
Opportunities for Napier Grass (Pennisetum purpureum) Improvement Using Molecular Genetics


Abstract
:1. Introduction
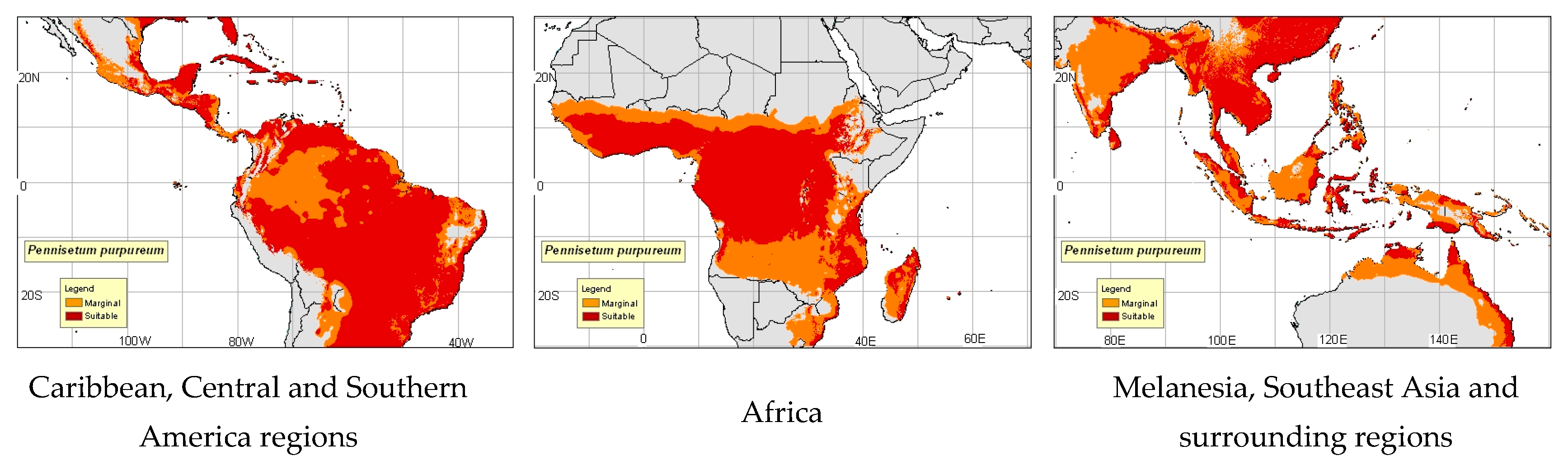
1.1. Origin, Propagation and Distribution
1.2. Economic Importance
1.3. Genetic Resources, Molecular Diversity and Breeding
2. Current Status
2.1. Yield and Morphology
2.2. Nutritional Qualities
2.3. Water Use Efficiency
2.4. Pests and Diseases
2.4.1. Viral Diseases
2.4.2. Fungal Diseases
2.4.3. Phytoplasma (Stunt) Disease
3. Future Prospects
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ADF | Acid Detergent Fibre |
| AFLP | Amplified Fragment Length Polymorphism |
| Bt | Bacillus thuringiensis |
| CIAT | International Center for Tropical Agriculture |
| CP | Crude Protein |
| CSIRO | Commonwealth Scientific and Industrial Research Organization |
| DM | dry Matter |
| DRC | Democratic Republic of Congo |
| EST | Expressed Sequence Tag |
| GBS | Genotyping by Sequencing |
| GRIN | Germplasm Resources Information Network |
| ha | hectare |
| icipe | International Centre of Insect Physiology and Ecology |
| ILRI | International Livestock Research Institute |
| ISSR | Inter-Sequential Simple Repeat |
| KALRO | Kenya Agricultural and Livestock Research Organization |
| L | Litre |
| NBPGR | National Bureau of Plant Genetic Resources, Delhi, India |
| NDF | Neutral Detergent Fibre |
| RNAi | RNA Interference |
| RAPD | Random Amplification of Polymorphic DNA |
| RFLP | Restriction Fragment Length Polymorphism |
| RBG | Royal Botanic Gardens |
| SNPs | single-nucleotide polymorphisms |
| SRAPs | Sequence-Related Amplified Polymorphisms |
| SSR | Simple Sequence Repeat |
| USDA | United States Department of Agriculture |
References
- Lukuyu, B.A.; Gachuiri, C.K.; Lukuyu, M.N.; Lusweti, C.; Mwendia, S. Feeding Dairy Cattle in East Africa; East Africa Dairy Development Project: Nairobi, Kenya, 2012. [Google Scholar]
- Kabirizi, J.; Muyekho, F.; Mulaa, M.; Msangi, R.; Pallangyo, B.; Kawube, G.; Zziwa, E.; Mugerwa, S.; Ajanga, S.; Lukwago, G.; et al. Napier Grass Feed Resource: Production, Constraints and Implications For Smallholder Farmers in Eastern and Central Africa; The Eastern African Agricultural Productivity Project: Naivasha, Kenya, 2015. [Google Scholar]
- Farrell, G.; Simons, S.; Hillocks, R. Pests, diseases and weeds of Napier grass, Pennisetum purpureum: A review. Int. J. Pest Manag. 2002, 48, 39–48. [Google Scholar] [CrossRef]
- Lowe, A.J.; Thorpe, W.; Teale, A.; Hanson, J. Characterisation of germplasm accessions of Napier grass (Pennisetum purpureum and P. purpureum× P. glaucum hybrids) and comparison with farm clones using RAPD. Genet. Resour. Crop Evol. 2003, 50, 121–132. [Google Scholar] [CrossRef]
- Sandhu, J.S.; Kumar, D.; Yadav, V.K.; Singh, T.; Sah, R.P.; Radhakrishna, A. Recent trends in breeding of tropical grass and forage species. In Proceedings of the 23rd International Grassland Congress, New Delhi, India, 20–24 November 2015; Vijay, D., Srivastava, M.K., Gupta, C.K., Malaviya, D.R., Roy, M.M., Mahanta, S.K., Singh, J.B., Maity, A., Ghosh, P.K., Eds.; Range Management Society of India: Jhansi, India, 2015; pp. 337–348. [Google Scholar]
- Singh, B.P.; Singh, H.P.; Obeng, E. Elephant grass. In Biofuel Crops: Production, Physiology and Genetics; Singh, B.P., Ed.; CAB International: Fort Valley State University, Fort Valley, GA, USA, 2013; pp. 271–291. [Google Scholar]
- Brunken, J.N. A systematic study of Pennisetum sect. Pennisetum (Gramineae). Am. J. Bot. 1977, 64, 161–176. [Google Scholar] [CrossRef]
- Donadío, S.; Giussani, L.M.; Kellogg, E.A.; Zuolaga, F.O.; Morrone, O. A preliminary molecular phylogeny of Pennisetum and Cenchrus (Poaceae-Paniceae) based on the trnL-F, rpl16 chloroplast markers. Taxon 2009, 58, 392–404. [Google Scholar]
- Dos Reis, G.B.; Mesquita, A.T.; Torres, G.A.; Andrade-Vieira, L.F.; Vander Pereira, A.; Davide, L.C. Genomic homeology between Pennisetum purpureum and Pennisetum glaucum (Poaceae). Comp. Cytogenet. 2014, 8, 199. [Google Scholar] [PubMed]
- Martel, E.; De Nay, D.; Siljak-Yakoviev, S.; Brown, S.; Sarr, A. Genome size variation and basic chromosome number in Pearl millet and fourteen related Pennisetum species. J. Hered. 1997, 88, 139–143. [Google Scholar] [CrossRef]
- Harris, K.; Anderson, W.; Malik, R. Genetic relationships among Napier grass (Pennisetum purpureum Schum.) nursery accessions using AFLP markers. Plant Genet. Resour. 2010, 8, 63–70. [Google Scholar] [CrossRef]
