Abstract
Temperate grazing systems require advanced management strategies to simultaneously enhance both productivity and resilience. Multispecies pasture (MSS) mixtures offer a promising alternative to conventional Lolium perenne-Trifolium repens (LP-TR) systems by leveraging niche complementarity through diverse and deeper rooting profiles that enhance drought tolerance and optimize nutrient acquisition from heterogeneous soil layers. In a field study, we compared LP-TR pastures with three functionally distinct MSS pastures subjected to varying rotation lengths (RL) and defoliation intensities (DIs). Seasonal and annual dry matter (DM) yield and botanical composition were assessed. MSS consistently outperformed LP-TR in total DM production, especially under shorter RL, which had a more pronounced effect on annual and spring biomass than DI. An 8 cm defoliation height generally enhances DM accumulation across systems. Species interactions significantly influenced botanical composition, with Plantago lanceolata contributing to greater pasture stability under environmental stress, accounting for 24–61% of total herbage DM. Furthermore, RL and DI interactions affected species prevalence, underscoring the need for adaptive, species-specific management strategies. These findings demonstrate that integrating strategic species selection with tailored defoliation practices in MSS mixtures can substantially enhance pasture productivity, compositional stability, and long-term sustainability of temperate grazing systems.
1. Introduction
Climate change (CC) poses a significant threat to pasture-based temperate livestock systems by disrupting seasonal patterns of forage availability and quality, increasing environmental stress, and ultimately affecting animal productivity and health [1,2]. In temperate regions, grazing systems are commonly based on improved pastures dominated by perennial ryegrass (Lolium perenne L.; LP) and white clover (Trifolium repens L.; TR) [3]. These species are characterized by shallow root systems, which limit their productivity and resilience during drought and heat stress, leading to higher supplementary feeding to satisfy animal nutritional needs [4,5]. Additionally, these pastures depend heavily on synthetic nitrogen (N) fertilization to maintain optimal growth [6], a practice that is increasingly scrutinized due to its environmental impacts, including greenhouse gas emissions, water pollution, and biodiversity loss [7].
Given the critical role of temperate pastures in global food security and land use [8,9], and considering that they account for over 50% of global forage intake by livestock [8], there is a need for sustainable strategies to enhance the resilience and productivity of pasture-based systems under CC conditions. One such strategy is increasing species diversity in pasture [6,10,11]. Multispecies pastures (MSSs), which are typically composed of from three to twelve plant species from different functional groups—including grasses, legumes, and herbs [12]—are strategically designed to enhance interspecific complementarity, such as biological N fixation by legumes (reducing reliance on off-farm inputs and production costs; [6]) and deeper rooting systems that improve drought tolerance and carbon sequestration in deeper soil layers [13]. Several studies report that these pastures improve forage dry matter (DM) production and enhance seasonal stability on both yield and herbage quality [14,15,16], thus improving resource-use efficiency and ecosystem resilience. In the meta-analysis of Jordon et al. [10], it was reported that forage yield increased by 1.63 t ha−1 for every additional meter of rooting depth compared to conventional pastures of perennial ryegrass and white clover, underscoring the benefits of multispecies pastures.
Deep-rooted forage species such as plantain (Plantago lanceolata L.; PL) and chicory (Cichorium intybus L.; CI) provide valuable ecosystem services due to their superior drought tolerance and ability to extract moisture from deeper soil layers—traits that enhance forage DM production and persistence under water-limited conditions [17,18,19]. In addition, these species also contribute to reducing the use of chemical anthelmintics, enhancing animal yields, and mitigating the environmental impacts—such as methane emissions and urinary nitrogen excretion—of grazing systems through their secondary metabolite compounds [20,21,22]. More diverse pastures may also reduce dependence on N fertilizers [15,23], and improved nutritional profiles compared to pure grass or legume pastures [6,24], reducing the requirements for supplementary concentrates and improving animal health and performance [25] and ultimately supporting more sustainable grazing systems.
Despite these potential benefits, maintaining productivity and desirable botanical composition of multispecies pastures is highly dependent on defoliation management. This is primarily due to the current lack of comprehensive field-based information regarding optimal defoliation strategies for MSS permanent pastures. Additionally, defoliation criteria commonly applied to MSS are largely adapted from those developed for L. perenne and T. repens mixtures (LP-TR), which may not fully account for the functional diversity present in more complex pasture systems [25]. Effective defoliation management, including optimized cutting frequency and residual height, plays a crucial role in the productivity and botanical composition of permanent pastures, especially in multispecies mixtures. While the effects of defoliation frequency and intensity on individual forage species are well documented [3,26,27], there are still critical knowledge gaps regarding the functional response of complex mixtures. For example, how should species be selected and combined to promote persistence and complementarity? Can multispecies pastures maintain productivity and composition under standard grazing regimes? To what extent do defoliation frequency and intensity shape competitive dynamics among functional groups in these complex systems?
Baker et al. [28] compared DM production and botanical composition of three pastures types: a perennial ryegrass monoculture (LP; 250 kg N ha−1 yr−1), a perennial ryegrass–white clover mixture (LP-TR; 90 kg N ha−1 yr−1), and a multispecies pasture (MSS), comprising grasses, legumes, and forbs, such as plantain and chicory (also receiving 90 kg N ha−1 yr−1), subjected to different defoliation frequencies (every 21 or 28 days) and residual heights (4 or 6 cm) over two growing seasons (from March to November). The authors reported that MSSs yielded 1923 kg DM ha−1 yr−1 more under less frequent defoliation (longer intervals), while LP and LP-TR pastures showed no significant response to defoliation frequency. Notably, MSSs maintained a more stable botanical composition, with fewer weed species regardless of residual height, whereas LP and LP-TR pastures showed an increase in unsown species—particularly under low residual heights.
Supporting these findings, other studies [29,30] confirm that increased species richness improves summer productivity, drought resilience, and weed suppression, although some functional species like red clover, chicory, and plantain may decline over time, highlighting the need for adaptive management strategies to sustain the benefits of complex pastures.
