
Minichromosomes: Vectors for Crop Improvement

Abstract
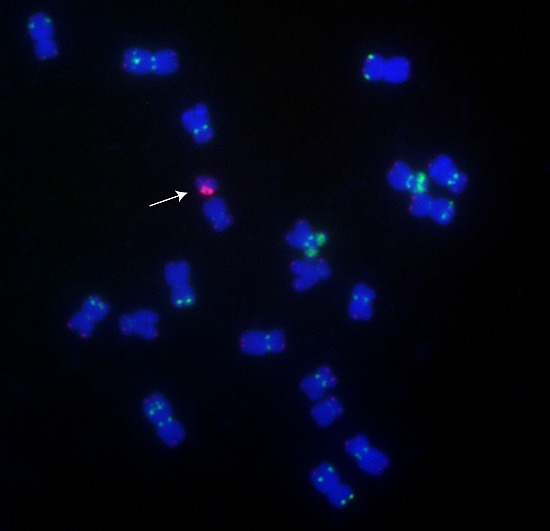
:
{kind=link}
1. Introduction
2. Discussion
2.1. Structure Overview
2.2. Centromeres
2.3. Origins of Replication
2.4. Telomeres
2.5. Utilization of B Chromosomes
2.6. Alternative Methods for Engineering Minichromosomes
2.7. Stability in Plants
2.8. Transgene Expression
2.9. Creating Large Minichromosomes with BIBACs
2.10. In Vivo Modification of Minichromosomes
2.11. Transfer of Minichromosomes with Haploid Induction
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- FAO; IFAD; WFP. Strengthening the enabling environment for food security and nutrition. In The State of Food Insecurity in the World 2014; FAO: Rome, Italy, 2014. [Google Scholar]
- Hirschi, K.D. Nutrient biofortification of food crops. Annu. Rev. Nutr. 2009, 29, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Bruulsema, T.W.; Heffer, P.; Welch, R.M.; Cakmak, I.; Moran, K. Fertilizing Crops to Improve Human Health: A Scientific Review; International Plant Nutrition Institute: Norcross, GA, USA, 2012. [Google Scholar]
- Zhu, C.; Naqvi, S.; Gomez-Galera, S.; Pelacho, A.M.; Capell, T.; Christou, P. Transgenic strategies for the nutritional enhancement of plants. Trends Plant Sci. 2007, 12, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Bevan, M.W.; Flavell, R.B.; Chilton, M. A chimaeric antibiotic resistance gene as a selectable marker for plant cell transformation. Nature 1983, 304, 184–187. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; Depicker, A.; Montagu, V.M.; Schell, J. Expression of chimaeric genes transferred into plant cells using a Ti-plasmid-derived vector. Nature 1983, 303, 209–213. [Google Scholar] [CrossRef]
- Fraley, R.T.; Rogers, S.G.; Horsch, R.B.; Sanders, P.R.; Flick, J.S.; Adams, S.P.; Bittner, M.L.; Brand, L.A.; Fink, C.L.; Fry, J.S.; et al. Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA 1983, 80, 4803–4807. [Google Scholar] [CrossRef] [PubMed]
- Murai, N.; Kemp, J.D.; Sutton, D.W.; Murray, M.G.; Slightom, J.L.; Merlo, D.; Reichert, N.A.; Sengupta-Gopalan, C.; Stock, C.A.; Barker, R.F.; et al. Phaseolin gene from bean is expressed after transfer to sunflower via tumor-inducing plasmid vectors. Science 1983, 222, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Han, F.; Birchler, J.A. Engineered minichromosomes in plants. Curr. Opin. Biotechnol. 2007, 18, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Gaeta, R.T.; Masonbrink, R.E.; Krishnaswamy, L.; Zhao, C.; Birchler, J.A. Synthetic chromosome platforms in plants. Annu. Rev. Plant Biol. 2012, 63, 307–330. [Google Scholar] [CrossRef] [PubMed]
- Kanizay, L.; Dawe, R. Centromeres: Long intergenic spaces with adaptive features. Funct. Integr. Genomics 2009, 9, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Burrack, L.S.; Berman, J. Neocentromeres and epigenetically inherited features of centromeres. Chromosome Res. 2012, 20, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Lamb, J.C.; Birchler, J.A. Chromosome painting using repetitive DNA sequences as probes for somatic chromosome identification in maize. Proc. Natl. Acad. Sci. USA 2004, 101, 13554–13559. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.X.; Marshall, J.B.; Topp, C.; Mroczek, R.; Kato, A.; Nagaki, K.; Birchler, J.A.; Jiang, J.; Dawe, R.K. Centromeric retroelements and satellites interact with maize kinetochore protein CENH3. Plant Cell 2002, 14, 2825–2836. [Google Scholar] [CrossRef] [PubMed]