- Kandel, R.; Singh, H.P.; Singh, B.P.; Harris-Shultz, K.R.; Anderson, W.F. Assessment of genetic diversity in Napier Grass (Pennisetum purpureum Schum.) using microsatellite, single-nucleotide polymorphism and insertion-deletion markers from Pearl Millet (Pennisetum glaucum (L.) R. Br.). Plant Mol. Biol. Rep. 2016, 34, 265–272. [Google Scholar] [CrossRef]
- Cook, B.G.; Pengelly, B.C.; Brown, S.D.; Donnelly, J.L.; Eagles, D.A.; Franco, M.A.; Hanson, J.; Mullen, B.F.; Partridge, I.J.; Peters, M.; et al. Tropical Forages: An Interactive Selection Tool; [CD-ROM], CSIRO, DPI&F(Qld), CIAT and ILRI: Brisbane, Australia, 2005. [Google Scholar]
- Diz, D.A. Breeding Procedures and Seed Production Management in Pearl millet × Elephant grass Hexaploids Hybrids. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 1994. [Google Scholar]
- Cheng, Y. Forage breeding in Taiwan-Review. Asian Australas. J. Anim.Sci. 1991, 4, 203–209. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G.; Giger-Reverdin, S.; Lebas, F. Elephant grass (Pennisetum purpureum). Available online: http://www.feedipedia.org/node/395 (accessed on 21 September 2016).
- Ishii, Y.; Hamano, K.; Kang, D.J.; Kannika, R.; Idota, S.; Fukuyama, K.; Nishiwaki, A. C4-Napier grass cultivation for cadmium phytoremediation activity and organic livestock farming in Kyushu, Japan. J. Agric. Sci. Technol. 2013, 3, 321. [Google Scholar]
- Orodho, A.B. The Role and Importance of Napier Grass in the Smallholder Dairy Industry in Kenya; Food and Agriculture Organization: Rome, Italy, 2006; p. 2011. [Google Scholar]
- Pandit, N.P.; Shrestha, M.K.; Yi, Y.; Diana, J.S.; Rampur, C. Polyculture of grass carp and Nile tilapia with Napier grass as the sole nutrient input in the subtropical climate of Nepal. In Proceedings of the 6th International Symposium on Tilapia in Aquaculture, Manila, Philippines, 12–16 September 2004; pp. 12–16. [Google Scholar]
- Shrestha, M.; Yadav, C. Feeding of Napier (Pennisetum purpureum) to grass carp in polyculture: A sustainable fish culture practice for small farmers. Asian Fish. Sci. 1998, 11, 287–294. [Google Scholar]
- Shaha, D.C.; Kundu, S.R.; Hasan, M.N. Production of organic grass carp (Ctenopharyngodon idella) and GIFT tilapia (Oreochromis niloticus) using Napier grass, Pennisetum purpureum. J. Fish. 2015, 3, 233–238. [Google Scholar] [CrossRef]
- Akah, N.; Onweluzo, J. Evaluation of water-soluble vitamins and optimum cooking time of fresh edible portions of Elephant Grass (Pennisetum purpureum L. Schumach) shoot. Niger. Food J. 2014, 32, 120–127. [Google Scholar] [CrossRef]
- Khan, Z.; Chiliswa, P.; Ampong-Nyarko, K.; Smart, L.; Polaszek, A.; Wandera, J.; Mulaa, M. Utilisation of wild gramineous plants for management of cereal stemborers in Africa. Int. J. Trop. Insect Sci. 1997, 17, 143–150. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.; Hutter, N.J.; Wilkins, R.M.; Wadhams, L.J. Assessment of the potential of Napier grass (Pennisetum purpureum) varieties as trap plants for management of Chilo partellus. Entomol. Exp. Appl. 2006, 119, 15–22. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.; Wadhams, L.J.; Pickett, J.A.; Mumuni, A. Evaluation of Napier grass (Pennisetum purpureum) varieties for use as trap plants for the management of African stemborer (Busseola fusca) in a push–pull strategy. Entomol. Exp. Appl. 2007, 124, 201–211. [Google Scholar] [CrossRef]
- Oliveira, M.L.F.; Daher, R.F.; Gravina, G.D.A.; da Silva, V.B.; Viana, A.P.; Rodrigues, E.V.; Shimoya, A.; Junior, A.T.D.A.; Menezes, B.R.D.S.; Rocha, A.D.S. Pre-breeding of Elephant grass for energy purposes and biomass analysis in Campos dos Goytacazes-RJ, Brazil. Afr. J. Agric. Res. 2014, 9, 2743–2758. [Google Scholar]
- Rengsirikul, K.; Ishii, Y.; Kangvansaichol, K.; Sripichitt, P.; Punsuvon, V.; Vaithanomsat, P.; Nakamanee, G.; Tudsri, S. Biomass yield, chemical composition and potential ethanol yields of eight cultivars of Napier grass (Pennisetum purpureum Schumach.) harvested 3-monthly in central Thailand. J. Sustain. Bioenergy Syst. 2013, 3, 107. [Google Scholar] [CrossRef]
- Lima, M.A.; Gomez, L.D.; Steele-King, C.G.; Simister, R.; Bernardinelli, O.D.; Carvalho, M.A.; Rezende, C.A.; Labate, C.A.; McQueen-Mason, S.J.; Polikarpov, I. Evaluating the composition and processing potential of novel sources of Brazilian biomass for sustainable biorenewables production. Biotechnol. Biofuels 2014, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Hanna, W.W.; Monson, W.G. Yield, quality, and breeding of Pearl millet × Napier grass interspecific hybrids. Agron. J. 1980, 72, 358–360. [Google Scholar] [CrossRef]