In this study, we hypothesized that multispecies pastures composed of grasses, legumes, and herbs would outperform conventional LP-TR mixtures in terms of forage yield, seasonal productivity, and stability of botanical composition, with longer defoliation intervals (long rotation length) and lower defoliation intensity (residual height). The objective of this study was to evaluate the effects of defoliation frequency (based on days according to season) and intensity (residual heights of 6 or 8 cm) on annual and seasonal DM yield and botanical composition of LP-TB compared to three types of multispecies pastures.
2. Materials and Methods
2.1. Experimental Site
This study was conducted at the experimental research station “Oromo” of the University of Chile, Purranque, Los Lagos, Chile (Latitude 40°53′ S, Longitude 73°06′30″ O; 150 m.s.n.m.), between August 2023 and June 2025. The soil type at the experimental site is classified as Typic Hapludand (Andisol) [31]. Prior to the establishment of the experimental plots, soil tests were taken in September 2022. The chemical composition of the soil (20 cm sampling depth) was 15.59 mg kg−1 total Kjeldahl N, 9.1 mg kg−1 Olsen P, 125.3 mg kg−1 exchangeable K, 0.95 cmol (+) kg−1 exchangeable Mg, 0.13 cmol (+) kg−1 Al, 22.5% of organic matter, and a pH (H2O) of 5.87. Rainfall, air, and soil temperature (20 cm depth) were recorded daily throughout the experimental period at the study site using a weather station (ATMOS 41, METER Group Inc., Pullman, WA, USA) and three soil temperature sensors (TEROS 12, METER Group Inc., Pullman, USA). The annual fertilization for the site consisted of the application of 51 kg N ha−1, 70 kg P2O5 ha−1 (30 kg P ha−1), and 62 kg K2O ha−1 (51.5 kg K ha−1) in the form of 15-20-18 (QROPMIX NPK, SQM, Santiago, Chile) applied in September 2022. Calcium carbonate (CaCO3) was spread in June 2022 at a rate of 1000 kg ha−1 (400 kg Ca ha−1)
2.2. Meteorological Data
Mean total annual rainfall during the 2023–2024 period (corresponding to pasture establishment) was 30% lower (−300 mm) than the 10-year average, with particularly pronounced deficits from June to August. Precipitation during spring (years 2023 and 2024) and summer (years 2024 and 2025) seasons was similar to the 10-year average. Mean annual air temperature during the experimental period was 0.3 °C higher than the 10-year average. In contrast, mean annual soil temperature was 0.18 °C lower and 1.17 °C higher than the 10-year average for the years 2023 and 2024, respectively (Table 1).

Table 1.
Monthly rainfall (mm), average daily air temperature (°C), and average daily soil temperature (°C) at the experimental site between 2023 and 2025 compared to the 10-year average (2014–2024) at Oromo Experimental Station, Purranque, Chile.
2.3. Establishment and Experimental Design
Prior to establishment, the experimental site was sprayed with glyphosate herbicide at a rate of 1.5 L ha−1 on two occasions (August and September 2023). Then, the site was cultivated using a disc harrow, and sown with a hand seeder (Earthway 1001b, SOLO Kleinmotoren GmbH, Stuttgart, Germany) on the 7th and 8th of October 2023. No fertilizers or irrigation were applied during the establishment or experimental period.
The 6000 m2 study area was arranged in a randomized complete block design according to a 4 × 2 × 2 factorial arrangement (four types of pastures, two defoliation frequencies, and two defoliation intensities). The experimental site was divided into 96 plots of 24 m2 each, considering six replicates (n = 6) for each treatment combination. Four types of pastures were used: one pasture composed of perennial ryegrass (Lolium perenne L.; LP) and white clover (Trifolium repens L.; TR), and three multispecies pastures composed of different proportions of seven species (LP, TR, cocksfoot (Dactylis glomerata L.; DG), tall fescue (Festuca arundinacea L.; LA), festulolium (×Festulolium; FL), plantain (Plantago lanceolata L.; PL), and chicory (Cichorium intybus L.; CI). The seed proportion for each species within each pasture, the cultivars for each species utilized in the experiment, and the respective seeding rate (g m−2, seeds m−2) of each species used are detailed in Table 2.
Table 2.
The proportion and seeding rates of each species per pasture type.
2.4. Defoliation Management
From seedling emergence until all plots reached an average herbage mass of 2500 kg DM ha−1 (91 days post-sowing; 7 January 2024), herbage production was measured every 15 days by cutting the herbage to an undisturbed residual height of 3 cm from four quadrats (0.0625 m2) per plot. The herbage collected in each sample was immediately dried to a constant weight at 105 °C for 24 h in an air-forced oven (UM500, Memmert GmbH and Co KG, Schwabach, Germany) to determine DM content. On 7 January 2024, defoliation management was initiated, using two levels of defoliation intensity (DI) (undisturbed residual heights of 5 cm and 8 cm) and two grazing rotation lengths (RL, days) (short RL and long RL). The DIs were selected based on previous studies [26,27,32] to balance the regrowth requirements of all component species. The defoliation treatments were applied using a lawn tractor (Husqvarna TS148, Huskvarna, Jönköping, Sweden), with the cutting height adjusted according to the specific defoliation intensity. The duration of the rotation length was considered adaptive between seasons, maintaining an average interval of from 7 to 12 days between the short RL and long RL treatments, depending on the season. Seasonal defoliation criteria are detailed in Table 3.
Table 3.
Defoliation criteria, including defoliation intensity (DI) and rotation length (RL), used in each season according to the treatments.