- Wolfgruber, T.K.; Sharma, A.; Schneider, K.L.; Albert, P.S.; Koo, D.; Shi, J.; Gao, Z.; Han, F.; Lee, H.; Xu, R.; et al. Maize centromere structure and evolution: Sequence analysis of centromeres 2 and 5 reveals dynamic Loci shaped primarily by retrotransposons. PLoS Genet. 2009, 5, e1000743. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Melo, J.R.; Nagaki, K.; Talbert, P.B.; Henikoff, S.; Dawe, R.K.; Jiang, J. Maize centromeres: Organization and functional adaptation in the genetic background of oat. Plant Cell 2004, 16, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yue, W.; Li, D.; Wang, R.; Kong, X.; Lu, K.; Wang, G.; Dong, Y.; Jin, W.; Zhang, X. Structure and dynamics of retrotransposons at wheat centromeres and pericentromeres. Chromosoma 2008, 117, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Nasuda, S.; Hudakova, S.; Schubert, I.; Houben, A.; Endo, T. Stable barley chromosomes without centromeric repeats. Proc. Natl. Acad. Sci. USA 2005, 102, 9842–9847. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Yu, H.; Huang, J.; Yi, C.; Gu, M. Unstable transmission of rice chromosomes without functional centromeric repeats in asexual propagation. Chromosome Res. 2009, 17, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Lv, Z.; Gao, Z.; Wu, H.; Pang, J.; Zhang, B.; Dong, Q.; Guo, X.; Wang, X.; Birchler, J.A.; et al. De novo centromere formation on a chromosome fragment in maize. Proc. Natl. Acad. Sci. USA 2013, 110, 6033–6036. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Lamb, J.C.; Birchler, J.A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Gao, Z.; Birchler, J.A. Reactivation of an inactive centromere reveals epigenetic and structural components for centromere specification in maize. Plant Cell 2009, 21, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A.; Gao, Z.; Sharma, A.; Presting, G.G.; Han, F. Epigenetic aspects of centromere function in plants. Curr. Opin. Plant Biol. 2011, 14, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Furuyama, T. Epigenetic inheritance of centromeres. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.C.; Mechali, M. DNA replication origins. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Hirose, Y.; Langmore, J. Long g tails at both ends of human chromosomes suggest a C strand degradation mechanism for telomere shortening. Cell 2001, 88, 657–666. [Google Scholar] [CrossRef]
- Fajkus, J.; Fulnecková, J.; Hulanova, M.; Berkova, K.; Ríha, K.; Matyásek, R. Plant cells express telomerase activity upon transfer to callus culture, without extensively changing telomere lengths. Mol. Gen. Genet. 1998, 260, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.S.; McKnight, T.D.; Shippen, D.E. Characterization and developmental patterns of telomerase expression in plants. Proc. Natl. Acad. Sci. USA 1996, 93, 14422–14427. [Google Scholar] [CrossRef] [PubMed]
- Kilian, A.; Heller, K.; Kleinhofs, A. Development patterns of telomerase activity in barley and maize. Plant Mol. Biol. 1998, 37, 621–628. [Google Scholar] [CrossRef]
- Hemann, M.T.; Strong, M.A.; Hao, L.; Greider, C.W. The Shortest Telomere, Not Average Telomere Length, Is Critical for Cell Viability and Chromosome Stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef]