- Pitman, W.D.; Sotomayor-Rios, A. Tropical Forage Plants: Development and Use; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Anderson, W.F.; Casler, M.D.; Baldwin, B.S. Improvement of perennial forage species as feedstock for bioenergy. In Genetic Improvement of Bioenergy Crops; Vermerris, W., Ed.; Springer Science + Business Media LLC: New York, NY, USA, 2008; pp. 347–376. [Google Scholar]
- Paiva, E.A.; Bustamante, F.O.; Barbosa, S.; Pereira, A.V.; Davide, L.C. Meiotic behavior in early and recent duplicated hexaploid hybrids of Napier grass (Pennisetum purpureum) and Pearl millet (Pennisetum glaucum). Caryologia 2012, 65, 114–120. [Google Scholar] [CrossRef]
- Omayio, D.O.; Ajanga, S.I.; Muoma, J.V.; Muyekho, F.N.; Yamame, M.K.; Kariuki, I.M.S. Using Napier grass accessions’ origins, neighbour joining groups and their response to Ustilago kamerunensis to predict a probable co-evolutionary scenario. Int. J. Recent Sci. Res. 2015, 6, 2639–2645. [Google Scholar]
- Kawube, G.; Alicai, T.; Otim, M.; Mukwaya, A.; Kabirizi, J.; Talwana, H. Resistance of Napier grass clones to Napier grass stunt Disease. Afr. Crop Sci. J. 2014, 22, 229–236. [Google Scholar]
- Hanna, W.W.; Monson, W.G. Registration of dwarf Tift N75 Napier grass germplasm. Crop Sci. 1988, 28, 870–871. [Google Scholar] [CrossRef]
- Premaratne, S.; Premalal, G.G.C. Hybrid Napier (Pennisetum perpureum × Pennisetum americarnum) var. CO-3: A resourceful fodder grass for dairy development in Sri Lanka. J. Agric. Sci. 2006, 2. [Google Scholar] [CrossRef]
- Genesys. Available online: https://www.genesys-pgr.org/welcome (acessed on 30 November 2016).
- Tcacenco, F.A.; Lance, G.N. Selection of morphological traits for characterisation of Elephant grass accessions. Trop. Grassl. 1992, 26, 145–155. [Google Scholar]
- Van De Wouw, M.; Hanson, J.; Leuthi, S. Morphological and argonomic characterisation of a collection of Napier grass (Pennisetum purpureum) and P. purpureum × P. glaucum. Trop. Grassl. 1999, 33, 150–158. [Google Scholar]
- Smith, R.L.; Schweder, M.; Chowdhury, M.; Seib, J.; Schank, S. Development and application of RFLP and RAPD DNA markers in genetic improvement of Pennisetum for biomass and forage production. Biomass Bioenergy 1993, 5, 51–62. [Google Scholar] [CrossRef]
- Babu, C.; Sundaramoorthi, J.; Vijayakumar, G.; Ram, S.G. Analysis of genetic diversity in Napier grass (Pennisetum purpureum Schum) as detected by RAPD and ISSR markers. J. Plant Biochem. Biotechnol. 2009, 18, 181–187. [Google Scholar] [CrossRef]
- Bhandari, A.P.; Sukanya, D.; Ramesh, C. Application of isozyme data in fingerprinting Napier grass (Pennisetum purpureum Schum.) for germplasm management. Genet. Resour. Crop Evol. 2006, 53, 253–264. [Google Scholar] [CrossRef]
- Struwig, M.; Mienie, C.; Van Den Berg, J.; Mucina, L.; Buys, M. AFLPs are incompatible with RAPD and morphological data in Pennisetum purpureum (Napier grass). Biochem. Syst. Ecol. 2009, 37, 645–652. [Google Scholar] [CrossRef]
- Wanjala, B.W.; Obonyo, M.; Wachira, F.N.; Muchugi, A.; Mulaa, M.; Harvey, J.; Skilton, R.A.; Proud, J.; Hanson, J. Genetic diversity in Napier grass (Pennisetum purpureum) cultivars: Implications for breeding and conservation. AoB Plants 2013, 5, plt022. [Google Scholar] [CrossRef] [PubMed]
- De Lima, R.; Daher, R.; Goncalves, L.; Rossi, D.; do Amaral Júnior, A.; Pereira, M.; Lédo, F. RAPD and ISSR markers in the evaluation of genetic divergence among accessions of Elephant grass. Genet. Mol. Res. 2011, 10, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.-M.; Zhou, F.; Zhang, X.-Q.; Zhang, J.-M. Genetic variability and relationship between MT-1 Elephant grass and closely related cultivars assessed by SRAP markers. J. Genet. 2009, 88, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Allouis, S.; Qi, X.; Lindup, S.; Gale, M.; Devos, K. Construction of a BAC library of pearl millet, Pennisetum glaucum. Theor. Appl. Genet. 2001, 102, 1200–1205. [Google Scholar] [CrossRef]
- Budak, H.; Pedraza, F.; Cregan, P.; Baenziger, P.; Dweikat, I. Development and utilization of SSRs to estimate the degree of genetic relationships in a collection of pearl millet germplasm. Crop Sci. 2003, 43, 2284–2290. [Google Scholar] [CrossRef]
- Mariac, C.; Luong, V.; Kapran, I.; Mamadou, A.; Sagnard, F.; Deu, M.; Chantereau, J.; Gerard, B.; Ndjeunga, J.; Bezançon, G. Diversity of wild and cultivated pearl millet accessions (Pennisetum glaucum (L.) R. Br.) in Niger assessed by microsatellite markers. Theor. Appl. Genet. 2006, 114, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Lindup, S.; Pittaway, T.; Allouis, S.; Gale, M.; Devos, K. Development of simple sequence repeat markers from bacterial artificial chromosomes without subcloning. Biotechniques 2001, 31, 358–362. [Google Scholar]