2.5. Herbage Measurements
Herbage production was measured in every defoliation event according to the corresponding RL and DI in each treatment by cutting herbage from four quadrats (0.0625 m2) per plot. The herbage collected in each sample was immediately dried to a constant weight at 105 °C for 24 h to determine DM content. Seasonal herbage production was determined by summing the herbage mass accumulated at each defoliation event for each plot. Annual herbage production was calculated by summing the accumulated herbage DM production at each defoliation event and the herbage DM produced between seedling emergence and the initial homogenization cut. The defoliation season was categorized as summer (January, February, and March), autumn (April, May, and June), winter (July, August, and September), and spring (October, November, and December). Herbage measurements were taken at all defoliation events between January 2024 and June 2025. Herbage daily growth rate (kg DM ha−1 day−1) was estimated weekly by cutting four quadrats (0.0625 m2) per plot to the targeted post-defoliation height according to the defoliation intensity treatments. The herbage collected in each sample was immediately dried to a constant weight at 105 °C for 24 h to determine DM concentration.
2.6. Botanical Composition
Prior to each defoliation event, a 100 g fresh weight sub-sample of herbage (cut to the respective post-defoliation residual height) was taken from six quadrats (0.0625 m2) per plot. Subsequently, the samples were placed in plastic bags and stored at −20 °C. Then, the samples were unfrozen and separated into sown and unsown species. Every species was weighed fresh and then dried to a constant weight at 60 °C for 48 h in an air-forced oven, to determine the proportional contribution of each species (% DM) to overall DM, expressed as a decimal.
2.7. Statistical Analysis
Herbage data were analyzed using the linear mixed model procedure in NCSS (version 2019, Kaysville, UT, USA). Each plot within a treatment was considered the experimental unit (n = 6). The variables of herbage production were evaluated using fixed effects for pasture type, defoliation intensity and rotation length, and season, along with their interactions. Botanical composition was analyzed within each pasture type, assessing the effects of season, defoliation intensity, rotation length, and their interactions. Differences among mean values were determined using least squares means, with pairwise comparisons by Tukey’s test. Statistical significance was set at p < 0.05, and results are expressed as least square means ± standard error of the mean (SEM).
3. Results
3.1. Herbage Measurements
3.1.1. Annual and Seasonal Dry Matter Production
Throughout the experimental period, all multispecies pastures (MSSs) produced significantly more herbage DM than LP-TR (p < 0.0001), with MSS2 exhibiting a 12% greater DM production (+1522 kg DM ha−1) compared to LP-TR. Significant effects of RL (p < 0.0001) and DI (p = 0.001) were observed. Short RL produced more herbage DM than long RL, and a DI of 8 cm produced more herbage DM than 5 cm (Table 4). Furthermore, a significant RL × DI interaction was detected (p = 0.0137), where the 8 cm/short RL produced the highest herbage DM compared to all other treatments (Table 5).
Table 4.
Effect of pasture type (PT), rotation length (RL), and defoliation intensity (DI) on annual and seasonal herbage dry matter production (kg DM ha−1).
Table 5.
Interaction between rotation length (RL) and defoliation intensity (DI) on annual and seasonal herbage dry matter production.
During the summer, pasture type had a significant effect on herbage production (p = 0.0135), with MSS3 and MSS2 exhibiting the highest DM yields, 13.6% and 9.3% greater, respectively, than LP-TR. Additionally, short RL resulted in higher herbage production than long RL (p = 0.0056; Table 4). No significant effects of DI or interactions were observed (p > 0.05).
In autumn, MSS1 and MSS2 produced more herbage DM than LP-TR and MSS3 (p < 0.0001). Shorth RL produced 30.5% more herbage DM than long RL (p < 0.0001), while a DI of 5 cm resulted in higher DM production compared to a DI of 8 cm (p < 0.0001; Table 4). A significant effect of PT × RL (p = 0.0026) and RL × DI (p = 0.0147) interactions was observed. In this season, DM production was higher in MSS1 (2168 kg DM ha−1) and MSS2 (2142 kg DM ha−1) defoliated under short RL, while herbage DM production was lower in LP-TR (1033 kg DM ha−1) and MSS3 (1101 kg DM ha−1) under long RL. In addition, significantly more herbage DM was produced in short RL/5 cm compared to all other treatments (p = 0.0147; Table 5).
During the winter, DM production was significantly higher in MSS2 (p < 0.0001). No significant effects of RL or DI were observed (p > 0.05). Significant interactions between PT and DI (p = 0.0298) and between RL and DI (p = 0.006; Table 4) were detected. The highest DM production were registered in MSS1 (1635 kg DM ha−1) and MSS2 (1764 kg DM ha−1) at a defoliation intensity of 5 cm, and in MSS2 (1778 kg DM ha−1) at a defoliation intensity of 8 cm, while the lowest DM production was observed in LP-TR (1303 kg DM ha−1) at a defoliation intensity of 8 cm. Furthermore, significantly more herbage DM was produced with 5 cm DI and long RL, with DM production similar to that obtained with 8 cm DI and short RL (p = 0.006; Table 5).
In spring, herbage DM production was significantly higher in MSS2 (p = 0.0012), but statistically similar to LP-TR. Forage production was 15.8% greater with short RL compared to long RL (p < 0.0001), while a 17.7% increase in forage production was observed with a defoliation intensity of 5 cm compared to 8 cm (p < 0.0001; Table 4). A significant interaction between RL and DI was observed (p = 0.0005), with a difference of 1736 kg DM ha−1 between the highest-yielding treatment (8 cm/short RL) and the lowest-yielding treatment (5 cm/long RL; Table 5).
3.1.2. Pastures Daily Growth Rates
During the winter, the daily pasture growth rate (kg DM ha−1 day−1) was significantly higher for the multispecies pastures (MSSs) compared to the LP-TR (Figure 1; p < 0.0001). On average, during the spring—the primary grazing season at the study site—the growth rate of MSS2 was 59.6 kg DM ha−1 day−1, which was significantly higher (p = 0.0007) than that observed for MSS1 and MSS3, but similar to LP-TR. Peak daily growth rates were observed in November, with LP-TR (68.2 kg DM ha−1 day−1) and MSS2 (69.1 kg DM ha−1 day−1) exhibiting significantly higher growth rates (p < 0.0001) than MSS1 and MSS3 (63.7 and 61.9 kg DM ha−1 day−1, respectively). During the summer, a period of very low precipitation in the experimental site, no significant differences (p > 0.05) in daily growth rates were observed among the different pastures. During autumn, the growth rates of MSS1 and MSS2 were 29% higher (p < 0.0001) than those observed for LP-TR and MSS3.
