Threshold Tolerance of New Genotypes of Pennisetum glaucum (L.) R. Br. to Salinity and Drought

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Growth Conditions
2.2. Plant Material
2.3. Laboratory Germination Conditions
2.4. Laboratory Growth Conditions
2.5. Water and Proline Contents
2.6. Lipid Peroxidation
2.7. Pigment Extraction and Quantification
2.8. Chl Fluorescence
2.9. Statistical Analysis
3. Results
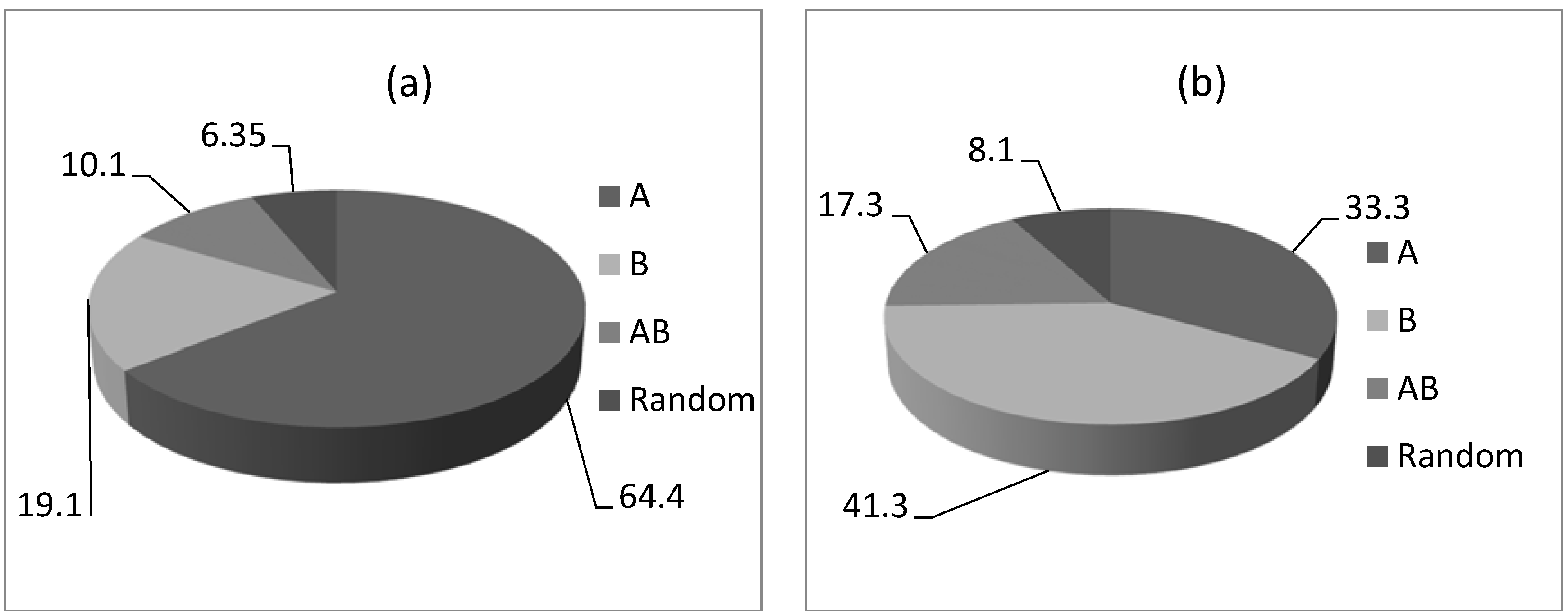
3.1. Field Trials
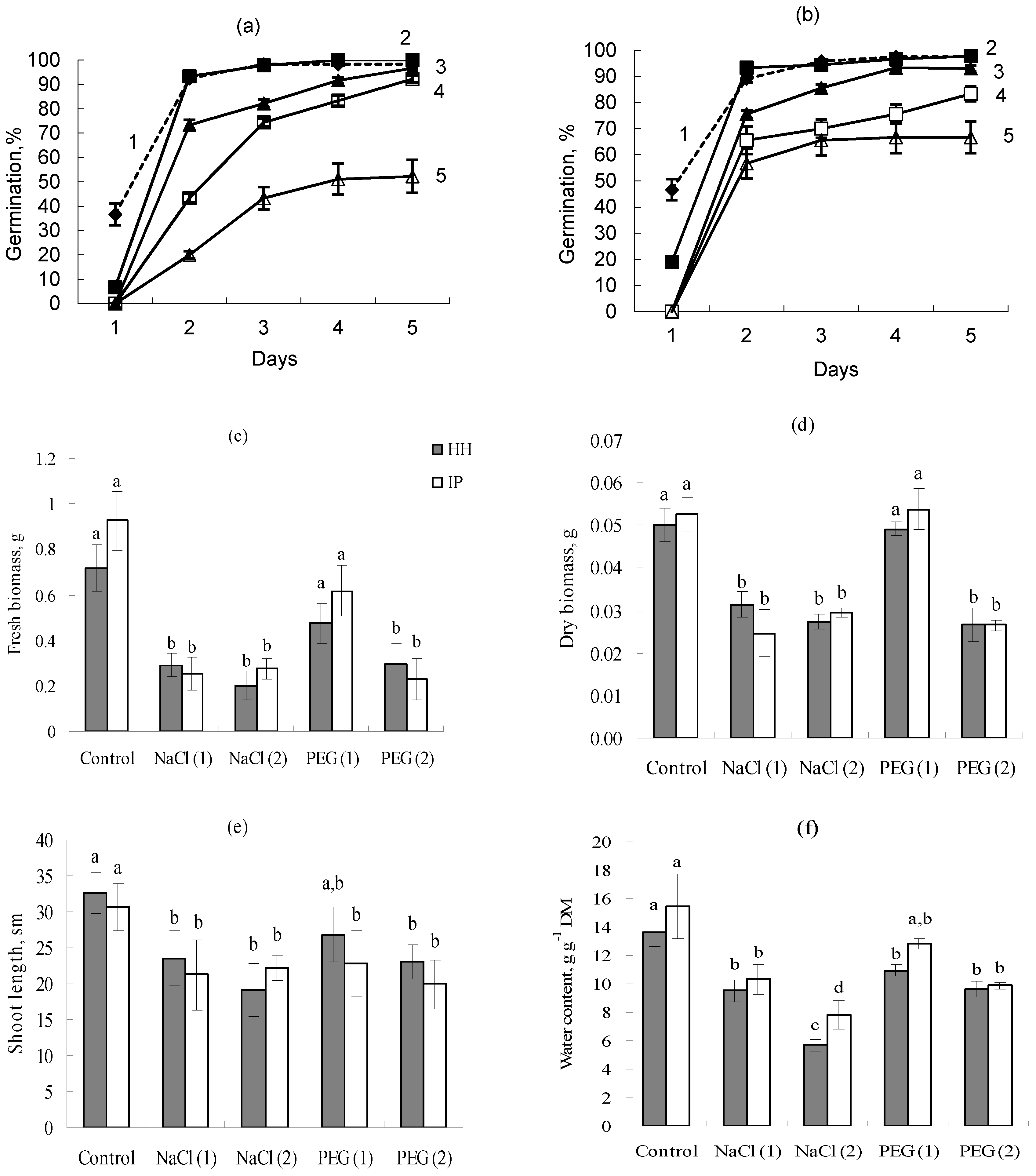
3.2. Laboratory Germination
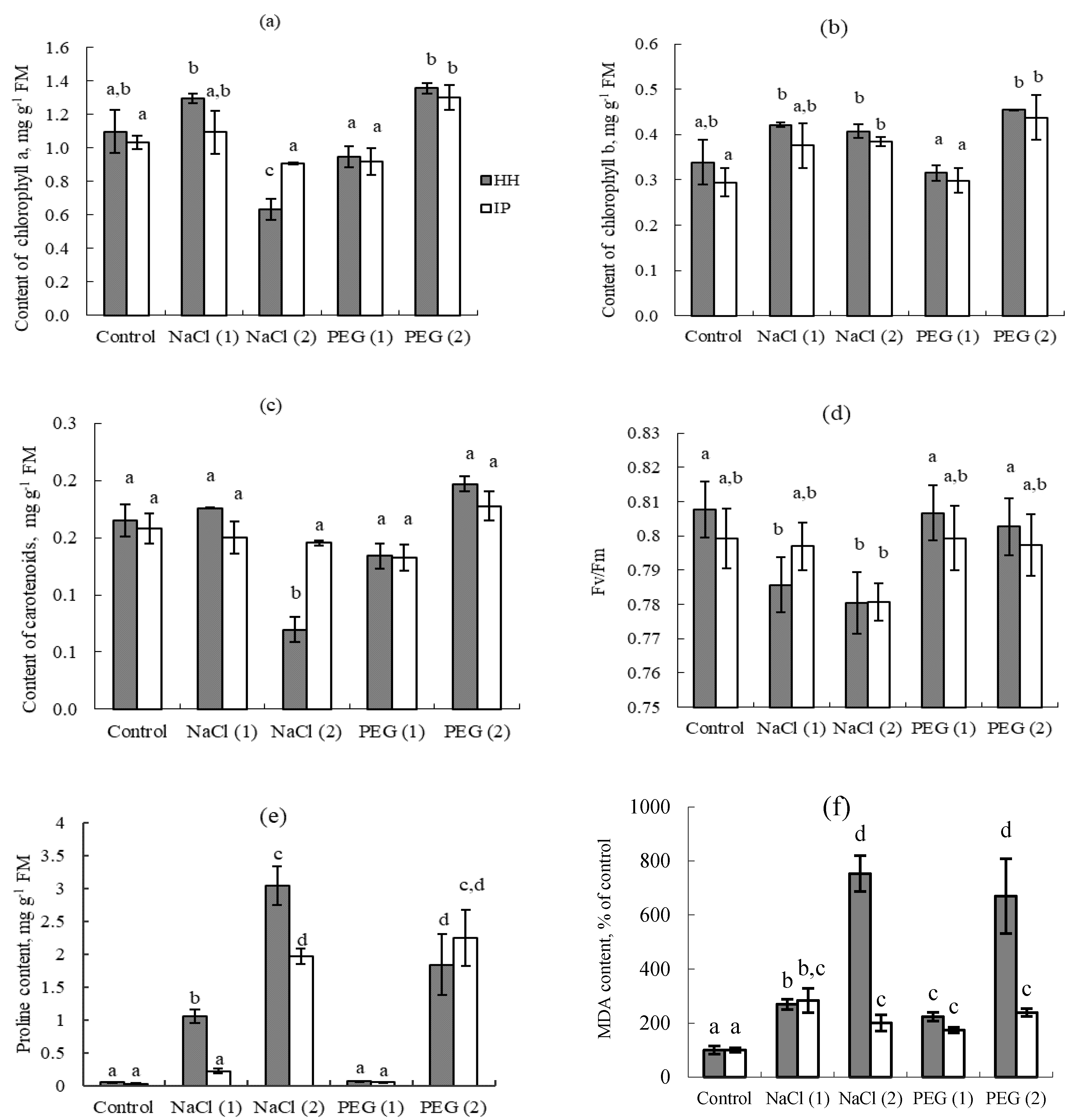