- Senthilvel, S.; Jayashree, B.; Mahalakshmi, V.; Kumar, P.S.; Nakka, S.; Nepolean, T.; Hash, C. Development and mapping of simple sequence repeat markers for pearl millet from data mining of expressed sequence tags. BMC Plant Biol. 2008, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Sousa Azevedo, A.L.; Costa, P.P.; Machado, J.C.; Machado, M.A.; Pereira, A.V.; José da Silva Lédo, F. Cross species amplification of microsatellite markers in and genetic diversity of Napier grass accessions. Crop Sci. 2012, 52, 1776–1785. [Google Scholar] [CrossRef]
- Dowling, C.D.; Burson, B.L.; Foster, J.L.; Tarpley, L.; Jessup, R.W. Confirmation of Pearl millet-Napier grass hybrids using EST-derived simple sequence repeat (SSR) markers. Am. J. Plant Sci. 2013, 4, 1004–1012. [Google Scholar] [CrossRef]
- Kawube, G.; Alicai, T.; Wanjala, B.; Njahira, M.; Awalla, J.; Skilton, R. Genetic diversity in Napier Grass (Pennisetum purpureum) assessed by SSR Markers. J. Agric. Sci. 2015, 7, 147. [Google Scholar] [CrossRef]
- Morais, R.F.D.; Souza, B.J.D.; Leite, J.M.; Soares, L.H.D.B.; Alves, B.J.R.; Boddey, R.M.; Urquiaga, S. Elephant grass genotypes for bioenergy production by direct biomass combustion. Pesqui. Agropecu. Bras. 2009, 44, 133–140. [Google Scholar] [CrossRef]
- Wangchuk, K.; Rai, K.; Nirola, H.; Dendup, C.; Mongar, D. Forage growth, yield and quality responses of Napier hybrid grass cultivars to three cutting intervals in the Himalayan foothills. Trop. Grassl. Forrajes Trop. 2015, 3, 142–150. [Google Scholar] [CrossRef]
- Lee, C.N.; Fukumoto, G.K.; Thorne, M.S.; Stevenson, M.H.; Nakahata, M.; Ogoshi, R.M. Bana Grass (Pennisetum purpureum): A Possible Forage for Ruminants in Hawai‘i; University of Hawai‘i: Honolulu, HI, USA, 2016. [Google Scholar]
- Magcale-Macandog, D.; Predo, C.; Menz, K.; Predo, A. Napier grass strips and livestock: A bioeconomic analysis. Agrofor. Syst. 1998, 40, 41–58. [Google Scholar] [CrossRef]
- Van den Berg, J.; Van Hamburg, H. Trap cropping with Napier grass, Pennisetum purpureum (Schumach), decreases damage by maize stem borers. Int. J. Pest Manag. 2015, 61, 73–79. [Google Scholar] [CrossRef]
- Tsai, W.-T.; Tsai, Y.-L. Thermochemical characterization of Napier grass as an energy source and its environmental and economic benefit analysis. Energy Sources Part B Econ. Plan. Policy 2016, 11, 130–136. [Google Scholar]
- Ra, K.; Shiotsu, F.; Abe, J.; Morita, S. Biomass yield and nitrogen use efficiency of cellulosic energy crops for ethanol production. Biomass Bioenergy 2012, 37, 330–334. [Google Scholar] [CrossRef]
- Mwendia, S.; Yunusa, I.; Whalley, R.; Sindel, B.; Kenney, D.; Kariuki, I. Use of plant water relations to assess forage quality and growth for two cultivars of Napier grass (Pennisetum purpureum) subjected to different levels of soil water supply and temperature regimes. Crop Pasture Sci. 2014, 64, 1008–1019. [Google Scholar]
- Williams, M.J.; Hanna, W.W. Performance and nutritive quality of dwarf and semi-dwarf Elephant grass genotypes in the south-eastern USA. Trop. Grassl. 1995, 29, 122–127. [Google Scholar]
- Halim, R.A.; Shampazuraini, S.; Idris, A.B. Yield and nutritive quality of nine Napier grass varieties in Malaysia. Malays. J. Anim. Sci. 2013, 16, 37–44. [Google Scholar]
- Mukhtar, M.; Ishii, Y.; Tudsri, S.; Idota, S.; Sonoda, T. Dry matter productivity and overwintering ability of the dwarf and normal Napier grasses as affected by the planting density and cutting frequency. Plant Prod. Sci. 2003, 6, 65–73. [Google Scholar] [CrossRef]
- Utamy, R.F.; Ishii, Y.; Idota, S.; Harada, N.; Fukuyama, K. Adaptability of dwarf Napier grass under cut and carry and grazing sysytems for smallholder beef farmers in southern Kyhushu, Japan. J. Warm Reg. Soc. Anim. Jpn. 2011, 54, 87–98. [Google Scholar]
- Kebede, G.; Feyissa, F.; Assefa, G.; Alemayehu, M.; Mengistu, A.; Kehaliew, A.; Melese, K.; Mengistu, S.; Tadesse, E.; Wolde, S. Chemical composition and in vitro organic matter digestibility of Napier Grass (Pennisetum purpureum (L.) Schumach) accessions in the mid and highland areas of Ethiopia. Int. J. Livest. Res. 2016, 6, 41–59. [Google Scholar] [CrossRef]
- Relwani, L.L.; Nakat, R.V.; Kandale, D.Y. Intercropping of four leuceana cultivars with three grasses. Leuceana Res. Rep. 1982, 3, 41. [Google Scholar]
