

Enhancing Soil Phosphorus Availability in Intercropping Systems: Roles of Plant Growth Regulators

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
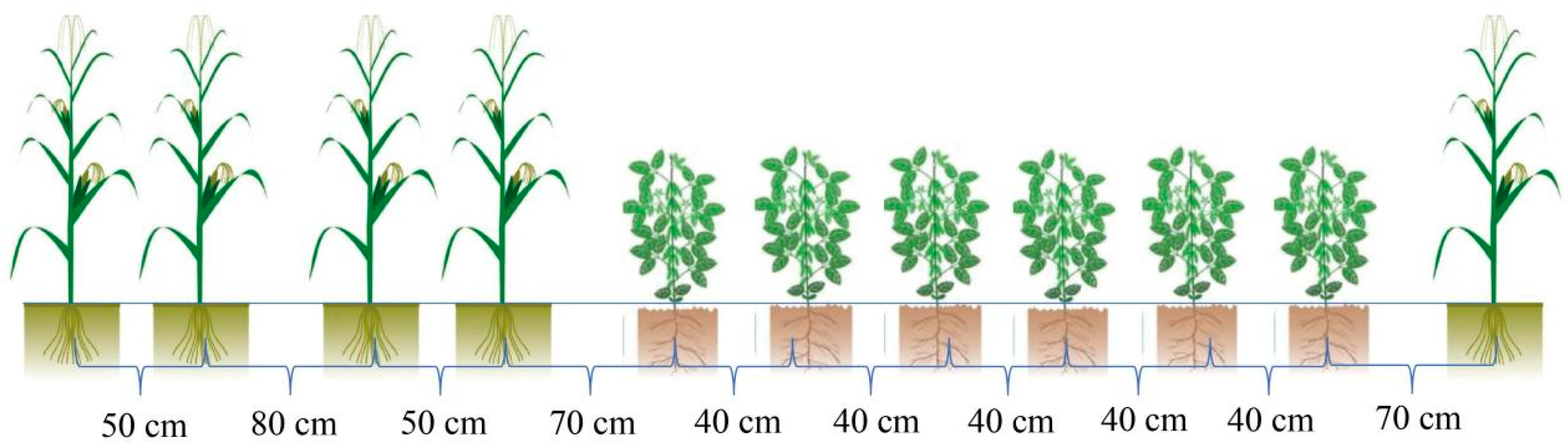
2.2. Experimental Design
2.3. Soil Sample Collection and Analysis
2.4. Library Preparation and Sequencing
2.5. Gene Prediction, Taxonomy, and Functional Annotation
2.6. Metagenomic Analysis
3. Results
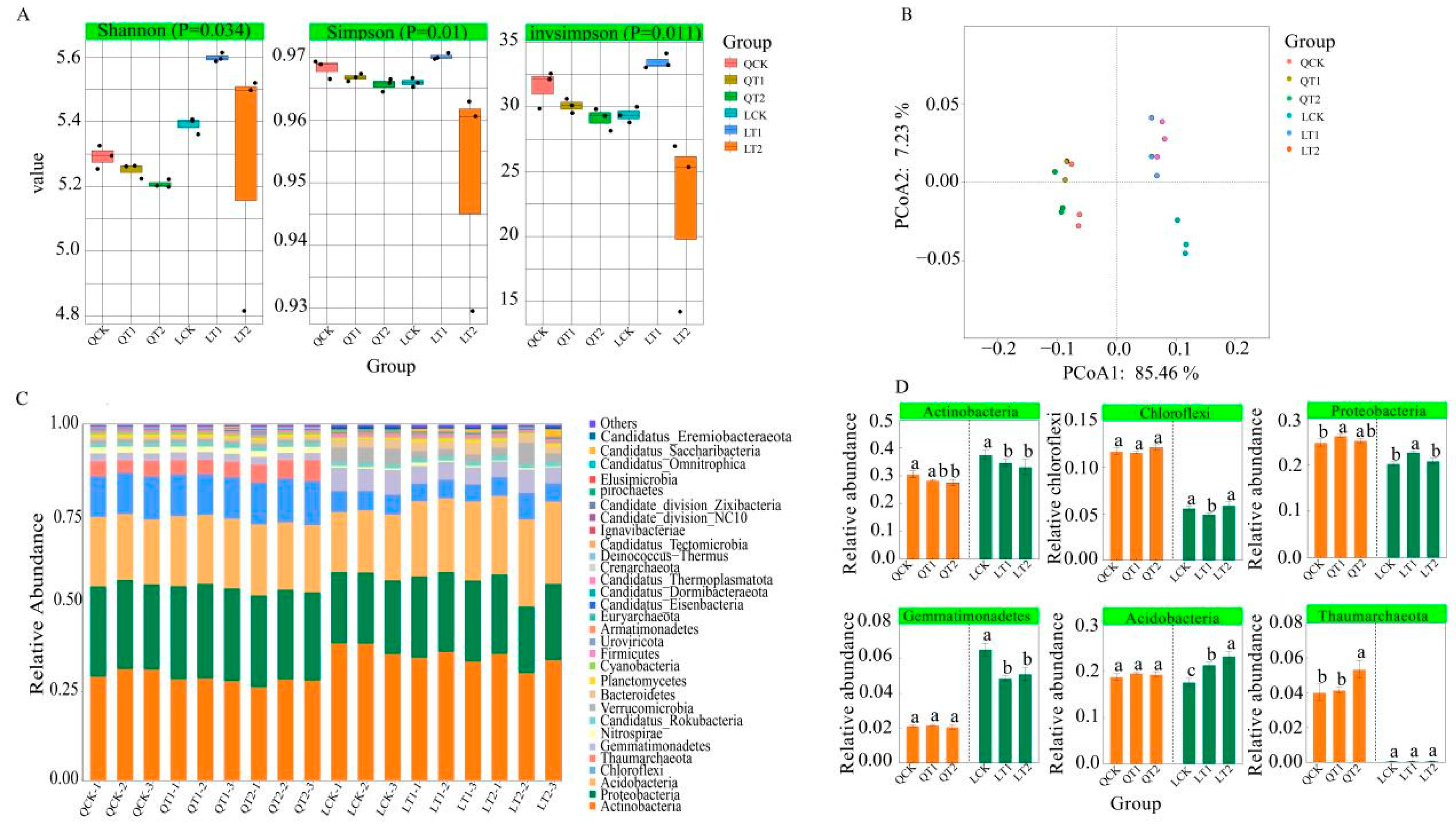
3.1. Soil Phosphorus Contents in Maize Strip in the Intercropping System
3.2. Analysis of Soil Samples from Maize Strip
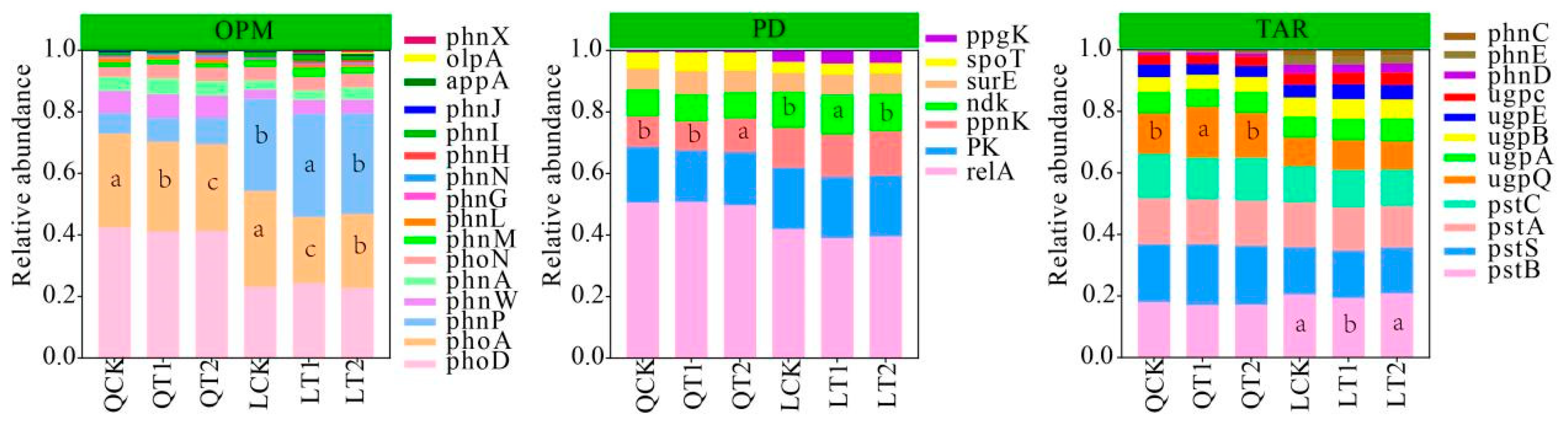
3.3. Functional Gene Abundances in Soil from Maize Strip
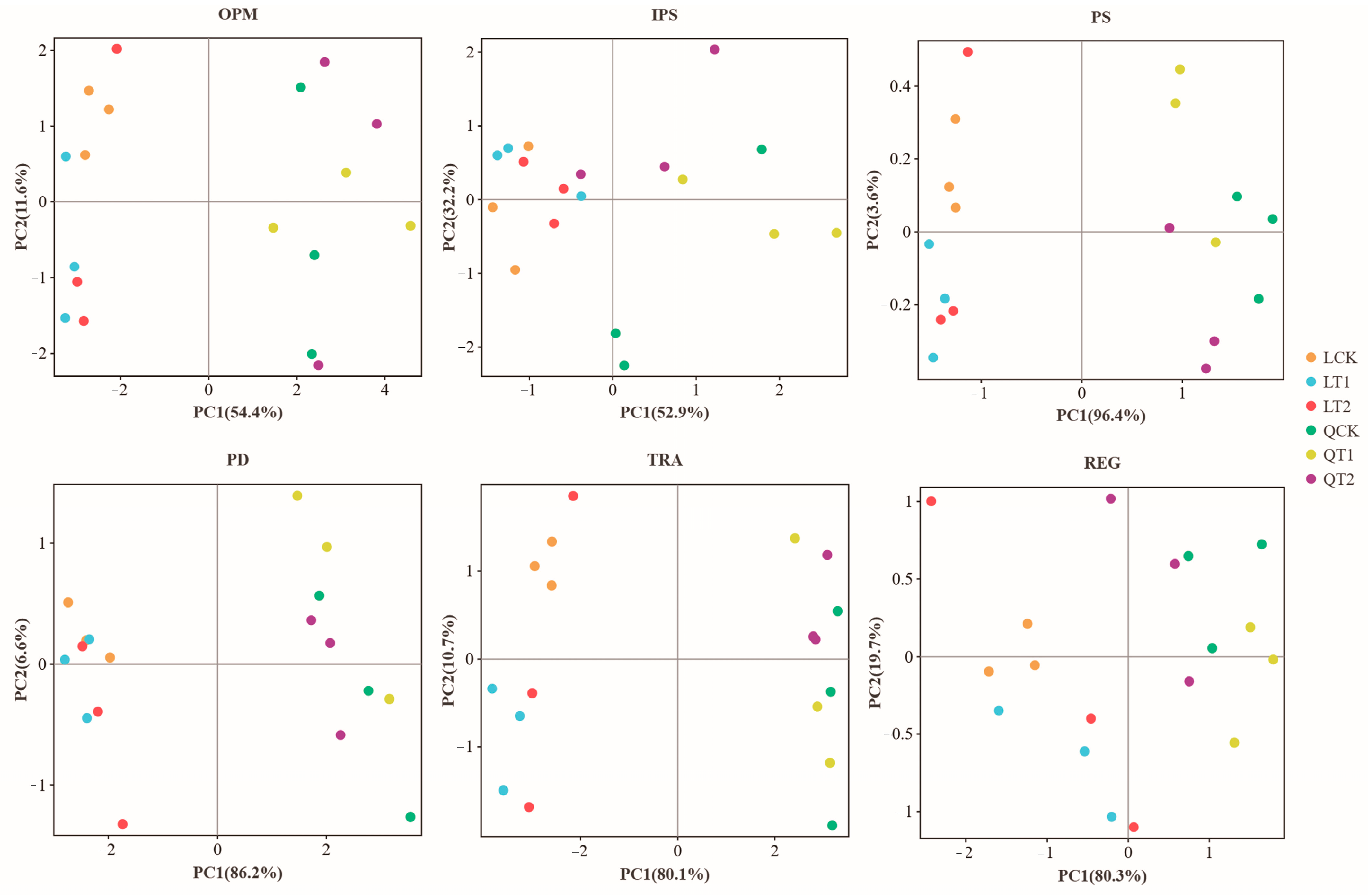
3.4. Contributions of Microorganisms in Maize Strip to Phosphorus Cycle Genes
3.5. Differences in Microbial Communities and Relationships with Phosphorus Cycling Functional Genes and Soil Phosphorus Availability
4. Discussion
4.1. Effects of Growth Regulators on Soil Phosphorus Contents in the Intercropping System
4.2. Effects of Growth Regulators on Soil Phosphorus Cycling Microbial Community
4.3. Effects of PGRs on Soil Phosphorus Cycling Genes
4.4. Differences in Contributions of Growth Regulators to Phosphorus Cycle Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Wang, K.Y.; Onodera, S.I.; Saito, M.; Ishida, T. Assessment of long-term phosphorus budget changes influenced by anthropogenic factors in a coastal catchment of Osaka Bay. Sci. Total Environ. 2022, 843, 156833. [Google Scholar] [CrossRef] [PubMed]
- An, X.X.; Liu, J.; Liu, X.S.; Ma, C.; Zhang, Q. Optimizing phosphorus application rate and the mixed inoculation of arbuscular mycorrhizal fungi and phosphate solubilizing bacteria can improve the phosphatase activity and organic acid content in Alfalfa Soil. Sustainability 2022, 14, 11342. [Google Scholar] [CrossRef]
- Demay, J.; Ringeval, B.; Pellerin, S.; Nesme, T. Half of global agricultural soil phosphorus fertility derived from anthropogenic sources. Nat. Geosci. 2023, 16, 69–74. [Google Scholar] [CrossRef]
- Zhu, X.H.; Tan, J.L.; Zhou, H.Y.; Wang, T.Q.; Zhang, B.B.; Lu, X.; Tian, J.H.; Liang, C.Y.; Tian, J. Effects of different genotypes soybean and maize intercropping on soil phosphorus fractions and crop phosphorus uptake. J. Appl. Ecol. 2024, 35, 1583–1589. [Google Scholar]
- Nasar, J.; Ahmad, M.; Gitari, H.; Tang, L.; Chen, Y.; Zhou, X.B. Maize/soybean intercropping increases nutrient uptake, crop yield and modifies soil physio-chemical characteristics and enzymatic activities in the subtropical humid region based in Southwest China. BMC Plant Biol. 2024, 24, 434. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.Y.; He, J.; Zhou, Z.H.; Xia, L.L.; Hu, Y.F.; Zhang, Y.L.; Zhang, Y.Y.; Luo, Y.Q.; Chu, H.Y.; Liu, W.J.; et al. Organic amendments enhance soil microbial diversity, microbial functionality and crop yields: A meta-analysis. Sci. Total Environ. 2022, 829, 154627. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.C.; Wang, W.M.; Wang, Z.; Li, L.H.; Jiang, G.J.; Wang, J.P.; Cheng, Z.B. Effects of maize and soybean intercropping on soil phosphorus bioavailability and microbial community structure in rhizosphere. J. Appl. Ecol. 2023, 34, 3030–3038. [Google Scholar]