3.3. Model Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Massino, A.I.; Edenbaev, D.; Khujanazarov, T.M.; Azizov, K.; Boboev, F.; Shuyskaya, E.V.; Massino, I.V.; Toderich, K.N. Comparative performance of corn, sorghum and pearl millet growing under saline soil and water environments in Aral Sea Basin. J. Arid Land Stud. 2015, 25, 269–272. [Google Scholar]
- Rao, N.K.; McCann, I.; Shahid, S.A.; Butt, K.U.; Al Araj, B.; Ismail, S. Sustainable use of salt-degraded and abandoned farms for forage production using halophytic grasses. Crop Pasture Sci. 2017, 68, 483–492. [Google Scholar] [CrossRef]
- Baltensperger, D.D. Progress with Proso, Pearl and Other Millets Reprinted from: Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002. [Google Scholar]
- Toderich, K.; Khalikulov, Z.; Popova, V.; Boboev, F.; Azizov, K.; Rafiev, B.; Akinshina, N.; Yuldashev, T.; Kuliev, T.; Kurbanbaev, A.; et al. Sorghum and Pearl Millet for Crop Diversification, Improved Crop-Livestock Productivity and Farmers’ Livelihood in Central Asia. ICBA. 2013, p. 4. Available online: www.cac-program.org/download/file/93 (accessed on 4 August 2018).