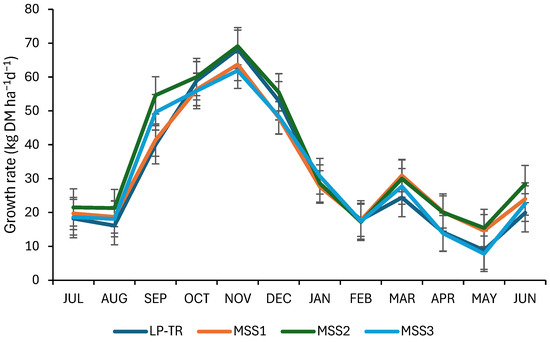
Figure 1.
Effect of pasture type on daily herbage growth rate (kg DM ha−1 day−1) during a growing season. LP-TR: L. perenne and T. repens pasture; MSS 1 to MSS3: multispecies pastures.
3.1.3. Botanical Composition
In LP-TR, the contribution of L. perenne exhibited seasonal variation, being higher during the winter and autumn (p < 0.001), and lower during the spring and summer (Table 6). The proportion of this species decreased by 5% (p = 0.0260) under short RL compared to long RL. In T. repens and unsown species, a significant increase in their contributions was observed during the spring and summer (p < 0.0001), averaging 13.5% and 21.5% for T. repens and unsown species, respectively, across both seasons. A significant interaction between RL and DI was detected for the contribution of T. repens (p < 0.001), where its contribution was greatest in the 5 cm/short RL and 8 cm/long RL treatments, relative to other combinations (Table 6). Rotation length had a significant effect on the proportion of unsown species (p < 0.0001), increasing their contribution by 6% under short RL defoliation compared to long RL (Table 6).
Table 6.
Effect of season (SEA), rotation length (RL), and defoliation intensity (DI) on the contribution of each species (sown and unsown) to herbage DM production per pasture type.
The contribution of all species within MSS1 varied significantly with the season (p < 0.02), with a notable increase in the contribution of P. lanceolata during the autumn and winter (>50%), and of unsown species during the spring and summer (averaging 18%; Table 6). No effect of RL on species contribution was observed (p > 0.05). Defoliation to a 5 cm residual height resulted in a decrease (p < 0.006) in the contribution of L. perenne, F. arundinacea, D. glomerata, ×Festulolium, and unsown species, compared to defoliation to an 8 cm residual height. For P. lanceolata, defoliation to a 5 cm residual height caused a 13% increase (p < 0.001) in its contribution to DM production, in relation to an 8 cm residual height. The contribution of T. repens tended to be greater (p = 0.0518) with defoliation to an 8 cm residual height, compared to 5 cm (Table 6). A significant interaction between RL × DI was observed for the contribution of C. intybus and unsown species (p < 0.03), where, for C. intybus, the higher contributions were observed in the 5 cm/ short RL and 8 cm/long RL treatments, while for unsown species, their contribution was higher under defoliation to an 8 cm residual height with a short RL (Table 7).
Table 7.
Interaction between rotation length (RL) and defoliation intensity (DI) on the contribution of each species (sown and unsown) to herbage DM production per pasture type.
Similar results to those reported for MSS1 were observed in MSS2, where the contribution of all species varied significantly with the season (p < 0.02), particularly P. lanceolata, which increased its contribution during autumn and winter to over 58% of DM production, and the unsown species, which increased their contribution during spring and summer (Table 6). No effect of RL was observed on the contribution of sown species (p > 0.05), whereas for unsown species, short RL resulted in a 4% increase in contribution compared to long RL (p = 0.0126). Defoliation to a 5 cm residual height caused a decrease (p < 0.001) in the contribution of all grasses and a 16% increase in the contribution of P. lanceolata, in relation to defoliation to 8 cm residual height (Table 6). Furthermore, a significant interaction between RL × DI was observed, where defoliation to an 8 cm residual height with long RL increased the contribution of all grasses (p < 0.007), while for P. lanceolata, its contribution increased (p = 0.0211) with defoliation to a 5 cm residual height in both short RL and long RL (Table 7).
In MSS3, all species exhibited changes in their contribution due to the season (p < 0.03), especially the significant increase in the contribution of P. lanceolata during autumn and winter (averaging 48% for both seasons; p < 0.001) compared to spring (24%) and summer (35%), and the higher contribution of unsown species during spring and summer (averaging 16% for both seasons; p < 0.001) compared to autumn and winter (averaging 3%; Table 6). There was no effect of RL on the contribution of species (p > 0.05).
The contribution of all grasses decreased significantly (p < 0.001) with defoliation to a 5 cm residual height compared to defoliation to an 8 cm residual height, while for forage herbs (P. lanceolata and C. intybus), defoliation to a 5 cm residual height generated a significant increase in their contribution (p < 0.03). A significant interaction between RL × DI was observed in the contribution of all grasses, T. repens, and P. lanceolata (p < 0.007; Table 7). In the case of grasses, the higher contribution to DM production was observed with defoliation to 8 cm/long RL, while the contribution of P. lanceolata reached 50% under defoliation to 5 cm/long RL, being significantly greater (p < 0.0001) than the other treatment combinations. For T. repens, the greatest contribution was observed at 5 cm/short RL, although it did not exceed 5% of DM production (Table 7).
4. Discussion
Reducing the vulnerability of grazing production systems is of paramount importance, and one promising strategy involves utilizing multispecies pastures. These pastures, composed of drought-tolerant species from diverse functional groups and managed under lower inputs [15], are thought to enhance resilience. Given that defoliation management, particularly the intensity and frequency of grazing, is a well-understood key driver of productivity in such systems [33], our study aimed to evaluate the effects of rotation length (RL) and defoliation intensity (DI) on both annual and seasonal dry matter (DM) production and botanical composition. By comparing conventional L. perenne-T. repens mixtures with three distinct MSS mixtures, our results provide important support for these strategies. Specifically, they offer critical insights into the complex interplay between environmental conditions, management practices (RL and DI), and species dynamics in temperate grazing systems.