- Raseduzzaman, M.; Jensen, E.S. Does intercropping enhance yield stability in arable crop production? A meta-analysis(Article). Eur. J. Agron. 2017, 91, 25–33. [Google Scholar] [CrossRef]
- Bedoussac, L.; Journet, E.P.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.; Prieur, L.; Justes, E. Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Han, C.Q.; Yang, D.L.; Yang, J.J.; Cade-Menun, B.J.; Chen, Y.Q.; Sui, P. Maize-soybean intercropping facilitates chemical and microbial transformations of phosphorus fractions in a calcareous soil. Front. Microbiol. 2022, 13, 1028969. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.; Tanaka, M.; Sakai, M.; Toyama, H.; Matsushita, K.; Adachi, O.; Yamada, M. C-terminal periplasmic domain of escherichia coli quinoprotein glucose dehydrogenase transfers electrons to ubiquinone. J. Biol. Chem. 2001, 276, 48356–48361. [Google Scholar] [CrossRef] [PubMed]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.; Li, H.; Lakshmanan, P.; Chen, Y.; Chen, X. phoD-harboring bacterial community composition dominates organic P mineralization under long-term P fertilization in acid purple soil. Front. Microbiol. 2022, 13, 1045919. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Liu, G.; Chen, H.; Chen, C.; Wang, J.; Ai, S.; Wei, D.; Li, D.; Ma, B.; Tang, C.; et al. Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. ISME J. 2020, 14, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Rensing, C.; Han, D.F.; Xiao, K.Q.; Dai, Y.X.; Tang, Z.X.; Liesack, W.; Peng, J.J.; Cui, Z.L.; Zhang, F.S. Genome-resolved metagenomics reveals distinct phosphorus acquisition strategies between soil microbiomes. mSystems 2022, 7, e0110721. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.J.; Wanner, B.L. Global regulation by the seven-component Pi signaling system. Curr. Opin. Microbiol. 2010, 13, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Enebe, M.C.; Babalola, O.O. The influence of soil fertilization on the distribution and diversity of phosphorus cycling genes and microbes community of maize rhizosphere using shotgun metagenomics. Genes 2021, 12, 1022. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, F.; Shao, M.; Huang, L.; Xie, Y.; Xu, Y.; Kong, L. Effects of rotations with legume on soil functional microbial communities involved in phosphorus transformation. Front. Microbiol. 2021, 12, 661100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, S.; Meng, L.; Liu, X.; Zhang, Y.; Zhao, S.; Zhao, H. Root exudation under maize/soybean intercropping system mediates the arbuscular mycorrhizal fungi diversity and improves the plant growth. Front. Plant Sci. 2024, 15, 1375194. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Wang, W.; Gan, Y.; Wang, L.; Chang, X.; Wang, Y.; Yang, W. Growth promotion ability of phosphate-solubilizing bacteria from the soybean rhizosphere under maize-soybean intercropping systems. JSFA Rep. 2022, 102, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Mir, R.A.; Hussain, S.J.; Prasad, B.; Kumar, P.; Aloo, B.N.; Sharma, C.M.; Dubey, R.C. Prospects of phosphate solubilizing microorganisms in sustainable agriculture. World J. Microbiol. Biotechnol. 