- Rose, D.J.; Santra, D.K. Proso millet (Panicum miliaceum L.) fermentation for fuel ethanol production. Ind. Crop. Prod. 2013, 43, 602–605. [Google Scholar] [CrossRef]
- Gupta, S.K.; Rai, K.N.; Singh, P.; Ameta, V.L.; Gupta, S.K.; Jayalekha, A.K.; Mahala, R.S.; Pareek, S.; Swami, M.L.; Verma, Y.S. Seed set variability under high temperatures during flowering period in pearl millet (Pennisetum glaucum L. (R.) Br.). Field Crop. Res. 2015, 171, 41–53. [Google Scholar] [CrossRef]
- Lyon, D.J.; Burgener, P.A.; DeBoer, K.L.; Harveson, R.M.; Hein, G.L.; Hergert, G.W.; Holman, T.L.; Nelson, L.A.; Johnson, J.J.; Nleya, T.; et al. Proso Millet in the Great Plains; Publication # EC137; University of Nebraska Extension Serv.: Lincoln, NB, USA, 2008. [Google Scholar]
- Borde, M.; Dudhane, M.; Jite, P. Growth photosynthetic activity and antioxidant responses of mycorrhizal and non-mycorrhizal bajra (Pennisetum glaucum) crop under salinity stress condition. Crop Prot. 2011, 30, 265–271. [Google Scholar] [CrossRef]
- Sneha, S.; Rishi, A.; Chandra, S. Effect of short term salt stress on chlorophyll content, protein and activities of catalase and ascorbate peroxidase enzymes in pearl millet. Am. J. Plant Physiol. 2014, 9, 32–37. [Google Scholar] [CrossRef]
- Ali, S.A.M.; Idris, A.Y. Germination and Seedling Growth of Pearl Millet (Pennisetum glaucum L.) Cultivars under Salinity Conditions. Int. J. Plant Sci. Ecol. 2015, 1, 1–5. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Mokhberdoran, F.; Nabavi Kalat, S.M.; Sadrabadi, R. Effect of temperature, iso-osmotic concentrations of NaCl and PEG agents on germination and some seedling growth yield components in rice (Oryza sativa L.). Asian J. Plant Sci. 2009, 8, 409. [Google Scholar] [CrossRef]
- Patade, V.Y.; Bhargava, S.; Suprasanna, P. Effects of NaCl and iso-osmotic PEG stress on growth, osmolytes accumulation and antioxidant defense in cultured sugarcane cells. Plant Cell Tissue Organ Cult. 2012, 108, 279–286. [Google Scholar] [CrossRef]
- Farsiani, A.; Ghobadi, M.E. Effects of PEG and NaCl stress on two cultivars of corn (Zea mays L.) at germination and early seedling stages. World Acad. Sci. Eng. Technol. 2009, 57, 382–385. [Google Scholar]
- Gholamin, R.; Khayatnezhad, M. Effects of polyethylene glycol and NaCl stress on two cultivars of wheat (Triticum durum) at germination and early seedling stages. Am.-Eurasian J. Agric. Environ. Sci. 2010, 9, 86–90. [Google Scholar]