- Shakirov, E.V.; Shippen, D.E. Length Regulation and dynamics of individual telomere tracts in wild-type arabidopsis. Plant Cell 2004, 16, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Houben, A. B chromosomes in plants: Escapees from the A chromosome genome. Trends Plant Sci. 2003, 8, 417–423. [Google Scholar] [CrossRef]
- Randolph, L.F. Genetic characteristics of the B chromosomes in maize. Genetics 1941, 26, 608–631. [Google Scholar] [PubMed]
- Staub, R.W. Leaf striping correlated with the presence of B chromosomes in maize. J. Hered. 1987, 78, 71–74. [Google Scholar]
- Roman, H. Mitotic nondisjunction in the case of interchanges involving the B-type chromosome in maize. Genetics 1947, 32, 391–409. [Google Scholar] [PubMed]
- Gaeta, R.T.; Masonbrink, R.E.; Zhao, C.; Sanyal, A.; Krishnaswamy, L.; Birchler, J.A. In vivo modification of a maize engineered minichromosome. Chromosoma 2013, 122, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Masonbrink, R.E.; Birchler, J.A. Accumulation of multiple copies of maize minichromosomes. Cytogenet. Genome Res. 2012, 137, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Yan, H.; Yu, H.; Tang, S.; Jiang, J.; Gu, M.; Zhu, L. Development and applications of a complete set of rice telotrisomics. Genetics 2001, 157, 361–368. [Google Scholar] [PubMed]
- Yu, W.; Han, F.; Gao, Z.; Vega, J.M.; Birchler, J.A. Construction and behavior of engineered minichromosomes in maize. Proc. Natl. Acad. Sci. USA 2007, 104, 8924–8929. [Google Scholar] [CrossRef] [PubMed]
- Teo, C.H.; Ma, L.; Kapusi, E.; Hensel, G.; Kumlehn, J.; Schubert, I.; Houben, A.; Mette, M.F. Induction of telomere-mediated chromosomal truncation and stability of truncated chromosomes in Arabidopsis thaliana. Plant J. 2011, 68, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Kapusi, E.; Ma, L.; Teo, C.H.; Hensel, G.; Himmelbach, A.; Schubert, I.; Mette, M.; Kumlehn, J.; Houben, A. Telomere-mediated truncation of barley chromosomes. Chromosoma 2012, 121, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Gao, Z.; Yu, W.; Birchler, J.A. Minichromosome analysis of chromosome pairing, disjunction, and sister chromatid cohesion in maize. Plant Cell 2007, 19, 3853–3863. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.D.; Gabay-Laughnan, S. Cytoplasmic male sterility and fertility restoration by nuclear genes. In Molecular Biology and Biotechnology of Plant Organelles; Daniell, H., Chase, C.D., Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 22, pp. 593–621. [Google Scholar]
- Birchler, J.; Krishnaswamy, L.; Gaeta, R.; Masonbrink, R.; Zhao, C. Engineered Minichromosomes in Plants. CRC Crit. Rev. Plant. Sci. 2010, 29, 135–147. [Google Scholar] [CrossRef]
- Ye, F.; Signer, E.R. RIGS (repeat-induced gene silencing) in Arabidopsis is transcriptional and alters chromatin configuration. Proc. Natl. Acad. Sci. USA 1996, 93, 10881–10886. [Google Scholar] [CrossRef] [PubMed]
- Matzke, A.J.; Matzke, M.A. Position effects and epigenetic silencing of plant transgenes. Curr. Opin. Plant Biol. 1998, 1, 142–148. [Google Scholar] [CrossRef]