- Amin, R.; Ali, N.R.S.M.Y.; Abul, H.M.; Khatun, M. Study on cutting intervals on biomass yield, nutritive value and their oxalate content of different high yielding Napier (P. purpureum) cultivars. Asian Australas. J. Biosci. Biotechnol. 2016, 1, 100–107. [Google Scholar]
- Zewdu, T. Variation in growth, yield, chemical composition and in vitro dry matter digestibility of Napier grass accessions (Pennisetum purpureum). Trop. Sci. 2005, 45, 67–73. [Google Scholar] [CrossRef]
- Nyambati, E.M.; Muyekho, F.N.; Onginjo, E.; Lusweti, C.M. Production, characterization and nutritional quality of Napier grass (Pennisetum purpureum (Schum.)) cultivars in Western Kenya. Afr. J. Plant Sci. 2010, 4, 496–502. [Google Scholar]
- Ortega-Gómez, R.; Castillo-Gallegos, E.; Jarillo-Rodríguez, J.; Escobar-Hernández, R.; Ocaña-Zavaleta, E.; de la Mora, B.V. Nutritive quality of ten grasses during the rainy season in a hot-humid climate and ultisol soil. Trop. Subtrop. Agroecosyst. 2011, 13, 481–491. [Google Scholar]
- Tavirimirwa, B.; Manzungu, E.; Ncube, S. The evaluation of dry season nutritive value of dominant and improved grasses in fallows in Chivi district, Zimbabwe. Online J. Anim. Feed Res. 2012, 2, 470–474. [Google Scholar]
- Gwayumba, W.; Christensen, D.; McKinnon, J.; Yu, P. Dry matter intake, digestibility and milk yield by Friesian cows fed two Napier grass varieties. Asian Australas. J. Anim. Sci. 2002, 15, 516–521. [Google Scholar] [CrossRef]
- Islam, M.; Saha, C.; Sarker, N.; Jalil, M.; Hasanuzzaman, M. Effect of variety on proportion of botanical fractions and nutritive value of different Napier grass (Pennisetum purpureum) and relationship between botanical fractions and nutritive Value. Asian Australas. J. Anim. Sci. 2003, 16, 837–842. [Google Scholar] [CrossRef]
- Keba, H.T.; Madakadze, I.C.; Angassa, A.; Hassen, A. Nutritive value of grasses in semi-arid rangelands of Ethiopia: Local experience based herbage preference evaluation versus laboratory analysis. Asian Australas. J. Anim. Sci. 2013, 26, 366. [Google Scholar] [CrossRef] [PubMed]
- Tessema, Z.K.; De Boer, W.F.; Baars, R.M.T.; Prins, H.H.T. Changes in soil nutrients, vegetation structure and herbaceous biomass in response to grazing in a semi-arid savanna of Ethiopia. J. Arid Environ. 2011, 75, 662–670. [Google Scholar] [CrossRef]
- Henkin, Z.; Ungar, E.D.; Dvash, L.; Perevolotsky, A.; Yehuda, Y.; Sternberg, M.; Voet, H.; Landau, S.Y. Effects of cattle grazing on herbage quality in a herbaceous Mediterranean rangeland. Grass Forage Sci. 2011, 66, 516–525. [Google Scholar] [CrossRef]
- Van der Westhuizen, H.; Snyman, H.; Fouché, H. A degradation gradient for the assessment of rangeland condition of a semi-arid sourveld in southern Africa. Afr. J. Range Forage Sci. 2005, 22, 47–58. [Google Scholar] [CrossRef]
- Okwori, A.I.; Magani, I.E. Influence of nitrogen sources and cutting interval on the digestibility of four grass species in the southern guinea savanna of Nigeria. Agric. Biol. J. N. Am. 2010, 1, 526–533. [Google Scholar]
- Hasyim, H.; Ishii, Y.; Ahmad, W.; Sachiko, I. Quality herbage production of dwarf Napier grass with Italian Ryegrass cropping under digested effluent application in Southern Kyushu, Japan. Am. J. Agric. Biol. Sci. 2015, 11, 35–44. [Google Scholar] [CrossRef]
- Na, C.I.; Fedenko, J.R.; Sollenberger, L.E.; Erickson, J.E. Harvest management affects biomass composition responses of C4 perennial bioenergy grasses in the humid subtropical USA. GCB Bioenergy 2016. [Google Scholar] [CrossRef]
- Butt, N.M.; Donart, G.B.; Southwara, M.G.; Pieper, R.D. Effect of defoliation on plant growth of Napier grass. Trop. Sci. Lond. 1993, 33, 111. [Google Scholar]
- Bayble, T.; Melaku, S.; Prasad, N. Effects of cutting dates on nutritive value of Napier (Pennisetum purpureum) grass planted sole and in association with Desmodium (Desmodium intortum) or Lablab (Lablab purpureus). Livest. Res. Rural Dev. 2007, 19, 120–136. [Google Scholar]
- Carvalho, C.A.B.D.; Menezes, J.B.D.O.X.D.; Cóser, A.C. Effect of fertilizer and cutting frequency on yield and nutritive value of Elephant grass. Cienc. Agrotecnol. 2000, 24, 233–241. [Google Scholar]
- Mirza, S.N.; Muhammad, N.; Qamar, I.A. Effect of growth stages on the yield and quality of forage grasses. Pak. J. Agric. Res. 2002, 17, 145–147. [Google Scholar]
- Boudet, A.M. Towards an understanding of the supramolecular organization of the lignified wall. Plant Cell Wall 2003, 8, 155–182. [Google Scholar]