2024, 40, 291. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Q.; Liu, W.; Li, R.; Zhang, D.; Zhang, G.; Xu, Y. Plant secretions and volatiles contribute to the evolution of bacterial antibiotic resistance in soil-crop system. J. Environ. Sci. 2025, 152, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Alzate Zuluaga, M.Y.; Fattorini, R.; Cesco, S.; Pii, Y. Plant-microbe interactions in the rhizosphere for smarter and more sustainable crop fertilization: The case of PGPR-based biofertilizers. Front. Microbiol. 2024, 15, 1440978. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; Pant, P.; Pant, B.D.; Krom, N.; Allen, R.D.; Mysore, K.S. Elucidating the unknown transcriptional responses and PHR1-mediated biotic and abiotic stress tolerance during phosphorus limitation. J. Exp. Bot. 2023, 74, 2083–2111. [Google Scholar] [CrossRef] [PubMed]
- Akond, Z.; Hasan, M.N.; Alam, M.J.; Alam, M.; Mollah, M.N.H. Classification of functional metagenomes recovered from different environmental samples. Bioinformation 2019, 15, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Kariya, S.; Kurokawa, J. Efficient PCR-Based amplification of diverse alcohol dehydrogenase genes from metagenomes for improving biocatalysis: Screening of gene-specific amplicons from metagenomes. Appl. Environ. Microbiol. 2014, 80, 6280–6289. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.N.S.; Sharma, P.; Maurya, S.; Yadav, R.K. Metagenomics and metatranscriptomics as potential driving forces for the exploration of diversity and functions of micro-eukaryotes in soil. 3 Biotech 2023, 13, 423. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.H.; Jiang, Y.H.; Han, W.W.; Shi, L.G.; Zhang, Y.R.; Liu, G.Z.; Cui, Y.H.; Du, X.; Gao, Z.; Liang, X.G. Simultaneous enhancement of maize yield and lodging resistance via delaying plant growth retardant application. Field Crops Res. 2024, 317, 109530. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 3 Chemical Methods; Wiley: New York, NY, USA, 1996; Volume 5.3, pp. 961–1010. [Google Scholar]
- Chen, M.M.; Zhang, S.R.; Liu, L.; Wu, L.P.; Ding, X.D. Combined organic amendments and mineral fertilizer application increase rice yield by improving soil structure, P availability and root growth in saline-alkaline soil. Soil Tillage Res. 2021, 212, 105060. [Google Scholar] [CrossRef]
- Roberts, T.; Ross, W.; Norman, R.; Slaton, N.; Wilson, C. Predicting nitrogen fertilizer needs for Rice in Arkansas using alkaline hydrolyzable-nitrogen. Soil Sci. Soc. Am. J. 2011, 75, 1161–1171. [Google Scholar] [CrossRef]
- Heinen, B.J.; Vaz, J.E.; Benzo, Z.; Mejias, C. A comparison of extraction and suspension methods for determining exchangeable potassium in soils. Appl. Clay Sci. 1999, 14, 245–255. [Google Scholar] [CrossRef]
- Bao, S. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 79–80. [Google Scholar]