- Eshghizaden, H.R.; Kafi, M.; Nezami, A. The mechanisms of salinity tolerance in the xero-halophyte Blue Panicgrass (Panicum antidotale Retz). Notulae Sci. Biol. 2012, 4, 59–64. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Sudhir, P.; Murthy, S.D.S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Murata, N. Salt stress inhibits photosystems II and I in cyanobacteria. Photosynth. Res. 2008, 98, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, U.; Robredo, A.; Lacuesta, M.; Sgherri, C.; Muñoz-Rueda, A.; Navari-Izzo, F.; Mena-Petite, A. The oxidative stress caused by salinity in two barley cultivars is mitigated by elevated CO2. Physiol. Plant 2009, 135, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Q.; Zhang, Y.; Cui, J.; Chen, G.; Xie, B.; Wu, C.; Liu, H. Synergistic and antagonistic effects of salinity and pH on germination in switchgrass (Panicum virgatum L.). PLoS ONE 2014, 9, e85282. [Google Scholar] [CrossRef] [PubMed]
- Sulas, L.; Re, G.A.; Bullitta, S.; Piluzza, G. Chemical and productive properties of two Sardinian milk thistle (Silybum marianum (L.) Gaertn.) populations as sources of nutrients and antioxidants. Genet. Resour. Crop Evol. 2016, 63, 315–326. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Pasker, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 180–198. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzymol. 1987, 148, 350–382. [Google Scholar]
- Schreiber, U. Chlorophyll Fluorescence and Photosynthetic Energy Conversion: Simple Introductory Experiments with the TEACHING-PAM Chlorophyll Fluorometer; Heinz Walz GmbH: Effeltrich, Germany, 1997; p. 73. [Google Scholar]
- Orlovsky, N.S.; Japakova, U.N.; Zhang, H.F.; Volis, S. Effect of salinity on seed germination, growth and ion content in dimorphic seeds of Salicornia europaea L. (Chenopodiaceae). Plant Divers. 2016, 38, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2015, 92, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Khairwal, I.S.; Rai, K.N.; Diwakar, B.; Sharma, Y.K.; Rajpurohit, B.S.; Nirwan, B.; Bhattacharjee, R. Pearl Millet Crop Management and Seed Production; Anual Manual; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, Andhra Pradesh, India, 2007. [Google Scholar]
- Al-Dakheel, A.J.; Iftikhar, M. Hussain Genotypic Variation for Salinity Tolerance in Cenchrus ciliaris L. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed]