- Jung, H.-J.G. Forage digestibility: The intersection of cell wall lignification and plant tissue anatomy. In Proceedings of the 23rd Annual Florida Ruminant Nutrition Symposium, Gainesville, FL, USA, 31 January–1 February 2012; University of Florida: Gainesville, FL, USA, 2012; pp. 162–174. [Google Scholar]
- Moore, K.J.; Jung, H.-J.G. Lignin and fiber digestion. J. Range Manag. 2001, 54, 420–430. [Google Scholar] [CrossRef]
- Agbor, V.B.; Cicek, N.; Sparling, R.; Berlin, A.; Levin, D.B. Biomass pretreatment: Fundamentals toward application. Biotechnol. Adv. 2011, 29, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Ripley, B.; Woodward, F.; Osborne, C. Drought limitation of photosynthesis differs between C3 and C4 grass species in a comparative experiment. Plant Cell Environ. 2011, 34, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.A.; Pineda, M.; de la Cruz Jiménez, J.; Vergara, M.F.; Rao, I.M. Contrasting strategies to cope with drought conditions by two tropical forage C4 grasses. AoB Plants 2015, 7, plv107. [Google Scholar] [CrossRef] [PubMed]
- Purbajanti, E.; Anwar, S.; Wydiati, F.K. Drought stress effect on morphology characters, water use efficiency, growth and yield of guinea and napier grasses. Int. Res. J. Plant Sci. 2012, 3, 47. [Google Scholar]
- Rott, P.; Chatenet, M.; Granier, M.; Baudin, P. L’échaudure des feuilles de canne à sucre provoquée par Xanthomonas albilineans (Ashby) Dowson. II: Diagnostic et spectres d'hôtes de l'agent pathogène en Afrique tropicale. L'Agron. Trop. 1988, 43, 244–251. [Google Scholar]
- Farrell, G. Towards the Management of Ustilago kamerunensis H Sydow and Sydow, a Smut Pathogen of Napier Grass (Pennisetum purpureum Schum.) in Kenya. Ph.D. Thesis, University of Greenwich, London, UK, 1998. [Google Scholar]
- Paterson, D.D. Further experiments with cultivated tropical fodder crops. Emp. J. Exp. Agric. 1938, 6, 323–340. [Google Scholar]
- Burton, G.W. Registration of ‘Merkeron’ Napier grass. Crop Sci. 1989, 29, 1327. [Google Scholar] [CrossRef]
- Maher, C. Elephant grass (Pennisetum purpureum) as a cattle fodder in Kenya. East Afr. Agric. J. 1936, 1, 340–342. [Google Scholar] [CrossRef]
- Nattrass, R.M. Notes on plant diseases. East Afr. Agric. J. 1941, 7, 56. [Google Scholar]
- Lenné, J.M. A World List of Fungal Diseases of Tropical Pasture Species; CIAT: Wallingford, UK, 1990. [Google Scholar]
- Mtisi, E.; de Milliano, W. False mildew on Pearl millet and other hosts in Zimbabwe. East Afr. Agric. For. J. 1993, 59, 145–153. [Google Scholar]
- Farrell, G.; Simons, S.; Hillocks, R. A novel technique for measuring biomass loss in a diseased tussock grass. Trop. Grassl. 2000, 34, 118–124. [Google Scholar]
- Boonman, G. East Africa’s Grasses and Fodders: Their Ecology and Husbandry; Springer Science + Business Media, B.V.: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Ledermann, C. Herbarium Record 162103, Systematic Botany and Mycology Laboratory, USDA, Maryland, USA. 1998. Available online: http://nt.ars-grin.gov/fungaldatabases/fungushost (accessed on 15 September 2016).
- Sydow, H.; Sydow, P. Fungi Africani novi. Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie/begr. Von A. Engler 1911, 45, 259–265. [Google Scholar]
- Snowden, J.D. Herbarium Record 162104, Systematic Botany and Mycology Laboratory, USDA, Maryland, USA. 1998. Available online: http://nt.ars-grin.gov/fungaldatabases/fungushost (accessed on 15 September 2016).
- Watson, A.J. Foreign Bacterial and Fungus Diseases of Food, Forage, and Fiber Crops: An Annotated List; Agricutural Research service, United States Department of Agrculture: Washington, DC, USA, 1971.
- Kung’u, J.N.; Waller, J.M. Occurrence of smut of Napier grass caused by Ustilago kamerunensis H Sydow and Sydow in Kenya. Int. J. Pest Manag. 2001. [Google Scholar] [CrossRef]
- Association of Strengthening Agricultural Research in East and Central Africa (ASARECA). Workshop on Mitigating the Impact of Napier Grass Smut and Stunt Diseases for the Smallholder Dairy Sector-Sharing Results: Final Report, June 1–3, 2010; ILRI: Addis Ababa Ethiopia, Ethiopia, 2010. [Google Scholar]
- Farrell, G.; Simons, S.; Hillocks, R. Ustilago kamerunensis on Napier grass in Kenya. Int. J. Pest Manag. 2002, 48, 25–28. [Google Scholar] [CrossRef]
- Simmonds, N.W. Some speculative calculations on the dispersal of sugarcane smut disease. Sugar Cane 1994, 1, 2–5. [Google Scholar]
- Mwendia, S.; Wanyoike, M.; Wahome, R.; Mwangi, D. Effect of napier head smut disease on Napier yields and the disease coping strategies in farming systems in central Kenya. Livest. Res. Rural Dev. 2007, 19. Available online: http://www.lrrd.org/lrrd19/8/mwen19109.htm (accessed on 11 November 2016).