- Wang, H.; Chen, J.P.; Ruan, Y.H.; Sun, W.; Wang, S.L.; Wang, H.T.; Zhang, Y.L.; Guo, J.M.; Wang, Y.C.; Guo, H.Y.; et al. Metagenomes reveal the effect of crop rotation systems on phosphorus cycling functional genes and soil phosphorus avail–ability. Agric. Ecosyst. Environ. 2024, 364, 108886. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Peng, J.; Liu, P.; Bei, Q.; Rensing, C.; Li, Y.; Yuan, H.; Liesack, W.; Zhang, F.; Cui, Z. Metagenomic insights into nitrogen and phosphorus cycling at the soil aggregate scale driven by organic material amendments. Sci. Total Environ. 2021, 785, 147329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Xu, J.; Nadia, R.; Jin, T.; Li, J.Y.; Wang, N. Huanglongbing impairs the rhizosphere-to-rhizoplane enrichment process of the citrus root-associated microbiome. Microbiome 2017, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Yang, C.; Halitschke, R.; Paetz, C.; Kessler, D.; Burkard, K.; Gaquerel, E.; Baldwin, I.T.; Li, D. Natural history-guided omics reveals plant defensive chemistry against leafhopper pests. Science 2022, 375, eabm2948. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Mou, T.; Sun, Y.; Su, J.; Yu, L.; Zhang, Y. Environmental distribution and genomic characteristics of Solirubrobacter, with proposal of two novel species. Front. Microbiol. 2023, 14, 1267771. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y. Search for natural products from actinomycetes of the genus Nocardia. J. Nat. Med. 2024, 78, 828–837. [Google Scholar] [CrossRef] [PubMed]
- He, M.Y.; Peng, S.; Zhang, J.H.; Wang, Y.M.; Hua, Q.Q.; Cheng, S.Q. The type and degree of salinized soils together shape the composition of phoD-harboring bacterial communities, thereby altering the effectiveness of soil phosphorus cycling. J. Environ. Manag. 2025, 385, 125621. [Google Scholar] [CrossRef] [PubMed]
- Namuli, A.; Patrick, E.O.; John, B.T.; Muwanika, V.B.; Johnson, M.; Bazira, J. Diversity of bacterial community in the rhizosphere and bulk soil of Artemisia annua grown in highlands of Uganda. PLoS ONE 2023, 18, e0269662. [Google Scholar]
- Tian, J.H.; Kuang, X.Z.; Tang, M.T.; Chen, X.D.; Huang, F.; Cai, Y.X.; Cai, K.Z. Biochar application under low phosphorus input promotes soil organic phosphorus mineralization by shifting bacterial phoD gene community composition. Sci. Total Environ. 2021, 779, 146556. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.C.; Zhou, Y.; Chen, P.; Zhang, X.N.; Du, Q.; Yang, H.; Wang, X.C.; Yang, F.; Xiao, T.; Li, L.; et al. Maize–legume intercropping promote N uptake through changing the root spatial distribution, legume nodulation capacity, and soil N availability. J. Integr. Agric. 2022, 21, 1755–1771. [Google Scholar] [CrossRef]
- Cuartero, J.; Pascual, J.A.; Vivo, J.M.; Özbolat, O.; Sánchez-Navarro, V.; Egea-Cortines, M.; Zornoza, R.; Mena, M.M.; Garcia, E.; Ros, M. A first-year melon/cowpea intercropping system improves soil nutrients and changes the soil microbial community. Agric. Ecosyst. Environ. 2022, 328, 107856. [Google Scholar] [CrossRef]
- Ma, W.Q.; Ma, L.; Li, J.H.; Wang, F.H.; Sisák, I.; Zhang, F.S. Phosphorus flows and use efficiencies in production and consumption of wheat, rice, and maize in China. Chemosphere 2011, 84, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Alori, E.T.; Glick, B.R.; Babalola, O.G. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Bohannan, B.J. Spatial scaling of microbial biodiversity. Trends Ecol. Evol. 2006, 21, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jason, A.P.; Aymé, S.; Omry, K.; Zhao, J.; Susannah, G.T.; Jeffery, L.D.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar]