- Aslam, R.; Bostan, N.; Nabgha-e-Amen Maria, M.; Safdar, W. A critical review on halophytes: Salt tolerant plants. J. Med. Plants Res. 2011, 5, 7108–7118. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pearl Millet Lines | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Hashaki 1 | HHVBC Tall | IP 19586 | GB 8735 | Sudan POP I | JBV 2 | ICMV 155 | Raj 171 | IP 13150 | IP 22269 | ICMS 7704 | |
| Low salinity | |||||||||||
| Period of vegetation (days) | 90 ± 1.6 d | 97 ± 1.9 c | 113 ± 2.3 a,b | 97 ± 2.9 c | 100 ± 1.8 b,c | 103 ± 3.2 b | 103 ± 2.6 b | 103 ± 3.6 b | 118 ± 2.6 a | 115 ± 1.1 a | 110 ± 3.5 b |
| Plant density (1000 plants ha−1) | 148.4 ± 33.4 a,b | 168.8 ± 27.7 a | 137.9 ± 13.5 a,b | 160.8 ± 26.7 | 177.8 ± 30.8 a | 137.8 ± 27.7 a,b | 146.6 ± 23.1 a,b | 166.6 ± 10.2 a | 110.2 ± 18.9 a | 191.2 ± 13.3 a | 152.8 ± 24.8 a,b |
| Field germination (%) | 50.2 ± 0.52 b | 59.8 ± 0.60 a | 58.5 ± 1.91 a | 57.6 ± 2.66 a | 61.0 ± 2.62 a | 59.6 ± 2.81 a | 53.8 ± 2.60 b | 59.8 ± 1.68 a | 57.3 ± 0.98 a | 53.1 ± 0.45 b | 59.5 ± 0.79 a |
| Plant height (m) | 2.02 ± 0.20 a | 2.47 ± 0.10 a | 2.54 ± 0.23 a | 1.72 ± 0.07 b | 2.04 ± 0.21 a | 2.32 ± 0.36 a | 2.23 ± 0.03 a | 2.24 ± 0.44 a | 2.37 ± 0.45 a | 2.51 ± 0.52 a | 2.19 ± 0.15 a |
| Green biomass (t ha−1) | 27.7 ± 3.1 d | 43.60 ± 2.9 c | 62.26 ± 5.3 a | 50.7 ± 4.6 b | 40.2 ± 3.4 c | 47.4 ± 5.1 b | 36.6 ± 4.0 c | 48.9 ± 2.7 b | 36.4 ± 2.1 c | 44.8 ± 2.7 c | 49.4 ± 2.8 b |
| Dry biomass (t ha−1) | 11.1 ± 1.7 d | 16.1 ± 1.4 c | 27.0 ± 1.7 a | 20.3 ± 1.4 b | 13.8 ± 1.2 c | 20.1 ± 0.7 b | 12.8 ± 2.5 c | 18.4 ± 0.4 b | 15.1 ± 1.9 c | 17.4 ± 1.9 c | 15.0 ± 2.6 c |
| Grain yields (t ha−1) | 2.51 ± 0.46 b | 4.37 ± 0.57 a | 2.14 ± 0.29 b,c | 2.85 ± 0.41 b | 2.93 ± 0.31 b | 2.88 ± 0.56 b | 1.89 ± 0.32 c | 2.84 ± 0.23 b | 0.79 ± 0.18 d | 1.17 ± 0.37 c,d | 2.75 ± 0.42 b |
| Medium salinity | |||||||||||
| Period of vegetation (days) | 93.2 ± 1.9 d | 102.4 ± 2.6 c | 121.4 ± 1.5 b | 109.4 ± 2.2 c | 136.3 ± 2.8 a | 105.4 ± 1.4 c | 103.3 ± 0.8 c | 116.5 ± 0.9 b | 117.0 ± 2.4 b | 136.1 ± 0.9 a | 133.4 ± 1.5 a |
| Plant density (1000 plants ha−1) | 133.9 ± 5.4 b | 159.1 ± 4.6 a | 155.6 ± 5.3 a | 152.5 ± 5.9 a | 162.5 ± 5.6 a | 152.2 ± 6.0 a | 143.8 ± 14.3 a,b | 157.8 ± 10.2 a | 110.2 ± 9.2 c | 160.0 ± 13.3 a | 148.4 ± 7.8 a |
| Field germination (%) | 37.1 ± 8.4 a | 42.2 ± 6.9 a | 36.7 ± 3.4 a | 40.2 ± 6.7 a | 44.4 ± 7.7 a | 34.4 ± 6.9 a | 36.7 ± 5.8 a | 39.4 ± 2.5 a | 27.6 ± 4.8 b | 40.0 ± 3.3 a | 37.1 ± 6.2 a |