- Friedman, A.R.; Baker, B.J. The evolution of resistance genes in multi-protein plant resistance systems. Curr. Opin. Genet. Dev. 2007, 17, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Rausher, M.D. Co-evolution and plant resistance to natural enemies. Nature 2001, 411, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Alicai, T.; Kabirizi, J.; Byenkya, S.; Kayiwa, S.; Ebong, C. Assessment of the Magnitude and Farmers' Management Practices of the Elephant Grass Stunting Disorder in Masaka District; Namulonge Agricultural and Animal Production Research Institute: Kampala, Uganda, 2004. [Google Scholar]
- Nielsen, S.L.; Ebong, C.; Kabirizi, J.; Nicolaisen, M. First report of a 16SrXI group phytoplasma (Candidatus phytoplasma oryzae) associated with Napier grass disease in Uganda. Plant Pathol. 2007, 56, 1039. [Google Scholar] [CrossRef]
- Rosete, Y.A.; Jones, P. Phytoplasma diseases of the Gramineae. In Phytoplasmas: Genomes, Plant Hosts and Vectors; Weintraub, P.G., Jones, P., Eds.; CAB International: Wallingford, UK, 2010; pp. 170–187. [Google Scholar]
- Jones, P.; Devonshire, B.; Holman, T.; Ajanga, S. Napier grass stunt: A new disease associated with a 16SrXI group phytoplasma in Kenya. Plant Pathol. 2004, 53, 519. [Google Scholar] [CrossRef]
- Jones, P.; Arocha, T.; Zerfy, J.; Proud, J.; Abebe, G.; Hanson, J. A stunting syndrome of Napier grass in Ethiopia is associated with a 16SrIII Group phytoplasma. New Dis. Rep. 2006, 10, 2006–2019. [Google Scholar] [CrossRef]
- Jones, P.; Arocha, Y.; Zerfy, T.; Proud, J.; Abebe, G.; Hanson, J. A stunting syndrome of Napier grass in Ethiopia is associated with a 16SrIII group phytoplasma. Plant Pathol. 2007, 56, 345. [Google Scholar] [CrossRef]
- Asudi, G.O.; Van den Berg, J.; Midega, C.A.; Schneider, B.; Seemüller, E.; Pickett, J.A.; Khan, Z.R. Detection, identification, and significance of Phytoplasmas in Wild Grasses in East Africa. Plant Dis. 2016, 100, 108–115. [Google Scholar] [CrossRef]
- Marcone, C. Movement of Phytoplasmas and the Development of Diseases in the Plant; CAB International: Wallingford, UK, 2010. [Google Scholar]
- Sharon, R.; Soroker, V.; Wesley, S.D.; Zahavi, T.; Harari, A.; Weintraub, P.G. Vitex agnus-castus is a preferred host plant for Hyalesthes obsoletus. J. Chem. Ecol. 2005, 31, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-M.; Davis, R.E.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic Mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Obura, E.; Midega, C.A.; Masiga, D.; Pickett, J.A.; Hassan, M.; Koji, S.; Khan, Z.R. Recilia banda Kramer (Hemiptera: Cicadellidae), a vector of Napier stunt phytoplasma in Kenya. Naturwissenschaften 2009, 96, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Arocha, Y.; Zerfy, T.; Abebe, G.; Proud, J.; Hanson, J.; Wilson, M.; Jones, P.; Lucas, J. Identification of potential vectors and alternative plant hosts for the phytoplasma associated with Napier grass stunt disease in Ethiopia. J. Phytopathol. 2009, 157, 126–132. [Google Scholar] [CrossRef]
- Lee, I.-M.; Martini, M.; Bottner, K.; Dane, R.; Black, M.; Troxclair, N. Ecological implications from a molecular analysis of phytoplasmas involved in an aster yellows epidemic in various crops in Texas. Phytopathology 2003, 93, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- International Centre of Insect Physiology and Ecology (ICIPE). Solving Napier Stunt Disease to Save the Smallholder Dairy Sector in East Africa—A Success Story; ICIPE: Nairobi, Kenya, 2014; Available online: http://www.push-pull.net/napier_stunt_brochure.pdf (accessed on 25 July 2016).
- Mulaa, M.; Awalla, B.; Hanson, J.; Proud, J.; Cherunya, A.; Wanyama, J.; Lusweti, C.; Muyekho, F. Stunting disease incidence and impact on Napier grass (Pennisetum purpureum Schumach) in western Kenya. In 12th Biennial Kenya Agricultural Research Institute (KARI) Conference: Transforming Agriculture for Improved Livelihoods through Agricultural Product Value Chains; Wasilwa, L.A., Ed.; Kenya Agricultural Research Institute: Nairobi, Kenya, 2010; pp. 936–943. [Google Scholar]
- Asudi, G.O.; van den Berg, J.; Midega, C.A.; Pittchar, J.; Pickett, J.A.; Khan, Z.R. Napier grass stunt disease in East Africa: Farmers’ perspectives on disease management. Crop Prot. 2015, 71, 116–124. [Google Scholar] [CrossRef]
- Kabirizi, J.; Nielsen, S.; Nicolaisen, M.; Byenkya, S.; Alacai, T. Napier stunt disease in Uganda: Farmers’ perceptions and impact on fodder production. In Proceedings of the 8th African Crop Science Society Conference Proceedings, El-Minia, Egypt, 27–31 Octber 2007; Ahmed, K.Z., Ed.; Volume 8, pp. 895–897. [Google Scholar]
- Khan, Z.R.; Midega, C.A.O.; Nyang’au, I.M.; Murage, A.; Pittchar, J.; Agutu, L.O.; Amudavi, D.M.; Pickett, J.A. Farmers’ knowledge and perceptions of the stunting disease of Napier grass in Western Kenya. Plant Pathol. 2014, 63, 1426–1435. [Google Scholar] [CrossRef]
- Faleiro, F.G.; Kannan, B.; Altpeter, F. Regeneration of fertile, hexaploid, interspecific hybrids of Elephant grass and pearl millet following treatment of embryogenic calli with antimitotic agents. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 124, 57–67. [Google Scholar] [CrossRef]
- Sastry, K.S.; Zitter, T.A. Management of virus and viroid diseases of crops in the tropics. In Plant Virus and Viroid Diseases in the Tropics, volume 2: Epidemiology and management; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 149–480. [Google Scholar]