- Song, J.S.; Zhang, X.L.; Kong, F.L.; Liu, X.L.; An, W.J.; Li, Y.Y. Effects of biomass conditioner on soil nutrient and microbial community characteristics of alpine desertified grassland in northwest Sichuan, China. J. Appl. Ecol. 2021, 6, 2217–2226. [Google Scholar]
- Zhao, M.L.; Zhao, J.; Yuan, J.; Hale, L.; Wen, T.; Huang, Q.W.; Vivanco, J.M.; Zhou, J.Z.; Kowalchuk, G.A.; Shen, Q.R. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ. 2021, 44, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Mander, C.; Wakelin, S.; Young, S.; Condron, L.; Callaghan, M. Incidence and diversity of phosphate-solubilising bacteria are linked to phosphorus status in grassland soils. Soil Biol. Biochem. 2012, 44, 93–101. [Google Scholar] [CrossRef]
- Timofeeva, A.; Galyamova, M.; Sedykh, S. Prospects for using phosphate-solubilizing microorganisms as natural fertilizers in agriculture. Plants 2022, 11, 2119. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Feng, J.P.; Zhu, X.L.; Zhu, F.; Ke, W.S.; Huang, Y.Y.; Wu, C.; Xu, X.G.; Guo, J.K.; Xue, S.G. Organic acid release and microbial community assembly driven by phosphate-solubilizing bacteria enhance Pb, Cd, and As immobilization in soils remediated with iron-doped hydroxyapatite. J. Hazard. Mater. 2025, 488, 137340. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Feng, L.L.; Huang, Y.; Liang, Y.M.; Pan, F.J.; Zhang, W.; Zhao, Y.; Xiao, Y.X. Planted citrus regulates the community and networks of phoD-harboring bacteria to drive phosphorus availability between Karst and Non-Karst Soils. Microorganisms 2024, 12, 2582. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.Z.; Yuan, Q.H.; Guan, T.; Cai, Y.W.; Liu, E.F.; Li, B.; Wang, Y. Biotic regulation of phoD-encoding gene bacteria on organic phosphorus mineralization in lacustrine sediments with distinct trophic levels. Water Res. 2024, 260, 121980. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Bisht, N.; Ansari, M.M.; Chauhan, P.S. Pseudomonas putida triggers phosphorus bioavailability and P-transporters under different phosphate regimes to enhance maize growth. Plant Physiol. Biochem. 2024, 217, 109279. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Liras, P. Molecular mechanisms of phosphate sensing, transport and signalling in streptomyces and related actinobacteria. Int. J. Mol. Sci. 2021, 22, 1129. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.N.; Downs, D.M. An unexpected role for the periplasmic phosphatase phoN in the salvage of B6 vitamers in salmonella enterica. Appl. Environ. Microbiol. 2021, 87, e02300-20. [Google Scholar] [CrossRef] [PubMed]
- Lidbury, I.D.E.A.; Fraser, T.; Murphy, A.R.J.; Scanlan, D.J.; Bending, G.D.; Jones, A.M.E.; Moore, J.D.; Goodall, A.; Tibbett, M.; Hammond, J.P.; et al. The ‘known’ genetic potential for microbial communities to degrade organic phosphorus is reduced in low-pH soils. MicrobiologyOpen 2017, 6, e00474. [Google Scholar] [CrossRef] [PubMed]