| Plant height (m) | 2.29 ± 0.09 b | 2.35 ± 0.06 b | 2.61 ± 0.10 a | 2.32 ± 0.11 b | 2.55 ± 0.12 a | 2.57 ± 0.10 a | 2.35 ± 0.06 a,b | 2.37 ± 0.10 a | 2.65 ± 0.07 a | 2.45 ± 0.18 a | 2.17 ± 0.09 c |
| Green biomass (t ha−1) | 25.4 ± 0.8 d | 40.4 ± 1.2 b | 50.5 ± 0.8 a | 42.6 ± 1.8 b | 37.4 ± 3.6 b,c | 26.7 ± 1.7 d | 25.1 ± 1.7 d | 44.9 ± 1.3 b | 32.9 ± 1.7 c | 36.4 ± 0.7 c | 39.0 ± 1.2 b,c |
| Dry biomass (t ha−1) | 7.3 ± 0.7 c | 12.8 ± 1.2 b | 17.0 ± 1.1 a | 10.7 ± 0.7 b | 10.3 ± 1.8 b | 7.1 ± 1.8 c | 7.5 ± 0.8 c | 10.7 ± 0.9 b | 10.1 ± 0.9 b | 9.5 ± 0.5 b | 8.4 ± 0.7 b,c |
| Grain yields (t ha−1) | 1.29 ± 0.07 b | 2.23 ± 0.09 a | 1.38 ± 0.15 b | 1.51 ± 0.08 b | 1.15 ± 0.21 b,c | 1.30 ± 0.10 b | 1.12 ± 0.15 b,c | 1.01 ± 0.20 c | 1.41 ± 0.09 b | 0.83 ± 0.07 c | 0.92 ± 0.09 c |
| Source | Green Biomass | Grain Yields | ||||
|---|---|---|---|---|---|---|
| df | F | Sig. | df | F | Sig. | |
| Genotype | 10 | 44.631 | <0.0001 | 10 | 18.041 | <0.0001 |
| Salinity level | 2 | 132.168 | <0.0001 | 2 | 223.707 | <0.0001 |
| Genotype and salinity level | 10 | 7.008 | <0.0001 | 10 | 9.351 | <0.0001 |
| Error | 44 | 44 | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toderich, K.; Shuyskaya, E.; Rakhmankulova, Z.; Bukarev, R.; Khujanazarov, T.; Zhapaev, R.; Ismail, S.; Gupta, S.K.; Yamanaka, N.; Boboev, F. Threshold Tolerance of New Genotypes of Pennisetum glaucum (L.) R. Br. to Salinity and Drought. Agronomy 2018, 8, 230. https://doi.org/10.3390/agronomy8100230
Toderich K, Shuyskaya E, Rakhmankulova Z, Bukarev R, Khujanazarov T, Zhapaev R, Ismail S, Gupta SK, Yamanaka N, Boboev F. Threshold Tolerance of New Genotypes of Pennisetum glaucum (L.) R. Br. to Salinity and Drought. Agronomy. 2018; 8(10):230. https://doi.org/10.3390/agronomy8100230
Chicago/Turabian StyleToderich, Kristina, Elena Shuyskaya, Zulfira Rakhmankulova, Roman Bukarev, Temur Khujanazarov, Rauan Zhapaev, Shoaib Ismail, Shashi K. Gupta, Norikazu Yamanaka, and Fazyl Boboev. 2018. "Threshold Tolerance of New Genotypes of Pennisetum glaucum (L.) R. Br. to Salinity and Drought" Agronomy 8, no. 10: 230. https://doi.org/10.3390/agronomy8100230
APA StyleToderich, K., Shuyskaya, E., Rakhmankulova, Z., Bukarev, R., Khujanazarov, T., Zhapaev, R., Ismail, S., Gupta, S. K., Yamanaka, N., & Boboev, F. (2018). Threshold Tolerance of New Genotypes of Pennisetum glaucum (L.) R. Br. to Salinity and Drought. Agronomy, 8(10), 230. https://doi.org/10.3390/agronomy8100230