- Liu, W.; Yuan, J.S.; Stewart, C. Advanced genetic tools for plant biotechnology. Nat. Rev. Genet. 2013, 14, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Halpin, C. Gene stacking in transgenic plants—The challenge for 21st century plant biotechnology. Plant Biotechnol. J. 2005, 3, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Shizuya, H.; Birren, B.; Kim, U.; Mancino, V.; Slepak, T.; Tachiiri, Y.; Simon, M. Cloning and stable maintenance of 300-kilobase-pair fragments of human DNA in Escherichia coli using an F-factor-based vector. Proc. Natl. Acad. Sci. USA 1992, 89, 8794–8797. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.M.; Frary, A.; Lewis, C.; Tanksley, S.D. Stable transfer of intact high molecular weight DNA into plant chromosomes. Proc. Natl. Acad. Sci. USA 1996, 93, 9975–9979. [Google Scholar] [CrossRef] [PubMed]
- Vega, J.M.; Yu, W.; Han, F.; Kato, A.; Peters, E.M.; Zhang, Z.S.; Birchler, J.A. Agrobacterium-mediated transformation of maize (Zea mays) with Cre-lox site specific recombination cassettes in BIBAC vectors. Plant Mol. Biol. 2008, 66, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.M.; Frary, A.; Xu, Y.; Tanksley, S.D.; Zhang, H. Construction of tomato genomic DNA libraries in a binary-BAC (BIBAC) vector. Plant J. 1999, 18, 223–229. [Google Scholar] [CrossRef]
- Grindley, N.D.; Whiteson, K.L.; Rice, P.A. Mechanisms of site-specific recombination. Annu. Rev. Biochem. 2006, 75, 567–605. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yau, Y.; Perkins-Balding, D.; Thomson, J.G. Recombinase technology: Applications and possibilities. Plant Cell Rep. 2011, 30, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Gidoni, D.; Srivastava, V.; Carmi, N. Site-specific excisional recombination strategies for elimination of undesirable transgenes from crop plants. In Vitro Cell. Dev. Biol. Plant 2008, 44, 457–467. [Google Scholar] [CrossRef]
- Ow, D.W. 2004 SIVB congress symposium proceeding: Transgene management via multiple site-specific recombination systems. In Vitro Cell. Dev. Biol. Plant 2005, 41, 213–219. [Google Scholar] [CrossRef]
- Coe, E.H. A line of maize with high haploid frequency. Am. Natl. 1959, 93, 381–382. [Google Scholar]
- Zhang, Z.; Qiu, F.; Liu, Y.; Ma, K.; Li, Z.; Xu, S. Chromosome elimination and in vivo haploid production induced by stock 6-derived inducer line in maize (Zea mays L.). Plant Cell Rep. 2008, 27, 1851–1860. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xu, X.; Xie, H.; Chen, S.; Jin, W. Fertilization and uniparental chromosome elimination during crosses with maize haploid inducers. Plant Physiol. 2013, 163, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Kato, A. Chromosome doubling of haploid maize seedlings using nitrous oxide gas at the flower primordial stage. Plant Breed. 2002, 121, 370–377. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cody, J.P.; Swyers, N.C.; McCaw, M.E.; Graham, N.D.; Zhao, C.; Birchler, J.A. Minichromosomes: Vectors for Crop Improvement. Agronomy 2015, 5, 309-321. https://doi.org/10.3390/agronomy5030309
Cody JP, Swyers NC, McCaw ME, Graham ND, Zhao C, Birchler JA. Minichromosomes: Vectors for Crop Improvement. Agronomy. 2015; 5(3):309-321. https://doi.org/10.3390/agronomy5030309
Chicago/Turabian StyleCody, Jon P., Nathan C. Swyers, Morgan E. McCaw, Nathaniel D. Graham, Changzeng Zhao, and James A. Birchler. 2015. "Minichromosomes: Vectors for Crop Improvement" Agronomy 5, no. 3: 309-321. https://doi.org/10.3390/agronomy5030309
APA StyleCody, J. P., Swyers, N. C., McCaw, M. E., Graham, N. D., Zhao, C., & Birchler, J. A. (2015). Minichromosomes: Vectors for Crop Improvement. Agronomy, 5(3), 309-321. https://doi.org/10.3390/agronomy5030309