- Skouri-Panet, F.; Benzerara, K.; Cosmidis, J.; Férard, C.; Caumes, G.; De, L.G.; Heulin, T.; Duprat, E. In vitro and in silico evidence of phosphatase diversity in the biomineralizing bacterium ramlibacter tataouinensis. Front. Microbiol. 2018, 8, 2592. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.J.; Xia, Y.H.; Sun, Q.; Liu, K.P.; Chen, X.B.; Ge, T.D.; Zhu, B.L.; Zhu, Z.K.; Zhang, Z.H.; Su, Y.R. Effects of long-term fertilization on phoD-harboring bacterial community in Karst soils. Sci. Total Environ. 2018, 628, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Yu, Z.H.; Yao, Q.; Hu, X.J.; Zhang, W.; Mi, G.; Chen, X.L.; Wang, G.H. Distinct soil bacterial communities in response to the cropping system in a mollisol of northeast China. Appl. Soil Ecol. 2017, 119, 407–416. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, S.; Chen, M.; Fei, C.; Zhang, W.; Li, Y.; Ding, X. Fe-modified biochar combined with mineral fertilization promotes soil organic phosphorus mineralization by shifting the diversity of phoD-harboring bacteria within soil aggregates in saline-alkaline paddy soil. J. Soils Sediments 2023, 23, 619–633. [Google Scholar] [CrossRef]
- Zhang, L.; Niu, J.; Lu, X.; Zhao, Z.; Li, K.; Wang, F.; Zhang, C.; Sun, R. Dosage effects of organic manure on bacterial community assemblage and phosphorus transformation profiles in greenhouse soil. Front. Microbiol. 2023, 14, 1188167. [Google Scholar] [CrossRef] [PubMed]
- Buenemann, E.K. Assessment of gross and net mineralization rates of soil organic phosphorus—A review. Soil Biol. Biochem. 2015, 89, 82–98. [Google Scholar] [CrossRef]
- Pistocchi, C.; Meszaros, E.; Tamburini, F.; Frossard, E.; Bunemann, E.K. Biological processes dominate phosphorus dynamics under low phosphorus availability in organic horizons of temperate forest soils. Soil Biol. Biochem. 2018, 126, 64–75. [Google Scholar] [CrossRef]
- Fatima, F.; Ahmad, M.M.; Verma, S.R.; Pathak, N. Relevance of phosphate solubilizing microbes in sustainable crop production: A review. Int. J. Environ. Sci. Technol. 2021, 19, 9283–9296. [Google Scholar] [CrossRef]
- Liu, Y.J.; Zhang, L.H.; Zhang, H.; Lan, B.; Lv, J.T.; Chen, G.; Wang, L.S.; Liu, Z.G. Role of phosphate solubilizing microorganisms in soil phosphorus cycle: A review. Microbiol. China 2023, 50, 3671–3687. [Google Scholar]
- Ai, P.H.; Sun, S.B.; Zhao, J.N.; Fan, X.R.; Xin, W.J.; Guo, Q.; Yu, L.; Shen, Q.R.; Wu, P.; Miller, A.J.; et al. Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation. Plant J. 2009, 57, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Wen, Y.; Ma, J.; Macdonald, A.; Hill, P.W.; Chadwick, D.R.; Wu, L.; Jones, D.L. Long-term farmyard manure application affects soil organic phosphorus cycling: A combined metagenomic and 33P/14C labelling study. Soil Biol. Biochem. 2020, 149, 107959. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, Y.; Fan, Y.F.; Zhang, L.; Li, X.Y.; Zhang, Q.Q.; Shu, Q.Y.; Huang, J.R.; Chen, G.Y.; Li, Q.; et al. Genetic improvement of phosphate-limited photosynthesis for high yield in rice. Proc. Natl. Acad. Sci. USA 2024, 121, e2404199121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.X.; Wang, H.R.; Ren, X.L.; Xiao, Y.H.; Liu, D.P.; Meng, W.J.; Qiu, Y.H.; Hu, B.; Xie, Q.J.; Chu, C.C.; et al. Brassinosteroid-dependent phosphorylation of PHOSPHATE STARVATION RESPONSE2 reduces its DNA-binding ability in rice. Plant Cell 2024, 36, 2253–2271. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Lai, H.L.; Gao, F.Y.; Zhang, R.P.; Wu, S.X.; Ge, F.R.; Li, Y.Y.; Yao, H.Y. The proliferation of beneficial bacteria influences the soil C, N, and P cycling in the soybean–maize intercropping system. Environ. Sci. Pollut. Res. 2024, 31, 25688–25705. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.L.; Zheng, Y.; Xiao, J.X. Below-ground biotic mechanisms of phosphorus uptake and utilization improved by cereal and legume intercropping-A review. Crops 2018, 4, 20–27. [Google Scholar]
- Zhu, S.G.; Cheng, Z.G.; Wang, J.; Gong, D.S.; Fazal, U.; Tao, H.Y.; Zhu, H.; Duan, H.X.; Yang, Y.M.; Xiong, Y.C. Soil phosphorus availability and utilization are mediated by plant facilitation via rhizosphere interactions in an intercropping system. Eur. J. Agron. 2023, 142 (Suppl. C), 126679. [Google Scholar] [CrossRef]
- Zhou, J.C.; Zhang, L.; Feng, G.; George, T.S. Arbuscular mycorrhizal fungi have a greater role than root hairs of maize for priming the rhizosphere microbial community and enhancing rhizosphere organic P mineralization. Soil Biol. Biochem. 2022, 171, 108713. [Google Scholar] [CrossRef]
- Zhou, Y.W.; Zhang, T.X.; Jin, S.Y.; Chen, S.; Zhang, Y.L. Effects of escherichia coli alkaline phosphatase PhoA on the mineralization of dissolved organic phosphorus. Water 2021, 13, 3315. [Google Scholar] [CrossRef]
- Hu, X.; Gu, H.; Liu, J.; Wei, D.; Zhu, P.; Cui, X.A.; Zhou, B.; Chen, X.; Jin, J.; Liu, X.; et al. Metagenomics reveals divergent functional profiles of soil carbon and nitrogen cycling under long-term addition of chemical and organic fertilizers in the black soil region. Geoderma 2022, 418, 115846. [Google Scholar] [CrossRef]
- Oliverio, A.M.; Bissett, A.; McGuire, K.; Saltonstall, K.; Turner, B.L.; Fierer, N. The role of phosphorus limitation in shaping soil bacterial communities and their metabolic capabilities. mBio 2020, 11, e01718–20. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Soil Texture | Organic Matter (g/kg) | Total Nitrogen (%) | Available Nitrogen (mg/kg) | Available Potassium (mg/kg) | Available Phosphorus (mg/kg) | Total Phosphorus (%) | Organic Phosphorus (mg/kg) | pH |
|---|---|---|---|---|---|---|---|---|---|
| Qihe | Fluvisols | 14.63 ± 0.45 | 0.10 ± 0.05 | 112.06 ± 0.35 | 142.12 ± 6.55 | 23.21 ± 1.02 | 0.097 ± 0.001 | 134.24 ± 9.91 | 7.91 ± 0.13 |
| Laizhou | Luvisols | 15.68 ± 0.24 | 0.07 ± 0.00 | 92.83 ± 0.43 | 114.64 ± 1.53 | 56.95 ± 1.04 | 0.044 ± 0.001 | 86.75 ± 1.93 | 5.44 ± 0.03 |
| Phylum | phnC | phnD | phnE | pstB | ugpC | pstA | ugpB | pstC | ugpE | ugpA | TP | ALP | AP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Thaumarchaeota | – | 0.85 | – | 0.8 | – | 0.81 | – | – | – | – | 0.81 | – | 0.84 |
| Acidobacteria | −0.81 | −0.85 | −0.84 | −0.89 | −0.82 | – | −0.82 | – | −0.85 | – | 0.88 | – | 0.88 |
