic. Sci. 1980, 105, 329–332. [Google Scholar]
- Tracy, W.F. Sweet corn. In Specialty Corns, 2nd ed; Hallauer, A.R., Ed.; CRC: Boca Raton, FL, USA, 2001; pp. 155–198. [Google Scholar]
- Styer, R.C.; Cantliffe, D.J. Dependence of seed vigor during germination on carbohydrate source in endosperm mutants of maize. Plant Physiol. 1984, 76, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Callan, N.W.; Mathre, D.E.; Miller, J.B. Bio-priming seed treatment for biological control of Pythium ultimum pre-emergence damping-off in sh2 sweet corn. Plant Dis. 1990, 74, 368–372. [Google Scholar] [CrossRef]
- Parera, C.A.; Cantliffe, D.J. Presowing seed treatments to enhance supersweet corn seed and seedling quality. HortScience 1994, 29, 277–278. [Google Scholar]
- Tracy, W.F. History, genetics, and breeding of supersweet (shrunken2) sweet corn. Plant Breed. Rev. 1997, 14, 189–236. [Google Scholar]
- Ordas, B.; Revilla, P.; Ordas, A.; Malvar, R.A. Hybrids sugary x sugary enhancer of sweet corn: A valuable option for cool environments. Sci. Hort. 2008, 118, 111–114. [Google Scholar] [CrossRef]
- Lertrat, K.; Thongnarin, N. Novel approach to eating quality improvement in local waxy corn: Improvement of sweet taste in local waxy corn variety with mixed kernels from super sweet corn. Acta Hoertic. 2008, 769, 145–150. [Google Scholar] [CrossRef]
- Carey, E.E.; Dickinson, D.B.; Rhodes, A.M. Sugar characteristics of sweet corn populations from a sugary enhancer breeding program. Euphytica 1984, 33, 609–622. [Google Scholar] [CrossRef]
- Available online: https://www.maizegdb.org (accessed on 6 December 2018).
- Simla, S.; Lertrat, K.; Suriharn, B. Combinations of multiple genes controlling endosperm characters in relation to maximum eating quality of vegetable waxy corn. SABRAO J. Breed. Genet. 2016, 48, 210–218. [Google Scholar]
- Borowski, A.M.; Fritz, V.A.; Waters, L. Seed maturity influences germination and vigor of two shrunken-2 sweet corn hybrids. J. Am. Soc. Hortic. Sci. 1991, 116, 401–404. [Google Scholar]
- Ketthaisong, D.; Suriharn, B.; Tangwongchai, R.; Lertrat, K. Changes in physicochemical properties of waxy corn starches at different stages of harvesting. Carbohydr. Polym. 2013, 98, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugar and related substance. Anal. Chem. 1956, 2, 350–356. [Google Scholar] [CrossRef]
- Gomaz, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons: New York, NY, USA, 1984; p. 680. [Google Scholar]
- Parera, C.A.; Cantliffe, D.J.; Rtoffella, P.J.; Scully, B.T. Field emergence of shrunken-2 corn predicted by single- and multiple-vigor laboratory. J. Am. Soc. Hortic. Sci. 1995, 120, 128–132. [Google Scholar]