4.1. Annual and Seasonal Forage Dry Matter Production
Our results demonstrate that MSS consistently outperformed the LP-TR mixture in terms of total herbage DM production. This finding aligns with a growing body of evidence supporting the ecological and agronomic benefits of diverse pasture systems [6,10], where complex interactions between species can lead to increased resource utilization and resilience to environmental stress. This enhanced performance is often attributed to complementary resource strategies of diverse forage mixtures, particularly the ability of legumes to fix nitrogen and deep-rooted forbs and grasses to mobilize water and nutrients from deeper soil layers, effectively redistributing these resources throughout the soil profile [6]. Previous studies, such as Baker et al. [28], have shown that multispecies pastures can yield significantly more forage DM (e.g., 1923 kg DM ha−1 yr−1 more)—under defoliation intervals of 28 days vs. 21 days—compared to conventional LP-TR pastures. This suggests that the functional diversity within MSSs allows for a more efficient capture and utilization of resources over extended periods, particularly under conditions that might stress monocultures. For example, the inclusion of deep-rooting forbs, such as P. lanceolata and C. intybus, within the MSS mixtures may have facilitated access to water and nutrients from deeper soil layers, particularly during periods of drought stress [17,18]. The fact that the production advantage of MSS2 and MSS3 became most pronounced during periods of environmental stress, such as summer and winter, coinciding with periods when P. lanceolata made a significantly larger contribution to total biomass, underscores the importance of this species in maintaining productivity under challenging conditions.
Differences among the MSS mixtures highlight the importance of species selection and seasonal growth patterns. Mixtures containing grasses with strong autumn and winter growth, such as D. glomerata, ×Festulolium, and F. arundinacea, and cool-season herbs such as P. lanceolata, exhibited higher productivity during those periods. These results emphasize the importance of tailoring pasture mixtures to specific environmental conditions and management goals to maximize productivity throughout the year. For example, MSS3 exhibited a 15.8% higher production rate compared to LP-TR in summer, whereas MSS2 had a 30.9% higher production rate in winter compared to LP-TR, reflecting the contribution of P. lanceolata in this period.
Regarding defoliation management, a shorter rotation length generally resulted in higher forage production across all mixtures, except for winter. This suggests that more frequent defoliation, allowing plants to remain in a vegetative state, can enhance overall productivity—an approach that may also improve the nutritional value of the pasture. Vegetative forage generally maintains higher crude protein content and lower fiber levels, thus increasing digestibility and animal intake [26,34]. However, it is crucial to acknowledge potential trade-offs associated with this management strategy. More frequent defoliation could compromise the long-term persistence of desirable species, particularly those with slower regrowth rates or limited capacity to restore WSC reserves to pre-defoliation levels in short RL [30,32]. Moreover, the impact of RL on forage quality and unsown species dynamics warrants further investigation.
The observation that DI did not have a significant impact on DM yield during winter and summer, but that the greatest growth rates were achieved during spring and autumn, indicates the importance of considering species-specific traits and seasonal growth patterns when designing defoliation strategies. Interestingly, a DI of 8 cm resulted in greater DM accumulation compared to the 5 cm treatment. Leaving a higher residual leaf area likely enhanced photosynthetic capacity and accelerated regrowth rates [3,27], leading to greater overall DM production. However, it’s important to consider species-specific responses to defoliation intensity. Some species may exhibit greater tolerance to close defoliation, while others may require a higher residual height to maintain productivity and persistence [35,36]. However, the effect of DI on DM production is considerably smaller than that observed when evaluating different RL, suggesting that defoliation interval exerts a more significant influence on annual and seasonal forage production.
The significant interaction effects observed between RL and DI underscore the complexity of pasture management and the need for adaptive grazing strategies, which should be tailored to both the species composition and the season. Notably, the synergistic effect of the 8 cm/short RL combination on annual and spring herbage DM production emphasizes the importance of balancing RL and DI to maximize pasture productivity. This defoliation management likely reflects the greater ability of plants to quickly recover and capitalize on regrowth opportunities afforded by the reduced intervals between cuts when combined with a less severe DI. This suggests a physiological shift towards prioritizing rapid leaf area recovery from remaining photosynthetic tissues, as opposed to relying primarily on WSC remobilized from storage. Our results further demonstrate that MSS pastures consistently exhibited higher daily growth rates compared to LP-TR during the winter and autumn months, due to a greater proportion of winter-growing grasses such as F. arundinacea, D. glomerata, and ×Festutolium, highlighting the importance of selecting appropriate species. Two alternative grazing approaches emerged as effective during winter: defoliation at 5 cm/long RI, and 8 cm/short RL. Both strategies aim to strike a balance between forage utilization and pasture persistence.
4.2. Botanical Composition
Pasture botanical composition is critical to both productivity and resilience, and is strongly influenced by defoliation management and the characteristics of the component species. Seasonal conditions and defoliation management, such as RL and DI, are recognized as key factors influencing the botanical composition of pastures by favoring certain forage species over others [28]. In our study, the seasonal changes had a marked effect on the botanical composition of sown pastures, representing the main driver in determining the relative abundance of individual species within the mixtures. These results reflect the differential growth responses and competitive abilities of each species to temperature, photoperiod, and moisture availability throughout the year [30].
In LP-TR, L. perenne exhibited strong seasonal fluctuations, with greater contributions during winter and autumn and reduced contributions during spring and summer. This decline is likely attributable to its shallow root system and low tolerance to heat and drought stress, which limit its competitiveness under dry and warm conditions [37,38]. Furthermore, L. perenne allocates its photoassimilates towards rapid regrowth of leaf area post-defoliation or post-grazing, potentially at the expense of root development, thus exacerbating its vulnerability under soil moisture deficits [39]. Conversely, T. repens and, particularly, unsown species (primarily, Erodium moschatum (L.) L’Hér., Spergula arvensis L., Polygonum aviculare L., and Cynodon dactylon (L.) Pers.), showed increased contributions during late spring and summer. This shift may indicate a release from competition due to L. perenne decline, enabling these more stress-tolerant species to exploit open niches [40].