- Buxton, D.R.; Redfearn, D.D. Plant limitations to fiber digestion and utilization. J. Nutr. 1997, 127, 814S–818S. [Google Scholar] [PubMed]
- Jung, H.; Allen, M. Characteristics of plant cell walls affecting intake and digestibility of forages by ruminants. J. Anima. Sci. 1995, 73, 2774–2790. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 5th ed.; W H Freeman: New York, NY, USA, 2002. [Google Scholar]
- Bhatt, N.; Gupta, P.; Naithani, S. Preparation of cellulose sulfate from α-cellulose isolated from Lantana camara by the direct esterification method. J. Appl. Polym. Sci. 2008, 108, 2895–2901. [Google Scholar] [CrossRef]
- Klemm, D.; Heublein, B.; Fink, H.P.; Bohn, A. Cellulose: Fascinating biopolymer and sustainable raw material. Angew. Chem. Int. Ed. 2005, 44, 3358–3393. [Google Scholar] [CrossRef] [PubMed]
- Casler, M.D.; Vogel, K.P. Accomplishments and impact from breeding for increased forage nutritional value. Crop Sci. 1999, 39, 12–20. [Google Scholar] [CrossRef]
- Sattler, S.E.; Funnell-Harris, D.L.; Pedersen, J.F. Brown midrib mutations and their importance to the utilization of Maize, Sorghum, and Pearl millet lignocellulosic tissues. Plant Sci. 2010, 178, 229–238. [Google Scholar] [CrossRef]
- Guo, D.; Chen, F.; Wheeler, J.; Winder, J.; Selman, S.; Peterson, M.; Dixon, R.A. Improvement of in-rumen digestibility of alfalfa forage by genetic manipulation of lignin O-methyltransferases. Transgenic Res. 2001, 10, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, R.D.; Rancour, D.M.; Marita, J.M. Grass Cell Walls: A story of cross-Linking. Front. Plant Sci. 2017, 7, 2056. [Google Scholar] [CrossRef] [PubMed]
- Thébaud, G.; Yvon, M.; Alary, R.; Sauvion, N.; Labonne, G. Efficient transmission of ‘Candidatus phytoplasma Prunorum’ is delayed by eight months due to a long latency in its host-alternating vector. Phytopathology 2009, 99, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Mih, A.M.; Hanson, J. Identification of potyviruses infecting forage grasses in Ethiopia. J. Cameroon Acad. Sci. 2004, 4, 205–210. [Google Scholar]
- Tiley, G.E.D. Elephant Grass. Kawanda Technical Communication No.23; Kawanda Research Station: Kawanda, Unganda, 1969. [Google Scholar]
- Mudavadi, P.O.; Otieno, K.; Wanambacha, J.W.; Odenya, J.O.; Odendo, M.; Njaro, O.K. Smallholder Dairy Production and Marketing in Western Kenya: A Review of Literature; ILRI: Nairobi, Kenya, 2001. [Google Scholar]


{kind=link}
{kind=link}
| Native to *: | Number of Accessions at **: | ||||||
|---|---|---|---|---|---|---|---|
| ILRI | ICRISAT | CIAT | EMBRAPA 1 | USDA GRIN 2 | RBG 3 | Total *** | |
| Tanzania | 6 | 9 | 15 | ||||
| Uganda | 1 | 1 | |||||
| Ethiopia | 1 | 12 c | 13 | ||||
| Malawi | 1 | 2 | 1 | 4 | |||
| Mozambique | 2 | 2 | |||||
| Zimbabwe | 11 | 5 | 8 | 24 | |||
| Côte d’Ivoire | 1 | 1 | |||||
| Nigeria | 1 | 1 | 3 | 5 | |||
| Cameroon | 8 | 1 | 9 | ||||
| Sub-total | 22 | 22 | 1 | 0 | 25 | 4 | 74 |
| Collected from | |||||||
| Australia | 4 | 4 | |||||
| Brazil | 8 | 39 | 7 | 54 | |||
| Burkina Faso | 1 | 1 | |||||
| Burundi | 1 | 1 | |||||
| Central African R. | 7 | 2 | 9 | ||||
| China | 1 | 1 | |||||
| Colombia | 1 | 5 | 6 | ||||
| Costa Rica | 1 | 1 | 1 | 3 | |||
| Cuba | 2 | 4 | 6 | ||||
| DRC (Zaire) | 4 | 4 | |||||
| Ecuador | 1 | 1 | |||||
| India | 2 | 8 | 3 | 2 | 15 | ||
| Mexico | 2 | 2 | |||||
| Namibia | 1 | 1 | |||||
| Panama | 1 | 1 | |||||
| South Africa | 12 | 12 | |||||
| Sudan | 2 | 2 | |||||
| Swaziland | 6 | 3 | 9 | ||||
| USA | 16 a | 1 | 6 | 23 | |||
| Unknown | 14 b | 2 | 29 | 44 | 90 | ||
| Sub-total | 52 | 17 | 2 | 83 | 86 | 4 | 244 |
| Total | 74 | 39 | 3 | 83 | 111 | 8 | 318 |
| Country | DM (t/ha/year) | CP (%) | NDF (%) | ADF (%) | No. of Accessions Evaluated | Ref. |
|---|---|---|---|---|---|---|
| Bangladesh | 14.9–16.5 | 10.3–11.4 | NA | 29.9–45.9 | 4 | [69] |
| Brazil | 14.9–78 | NA | NA | NA | 85 | [26] |
| Ethiopia | 4.6–20.5 | 7.5–15.7 | 52–64.6 | 28.8–36.6 | 9 | [70] |
| Kenya | 12.1–19 | NA | NA | NA | 8 | [71] |
| Malaysia | 43.7–65.9 | 10–12 | 60–70 | 35–40 | 9 | [64] |
| Mexico | NA | 9.2–9.9 | 65.2–69.7 | 42.2–44.7 | 3 | [72] |
| Thailand | 27.1–58.4 | NA | NA | NA | 8 | [27] |
| USA | 25.3–28.2 | 12.42–15.68 | 62.7–66.8 | 37.2–39.6 | 2 | [57] |
| Zimbabwe | 90.2 | 5.35 | 56.8 | 39.2 | 2 | [73] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negawo, A.T.; Teshome, A.; Kumar, A.; Hanson, J.; Jones, C.S. Opportunities for Napier Grass (Pennisetum purpureum) Improvement Using Molecular Genetics. Agronomy 2017, 7, 28. https://doi.org/10.3390/agronomy7020028
Negawo AT, Teshome A, Kumar A, Hanson J, Jones CS. Opportunities for Napier Grass (Pennisetum purpureum) Improvement Using Molecular Genetics. Agronomy. 2017; 7(2):28. https://doi.org/10.3390/agronomy7020028
Chicago/Turabian StyleNegawo, Alemayehu Teressa, Abel Teshome, Alok Kumar, Jean Hanson, and Chris S. Jones. 2017. "Opportunities for Napier Grass (Pennisetum purpureum) Improvement Using Molecular Genetics" Agronomy 7, no. 2: 28. https://doi.org/10.3390/agronomy7020028
APA StyleNegawo, A. T., Teshome, A., Kumar, A., Hanson, J., & Jones, C. S. (2017). Opportunities for Napier Grass (Pennisetum purpureum) Improvement Using Molecular Genetics. Agronomy, 7(2), 28. https://doi.org/10.3390/agronomy7020028