- Adetimirin, V.O.; Kim, S.K.; Szczech, M. Factors associated with emergence of shrunken-2 maize in Korea. J. Agric. Sci. 2006, 144, 63–68. [Google Scholar] [CrossRef]
- Cao, D.D.; Hu, J.; Huang, X.X.; Wang, X.J.; Guan, Y.J.; Wang, Z.F. Relationship between changes of kernel nutritive components and seed vigor during development stages of F1 seeds of sh2 sweet corn. J. Zhejiang Univ. Sci. B 2008, 9, 964–968. [Google Scholar] [CrossRef]
- Simla, S.; Lertrat, K.; Suriharn, B. Gene effects of sugar compositions in waxy corn. Asian J. Plant Sci. 2009, 8, 417–424. [Google Scholar]
- Vitor, H.V.M.; Silvio, M.C.; Durval, D.N.; Tulio, L.P.; Marcos, A.N.D. Effects of seed vigor on intra-specific competition and grain yield in maize. Agron. J. 2012, 105, 222–228. [Google Scholar]
- Liu, J.B.; Fu, Z.Y.; Xie, H.L.; Hu, Y.M.; Liu, Z.H.; Duan, L.J.; Xu, S.Z.; Tang, J.H. Identification of QTLs for maize seed vigor at three stages of seed maturity using a RIL population. Euphytica 2011, 178, 127–135. [Google Scholar]
- Churchill, G.A.; Andrew, R.H. Effect of two maize endosperm mutants on kernel maturity, carbohydrates, and germination. Crop Sci. 1984, 24, 76–81. [Google Scholar] [CrossRef]
- Boyer, C.D.; Shannon, J.C. The use of the endosperm genes for sweet corn improvement. Plant Breed. Rev. 1984, 1, 139–161. [Google Scholar]
- Li, L.; Blanco, M.; Jane, J. Physicochemical properties of endosperm and pericarp starches during maize development. Carbohydr. Polym. 2007, 67, 630–639. [Google Scholar] [CrossRef]
- Boyer, C.D.; Hannah, L.C. Kernel mutants of corn. In Specialty Corns, 2nd ed.; Hallauer, A.R., Ed.; CRC press: New York, NY, USA; Washington, DC, USA, 2001; pp. 1–31. [Google Scholar]
- Sanwo, M.M.; DeMason, D.A. Characteristics of -amylase during germination of two high-sugar sweet cultivars of Zea mays L. Plant Physiol. 1992, 99, 1184–1192. [Google Scholar] [CrossRef]
- Xu, J.G.; Hu, Q.P.; Wang, X.D.; Luo, J.Y.; Liu, Y.; Tain, C.R. Changes in the main nutrients, phytochemicals and antioxidant activity in yellow corn grain during maturation. J. Agric. Food Chem. 2010, 58, 5751–5756. [Google Scholar] [CrossRef] [PubMed]
- Simla, S.; Lertrat, K.; Suriharn, B. Carbohydrate characters of six vegetable waxy corn varieties as affected by harvest time and storage duration. Asian J. Plant Sci. 2010, 9, 463–470. [Google Scholar] [CrossRef]
- Vashisth, A.; Nagarajan, S. Germination characteristics of seeds of maize (Zea mays L.) exposed to magnetic fields under accelerated aging condition. Int. Agrophys. 2009, 9, 50–58. [Google Scholar]
- Shah, F.S.; Watson, C.E.; Cabrera, E.R. Seed vigor testing of subtropical corn hybrids. MAFES 2002, 23, 1–5. [Google Scholar]