The MSS pastures exhibited more complex botanical dynamics characterized by both competitive interactions and functional complementarity [15,28]. Plantago lanceolata emerged as a key driver of seasonal productivity, particularly in autumn and winter, demonstrating its adaptability to a range of environmental conditions and its ability to maintain productivity during periods when other species may be less active [41,42]. However, its dominance fluctuated significantly across seasons, with a notable decline in spring, especially in MSS3, where L. perenne temporarily gained competitive ground. The varying proportions of P. lanceolata and other species, such as D. glomerata, ×Festulolium, and F. arundinacea, contributed to distinct seasonal patterns. Seasonal patterns reveal distinct temporal niche partitioning, where different species dominate at different times throughout the year [15]. This study confirms that MSS pastures maintained a more stable botanical composition with fewer unsown species compared to LP-TR pastures, regardless of residual height. The greater diversity in MSS pastures, with functional group complementarity contributing to overall system stability and resilience [28], emphasizes a vital trait to consider when thinking of long-term persistence of a pasture.
Regarding management effects, rotation length (RL) exerted a surprisingly minor influence on species composition compared to defoliation intensity (DI), especially when compared to the strong influence of seasonal changes. This contrasts with earlier research comparing the effect of DI and RL on chemical and P. lanceolata and or C. intybus [26,27,32], which suggested that longer RL tends to benefit species with deeper roots and slower establishment rates [28]. Similarly, defoliation intensity (DI) had only minor impacts, suggesting that climatic conditions and interspecific interactions exert stronger control over community composition than the short-term effects of defoliation regimes under the tested conditions.
The significant interaction between DI and RL on botanical composition highlights the complex interactions between management and species dynamics in more diverse pastures. When increasing the percentage of grasses, the interaction of DI × RL becomes more evident, resulting in management being vital for the balance of composition.
4.3. Management Implications
This study provides valuable insights into optimizing pasture management strategies for temperate grazing systems, underscoring the importance of considering both pasture composition and defoliation management to enhance forage production, maintain sward stability, and promote long-term sustainability. Higher herbage DM production of MSS pastures over LP-TR highlights the benefits of incorporating a diverse range of species with complementary traits.
Implementing shorter rotation lengths (i.e., more frequent grazing) can enhance forage yield, although careful monitoring is necessary to ensure the persistence of desirable species. While a defoliation intensity of 8 cm generally resulted in greater DM accumulation, seasonal adjustments may be warranted. This adaptive approach requires careful observation of pasture growth and species composition throughout the year. The significant interactions observed between RL and DI highlight the need for customized management strategies based on the specific pasture composition. For instance, in MSS pastures, a combination of shorter RL and 8 cm residual height may be optimal for maximizing forage production, while, in LP-TR pastures, a longer RL may be necessary to maintain L. perenne persistence. The varying productive response of multispecies pastures to the defoliation strategy throughout the production season highlights the importance that defoliation management in this type of pasture should be adaptive, relative to the dominant species in the botanical composition, according to the different seasons of the year. Regular monitoring of botanical composition is essential to detect shifts in species dominance and to adjust management practices accordingly.
While our study provides valuable insights into short-term responses to different management practices in MSS pastures, long-term trials are needed to assess the sustainability of these effects. This research provides a valuable framework for developing management guidelines that are adapted to specific pasture compositions and management goals, ultimately contributing to more resilient and profitable temperate grazing systems.
5. Conclusions
Our findings demonstrate that multispecies pastures enhance productivity in temperate grazing systems relative to traditional ryegrass–white clover mixtures. The manipulation of defoliation strategies, particularly by reducing rotation length while being mindful of residual sward height, significantly influenced forage yield and botanical composition. These results emphasize the value of adaptive grazing management approaches that consider species composition and seasonal variations to optimize forage production, promote pasture stability, and increase the resilience and persistence of temperate pastures, ultimately benefiting sustainable livestock production systems.
Author Contributions
Conceptualization, L.F.P., V.M.M., C.M.F., and M.J.N.; methodology, L.F.P., V.M.M., and C.M.F.; validation, L.F.P., and V.M.M.; formal analysis, L.F.P., C.L., G.S., A.A., T.S., and M.J.N.; investigation, C.L., G.S., A.A., and T.S.; resources, L.F.P., V.M.M., C.M.F., and M.J.N.; data curation, L.F.P., M.J.N., C.L., G.S., A.A., and T.S.; writing—original draft preparation, L.F.P., V.M.M., and M.J.N.; writing—review and editing, L.F.P., and V.M.M.; visualization, L.F.P., and V.M.M.; supervision, C.M.F., M.J.N., and L.F.P.; project administration, M.J.N., C.M.F., L.F.P., and V.M.M.; funding acquisition, L.F.P., V.M.M., and C.M.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Agencia Nacional de Investigación y Desarrollo (ANID), Chile, SIA/IDeA I+D grant number ID23I10191.
Data Availability Statement
The data presented in this study are available on request from the corresponding author, because they are part of an ongoing study.