- Noli, E.; Casarini, E.; Urso, G.; Conti, S. Suitability of three vigour test procedures to predict field performance of early sown maize seed. Seed Sci. Technol. 2008, 36, 168–176. [Google Scholar] [CrossRef]
- Wann, E.V. Seed vigor and respiration of maize kernels with different endosperm genotypes. J. Am. Soc. Hortic. Sci. 1980, 105, 31–34. [Google Scholar]
- Azanza, V.; Zur, A.B.; Juvik, J.A. Variation in sweet corn kernel characteristics associated with stand establishment and eating quality. Euphytica 1996, 87, 7–18. [Google Scholar] [CrossRef]


{kind=link}
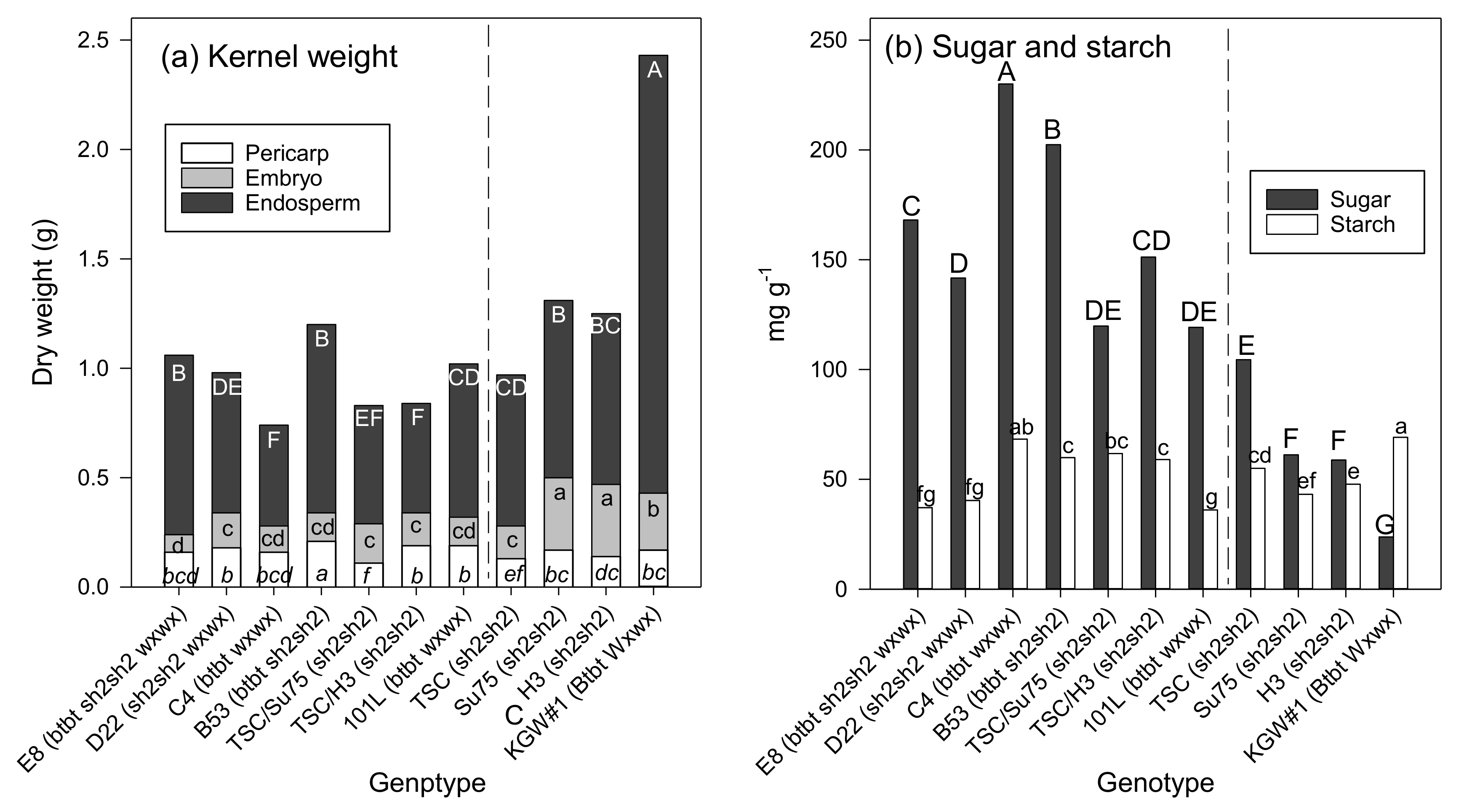{kind=link}
| Lines/Hybrids | Types | Genotypes | Phenotypes |
|---|---|---|---|
| 1. D8 | Inbred line; Triple recessive genes | btbt shsh2 wxwx | shrunken |
| 2. D22 | Inbred line; Double recessive genes | btbt ShSh2 wxwx | shrunken |
| 3. C4 | Inbred line; Double recessive genes | BtBt shsh2 wxwx | shrunken |
| 4. B52 | Inbred line; Double recessive genes | btbt shsh2 WxWx | shrunken |
| 5. Su75/TSC | Inbred line; Single recessive gene | BtBt shsh2 WxWx | shrunken |
| 6. H3/TSC | Inbred line; Single recessive gene | BtBt shsh2 WxWx | shrunken |
| 7. Su75 | F1 hybrid | BtBt shsh2 WxWx | shrunken |
| 8. H3 | F1 hybrid | BtBt shsh2 WxWx | shrunken |
| 9. TSC | F1 hybrid | BtBt shsh2 WxWx | shrunken |
| 10. 101bt | Inbred line | btbt ShSh2 wxwx | shrunken |
| 11. KGW1 | F1 hybrid | Btbt ShSh2 wxwx | flint |
| Line/Hybrid | Genotype | Germination | Speed of Germination | Seedling Length (cm) | Seedling Dry Weight | |
|---|---|---|---|---|---|---|
| (%) | (No. of plants/day) | Stem | Root | (g) | ||
| BP test | ||||||
| E8 | btbt sh2sh2 wxwx | 79.0 e | 8.2 e | 8.0 de | 10.5 de | 0.043 def |
| D22 | sh2sh2 wxwx | 40.0 f | 9.2 de | 6.3 ef | 8.2 ef | 0.066 ab |
| C4 | btbt wxwx | 36.0 f | 6.0 f | 5.7 f | 6.3 f | 0.040 ef |
| B53 | btbt sh2sh2 | 83.0 de | 9.7 d | 8.5 cd | 12.1 bcd | 0.068 a |
| TSC/Su75 | sh2sh2 | 88.0 cd | 11.5 b | 10.7 ab | 13.7 ab | 0.045 def |
| TSC/H3 | sh2sh2 | 96.7 ab | 12.0 b | 10.1 bc | 11.0 cd | 0.050 cd |
| 101L | btbt wxwx | 40.7 f | 5.1 f | 8.3 cde | 10.4 de | 0.035 f |
| TSC | sh2sh2 | 84.7 cde | 10.0 cd | 9.0 bcd | 13.4 abc | 0.057 bc |
| Su75 | sh2sh2 | 90.7 bc | 13.7 a | 12.7 a | 13.5 abc | 0.050 cde |
| H3 | sh2sh2 | 98.0 a | 13.4 a | 12.5 a | 15.9 a | 0.067 ab |
| KGW#1 | Btbt Wxwx | 96.7 ab | 11.0 bc | 10.9 ab | 11.2 bcd | 0.064 ab |
| AA test | ||||||
| E8 | btbt sh2sh2 wxwx | 16.0 e | 2.0 h | 4.8 ef | 4.0 e | 0.059 abc |
| D22 | sh2sh2 wxwx | 39.0 c | 4.9 e | 9.6 cd | 8.1 d | 0.068 a |
| C4 | btbt wxwx | 3.0 f | 0.4 i | 3.4 g | 2.4 f | 0.028 d |
| B53 | btbt sh2sh2 | 24.0 d | 4.0 f | 4.3 fg | 3.7 ef | 0.036 d |
| TSC/Su75 | sh2sh2 | 58.0 b | 8.0 c | 10.8 bc | 12.7 ab | 0.065 a |