Acknowledgments
The authors wish to thank D. Figueroa, C. Madrid, O. Coronado, and B. Sandoval for their technical assistance. The authors gratefully acknowledge the support of the staff at the Oromo Experimental Station for their assistance in maintaining the experimental area.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hidosa, D.; Guyo, M. Climate change effects on livestock feed resources: A review. J. Fish. Livest. Prod. 2017, 5, 259. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014—Impacts, Adaptation and Vulnerability. Part A: Global and Sectoral Aspects. In Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change 2014; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Lee, J.M.; Matthew, C.; Thom, E.R.; Chapman, D.F. Perennial ryegrass breeding in New Zealand: A dairy industry perspective. Crop Pasture Sci. 2012, 63, 107–127. [Google Scholar] [CrossRef]
- Horan, B.; Roche, J. Defining resilience in pasture-based dairy-farm systems in temperate regions. Anim. Prod. Sci. 2020, 60, 55–66. [Google Scholar] [CrossRef]
- Goh, K.M.; Bruce, G.E. Comparison of biomass production and biological nitrogen fixation of multi-species pastures (mixed herb leys) with perennial ryegrass-white clover pasture with and without irrigation in Canterbury, New Zealand. Agric. Ecosyst. Environ. 2005, 110, 230–240. [Google Scholar] [CrossRef]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of legume-based grassland–livestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef]
- Herrero, M.; Henderson, B.; Havlík, P.; Thornton, P.K.; Conant, R.T.; Smith, P.; Wirsenius, S.; Hristov, A.N.; Gerber, P.; Gill, M.; et al. Greenhouse gas mitigation potentials in the livestock sector. Nat. Clim. Change 2016, 6, 452–461. [Google Scholar] [CrossRef]
- Ghahramani, A.; Howden, S.M.; del Prado, A.; Thomas, D.T.; Moore, A.D.; Ji, B.; Ates, S. Climate change impact, adaptation, and mitigation in temperate grazing systems: A review. Sustainability 2019, 11, 7224. [Google Scholar] [CrossRef]
- Rivera-Ferre, M.G.; López-i-Gelats, F.; Howden, M.; Smith, P.; Morton, J.F.; Herrero, M. Re-framing the climate change debate in the livestock sector: Mitigation and adaptation options. Rev. Clim. Change 2016, 7, 869–892. [Google Scholar] [CrossRef]
- Jordon, M.W.; Willis, K.J.; Bürkner, P.C.; Petrokofsky, G. Rotational grazing and multispecies herbal leys increase productivity in temperate pastoral systems—A meta-analysis. Agric. Ecosyst. Environ. 2022, 337, 108075. [Google Scholar] [CrossRef]
- Jing, J.; Søegaard, K.; Cong, W.F.; Eriksen, J. Species diversity effects on productivity, persistence and quality of multispecies swards in a four-year experiment. PLoS ONE 2017, 12, e0169208. [Google Scholar] [CrossRef] [PubMed]
- Hearn, C.; Egan, M.; Lynch, M.B.; Flynn, D.; O’Donovan, M. Can the Inclusion of Ribwort Plantain or Chicory Increase the Seasonal and Annual Dry Matter Production of Intensive Dairy Grazing Swards? Eur. J. Agron. 2024, 152, 127020. [Google Scholar] [CrossRef]
- Finn, J.A.; Kirwan, L.; Connolly, J.; Sebastiá, M.T.; Helgadottir, A.; Baadshaug, O.H.; Bélanger, G.; Black, A.; Brophy, C.; Collins, R.; et al. Ecosystem function enhanced by combining four functional types of plant species in intensively managed grassland mixtures: A 3-year continental-scale field experiment. J. Appl. Ecol. 2013, 50, 365–375. [Google Scholar] [CrossRef]
- Malisch, C.S.; Finn, J.A.; Eriksen, J.; Loges, R.; Brophy, C.; Huguenin-Elie, O. The importance of multi-species grassland leys to enhance ecosystem services in crop rotations. Grass Forage Sci. 2024, 79, 120–134. [Google Scholar] [CrossRef]
- Baker, S.; Lynch, M.B.; Godwin, F.; Boland, T.M.; Kelly, A.K.; Evans, A.; Murphy, P.; Sheridan, H. Multispecies swards outperform perennial ryegrass monoculture and perennial ryegrass-white clover swards—A beef system grazing experiment. SSRN Electron. J. 2022. [Google Scholar] [CrossRef]
- Jaramillo, D.M.; Sheridan, H.; Soder, K.; Dubeux, J.C.B. Enhancing the Sustainability of Temperate Pasture Systems through More Diverse Swards. Agronomy 2021, 11, 1912. [Google Scholar] [CrossRef]
- Ordoñez, I.P.; López, I.F.; Kemp, P.D.; Donaghy, D.J.; Dörner, J.; García-Favre, J.; Zhang, Y. A short-term effect of multi-species pastures and the plant’s physiological response on pasture growth. Eur. J. Agron. 2024, 159, 127232. [Google Scholar] [CrossRef]
- Teixeira, A.; Toorop, P.E.; Iannetta, P.P.M. Differential Interspecific Adaptation to Abiotic Stress by Plantago Species. Front. Plant Sci. 2020, 11, 573039. [Google Scholar] [CrossRef]
- Hoekstra, N.J.; Suter, M.; Finn, J.A.; Husse, S.; Lüscher, A. Do belowground vertical niche differences between deep- and shallow-rooted species enhance resource uptake and drought resistance in grassland mixtures? Plant Soil 2015, 394, 21–34. [Google Scholar] [CrossRef]
- Chesney, L.E.; Carnovale, F.; Huson, K.M.; Rutherford, N.; Patterson, D. Plantain (Plantago lanceolata L.) as an Alternative Forage to Build Resilience and Reduce the Environmental Footprint of Grazing Dairy Systems in Temperate Northern Climates: A Review. Sustainability 2025, 17, 3131. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Navarrete, S.; Horne, D.J.; Donaghy, D.J.; Kemp, P.D. Forage plantain (Plantago lanceolata L.): Meta-analysis quantifying the decrease in nitrogen excretion, the increase in milk production, and the changes in milk composition of dairy cows grazing pastures containing plantain. Anim. Feed. Sci. Technol. 2022, 285, 115244. [Google Scholar] [CrossRef]
- Della Rosa, M.M.; Sandoval, E.; Luos, D.; Pacheco, D.; Jonker, A. Effect of feeding fresh forage plantain (Plantago lanceolata) or ryegrass-based pasture on methane emissions, total-tract digestibility, and rumen fermentation of nonlactating dairy cows. J. Dairy Sci. 2022, 105, 6628–6638. [Google Scholar] [CrossRef] [PubMed]
- Distel, R.A.; Arroquy, J.I.; Lagrange, S.; Villalba, J.J. Designing diverse agricultural pastures for improving ruminant production systems. Front. Sustain. Food Syst. 2020, 4, 215. [Google Scholar] [CrossRef]
- Darch, T.; Mcgrath, S.P.; Lee MR, F.; Beaumont, D.A.; Blackwell MS, A.; Horrocks, C.A.; Storkey, J. The Mineral Composition of Wild-Type and Cultivated Varieties of Pasture Species. Agronomy 2020, 10, 1463. [Google Scholar] [CrossRef]
- Grace, C.; Lynch, M.B.; Sheridan, H.; Lott, S.; Fritch, R.; Boland, T.M. Grazing multispecies swards improves ewe and lamb performance. Animal 2019, 13, 1721–1729. [Google Scholar] [CrossRef] [PubMed]
- Merino, V.M.; Aguilar, R.; Piña, L.F.; Garriga, M.; Ostria-Gallardo, E.; López, M.D.; Noriega, F.; Campos, J.; Navarrete, S.; Rivero, M.J. Regrowth dynamics and morpho-physiological characteristics of Plantago lanceolata under different defoliation frequencies and intensities. PLoS ONE 2024, 19, e0310009. [Google Scholar] [CrossRef]
- Lee, J.M.; Hemmingson, N.R.; Minnee, E.M.; Clark, C.E. Management strategies for chicory (Cichorium intybus) and plantain (Plantago lanceolata): Impact on dry matter yield, nutritive characteristics and plant density. Crop Pasture Sci. 2015, 66, 168–183. [Google Scholar] [CrossRef]
- Baker, S.; Lynch, M.; Godwin, F.; Brennan, E.; Boland, T.; Evans, A.; Kelly, A.; Sheridan, H. Dry-matter production and botanical composition of multispecies and perennial ryegrass swards under varying defoliation management. Grass Forage Sci. 2023, 78, 390–401. [Google Scholar] [CrossRef]
- Nobilly, F.; Bryant, R.H.; McKenzie, B.A.; Edwards, G.R. Productivity of rotationally grazed simple and diverse pasture mixtures under irrigation in Canterbury. Proc. N. Z. Grassl. Assoc. 2013, 75, 165–172. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Soder, K.J.; Muller, L.D.; Klement, K.D.; Skinner, R.H.; Goslee, S.C. Forage Mixture Productivity and Botanical Composition in Pastures Grazed by Dairy Cattle. Agronomy 2005, 97, 1465–1471. [Google Scholar] [CrossRef]
- CIREN. Estudio Agrológico X Región; Publicación CIREN N° 123; Centro de Información de Recursos Naturales, Gobierno de Chile: Santiago, Chile, 2003; p. 136. [Google Scholar]
- Merino, V.M.; Aguilar, R.I.; Rivero, M.J.; Ordóñez, I.P.; Piña, L.F.; López-Belchí, M.D.; Schoebitz, M.I.; Noriega, F.A.; Pérez, C.I.; Cooke, A.S.; et al. Distribution of Non-Structural Carbohydrates and Root Structure of Plantago lanceolata L. under Different Defoliation Frequencies and Intensities. Plants 2024, 13, 2773. [Google Scholar] [CrossRef]
- Tozer, K.N.; Cameron, C.A.; Thom, E.R. Pasture persistence: Farmer observations and field measurements. NZGA Res. Pract. Ser. 2011, 15, 25–30. [Google Scholar] [CrossRef]
- Merino, V.M.; Balocchi, O.A.; Pulido, R. Effect of daily herbage allowance restriction on pasture characteristics and milk production by grazing dairy cows in spring. Cienc. Investig. Agrar. 2018, 45, 21–24. [Google Scholar] [CrossRef]
- Phelan, P.; Casey, I.A.; Humphreys, J. The effect of target postgrazing height on sward clover content, herbage yield, and dairy production from grass-white clover pasture. J. Dairy Sci. 2013, 96, 1598–1611. [Google Scholar] [CrossRef]
- Kaufononga, S.; Donaghy, D.J.; Hendriks, S.J.; Matthew, C.; Kemp, P.D.; Cranston, L.M. Comparative response of tall fescue and perennial ryegrass swards to variation in defoliation interval and height. N. Z. J. Agric. Res. 2017, 60, 363–375. [Google Scholar] [CrossRef]
- Wims, C.M.; McEvoy, M.; Delaby, L.; Boland, T.M.; O’Donovan, M. Effect of perennial ryegrass (Lolium perenne L.) cultivars on the milk yield of grazing dairy cows. Animal 2013, 7, 410–421. [Google Scholar] [CrossRef]
- Franzluebbers, A.; Seman, D.; Stuedemann, J. Forage dynamics in mixed tall fescue–bermudagrass pastures of the Southern Piedmont USA. Agric. Ecosyst. Environ. 2013, 168, 37–45. [Google Scholar] [CrossRef]
- Donaghy, D.J.; Fulkerson, W.J. The importance of water-soluble carbohydrate reserves on regrowth and root growth of Lolium perenne (L.). Grass Forage Sci. 1997, 52, 401–407. [Google Scholar] [CrossRef]
- Hernandez, P.; Picon-Cochard, C. Presence of Trifolium repens Promotes Complementarity of Water Use and N Facilitation in Diverse Grass Mixtures. Front. Plant Sci. 2016, 6, 538. [Google Scholar] [CrossRef] [PubMed]
- Cranston, L.; Kenyon, P.; Morris, S.; Lopez-Villalobos, N.; Kemp, P. Ewe lamb diet selection on plantain (Plantago lanceolata) and on a herb and legume mix, including plantain, chicory (Cichorium intybus), red clover (Trifolium pratense) and white clover (Trifolium repens). Anim. Prod. Sci. 2015, 55, 515. [Google Scholar] [CrossRef]
- Nie, Z.; Miller, S.; Moore, G.; Hackney, B.; Boschma, S.; Reed, K.; Mitchell, M.; Albertsen, T.; Clark, S.; Craig, A. Field evaluation of perennial grasses and herbs in southern Australia. 2. Persistence, root characteristics and summer activity. Aust. J. Exp. Agric. 2008, 48, 424–435. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).