| TSC/H3 | sh2sh2 | 44.7 c | 6.7 d | 11.5 b | 11.6 bc | 0.053 bc |
| 101L | btbt wxwx | 12.7 e | 1.6 h | 5.7 e | 3.7 ef | 0.035 d |
| TSC | sh2sh2 | 24.7 d | 2.9 g | 8.4 d | 7.8 d | 0.048 c |
| Su75 | sh2sh2 | 57.0 b | 8.0 c | 10.8 bc | 11.4 bc | 0.058 abc |
| H3 | sh2sh2 | 89.3 a | 13.3 a | 13.1a | 14.0 a | 0.061 ab |
| KGW#1 | Btbt Wxwx | 87.3 a | 10.6 b | 9.6 cd | 10.7 c | 0.058 abc |
| BP Test | AA Test | |||
|---|---|---|---|---|
| Parameters | Germination | Speed of Germination | Germination | Speed of Germination |
| Speed of germination | 0.83 ** | 0.99 ** | ||
| Stem length | 0.83 ** | 0.84 ** | 0.83 ** | 0.85 ** |
| Root length | 0.79 ** | 0.74 ** | 0.88 ** | 0.90 ** |
| Seedling dry weight | 0.39 | 0.53 | 0.66 * | 0.63 * |
| Pericarp wt. | −0.24 | −0.28 | −0.28 | −0.27 |
| Embryo wt. | 0.50 | 0.75 ** | 0.85 ** | 0.86 ** |
| Endosperm wt. | 0.36 | 0.18 | 0.53 | 0.43 |
| Total sugar | −0.52 | −0.59 | −0.82 ** | −0.76 ** |
| Total starch | 0.26 | 0.13 | 0.19 | 0.17 |
| BP Test | AA Test | |||
|---|---|---|---|---|
| Parameters | Germination | Speed of Germination | Germination | Speed of Germination |
| Speed of germination | 0.83 ** | 0.99 ** | ||
| Stem length | 0.83 ** | 0.84 ** | 0.83 ** | 0.85 ** |
| Root length | 0.79 ** | 0.74 ** | 0.88 ** | 0.90 ** |
| Seedling dry weight | 0.39 | 0.53 | 0.66 * | 0.63 * |
| Pericarp wt. | −0.24 | −0.28 | −0.28 | −0.27 |
| Embryo wt. | 0.50 | 0.75 ** | 0.85 ** | 0.86 ** |
| Endosperm wt. | 0.36 | 0.18 | 0.53 | 0.43 |
| Total sugar | −0.52 | −0.59 | −0.82 ** | −0.76 ** |
| Total starch | 0.26 | 0.13 | 0.19 | 0.17 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pairochteerakul, P.; Jothityangkoon, D.; Ketthaisong, D.; Simla, S.; Lertrat, K.; Suriharn, B. Seed Germination in Relation to Total Sugar and Starch in Endosperm Mutant of Sweet Corn Genotypes. Agronomy 2018, 8, 299. https://doi.org/10.3390/agronomy8120299
Pairochteerakul P, Jothityangkoon D, Ketthaisong D, Simla S, Lertrat K, Suriharn B. Seed Germination in Relation to Total Sugar and Starch in Endosperm Mutant of Sweet Corn Genotypes. Agronomy. 2018; 8(12):299. https://doi.org/10.3390/agronomy8120299
Chicago/Turabian StylePairochteerakul, Potchanee, Darunee Jothityangkoon, Danupol Ketthaisong, Sakunkan Simla, Kamol Lertrat, and Bhalang Suriharn. 2018. "Seed Germination in Relation to Total Sugar and Starch in Endosperm Mutant of Sweet Corn Genotypes" Agronomy 8, no. 12: 299. https://doi.org/10.3390/agronomy8120299
APA StylePairochteerakul, P., Jothityangkoon, D., Ketthaisong, D., Simla, S., Lertrat, K., & Suriharn, B. (2018). Seed Germination in Relation to Total Sugar and Starch in Endosperm Mutant of Sweet Corn Genotypes. Agronomy, 8(12), 299. https://doi.org/10.3390/agronomy8120299