| Actinobacteria | – | – | – | 0.86 | – | 0.82 | – | – | – | – | – | – | – |
| Armatimonadetes | −0.97 | −0.98 | −0.96 | −0.97 | −0.95 | −0.93 | −0.97 | −0.87 | −0.91 | −0.87 | 1 | 0.97 | 0.98 |
| Bacteroidetes | −0.97 | −0.98 | −0.96 | −0.98 | −0.94 | −0.93 | −0.96 | −0.87 | −0.91 | −0.87 | 1 | 0.96 | 0.96 |
| Candidatus Dormibacteraeota | 0.97 | 0.98 | 0.96 | 0.98 | 0.94 | 0.93 | 0.96 | 0.87 | 0.91 | 0.86 | 1 | 0.91 | 0.98 |
| Candidatus Eremiobacteraeota | −0.97 | −0.98 | −0.96 | −0.98 | −0.94 | −0.93 | −0.96 | −0.87 | −0.91 | −0.87 | 1 | 0.96 | 0.96 |
| Chloroflexi | 0.95 | 0.96 | 0.95 | 0.96 | 0.95 | 0.92 | 0.96 | 0.85 | 0.92 | 0.92 | 0.97 | 0.96 | 0.97 |
| Cyanobacteria | 0.84 | 0.8 | 0.88 | 0.81 | 0.87 | – | 0.84 | – | 0.86 | 0.88 | 0.81 | – | – |
| Deinococcus Thermus | −0.97 | −0.98 | −0.96 | −0.98 | −0.95 | −0.93 | −0.97 | −0.87 | −0.92 | −0.88 | 1 | 0.96 | 0.97 |
| Elusimicrobia | −0.81 | – | −0.85 | – | −0.85 | – | −0.81 | – | −0.85 | – | – | – | – |
| Gemmatimonadetes | −0.97 | −0.96 | −0.91 | −0.91 | −0.88 | −0.93 | −0.96 | −0.83 | −0.88 | 0.83 | 0.97 | 0.97 | 0.94 |
| Nitrospirae | 0.94 | 0.98 | 0.9 | 0.96 | 0.88 | 0.94 | 0.91 | 0.88 | 0.83 | – | 0.98 | 0.96 | 0.98 |
| Proteobacteria | 0.97 | 0.97 | 0.96 | 0.97 | 0.96 | 0.9 | 0.97 | 0.86 | 0.9 | 0.88 | 0.98 | 0.96 | 0.96 |
| phnC | phnD | phnE | pstB | ugpC | pstA | ugpB | ugpE | pstC | ugpA | |
|---|---|---|---|---|---|---|---|---|---|---|
| TP | 0.97 | 0.98 | 0.95 | 0.97 | 0.94 | 0.92 | 0.96 | 0.91 | 0.87 | 0.86 |
| AP | −0.94 | −0.97 | −0.93 | −0.97 | −0.93 | −0.9 | −0.93 | −0.89 | −0.83 | −0.85 |
| ALP | 0.85 | 0.96 | 0.9 | 0.92 | 0.89 | 0.88 | 0.92 | 0.84 | 0.81 | 0.8 |
| Treatment | Solirub robacter | Nocar dioides | Gaiella | Nitrospira | Aeromi crobium | Strep tomyces | Bradyr hizobium | Conexibacter | Sphing omonas | Other | Unclassified |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | 3.92 | 3.67 | 2.41 | 1.29 | 0.98 | 0.91 | 0.97 | 0.74 | 0.53 | 15.46 | 20.21 |
| T1 | 3.47 | 2.81 | 2.38 | 1.17 | 0.93 | 0.89 | 0.86 | 0.67 | 0.53 | 14.96 | 20.51 |
| T2 | 2.92 | 2.65 | 2.42 | 1.24 | 1.19 | 0.82 | 0.78 | 0.59 | 0.43 | 14.91 | 20.04 |
| Treatment | Not available | Gaiella | Bradyrhizobium | Pseudolabrys | Nocardioides | Streptomyces | Acidipila | Occallatibacter | Solirubrobacter | Ferru-ginibacter | Other | Unclassified |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | 42.14 | 4.80 | 2.35 | 2.11 | 3.16 | 1.22 | 0.76 | 0.83 | 0.91 | 0.64 | 22.72 | 18.35 |
| T1 | 39.74 | 3.91 | 2.36 | 2.31 | 0.97 | 1.4 | 1.79 | 1.66 | 0.84 | 0.89 | 25.78 | 18.32 |
| T2 | 45.04 | 3.92 | 2.51 | 2.2 | 1.14 | 1.34 | 0.97 | 0.98 | 0.84 | 0.55 | 22.59 | 17.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, C.; Kong, W.; Zhao, F.; Ju, F.; Liu, P.; Li, Z.; Liu, K.; Zhao, H. Enhancing Soil Phosphorus Availability in Intercropping Systems: Roles of Plant Growth Regulators. Agronomy 2025, 15, 1748. https://doi.org/10.3390/agronomy15071748
Gao C, Kong W, Zhao F, Ju F, Liu P, Li Z, Liu K, Zhao H. Enhancing Soil Phosphorus Availability in Intercropping Systems: Roles of Plant Growth Regulators. Agronomy. 2025; 15(7):1748. https://doi.org/10.3390/agronomy15071748
Chicago/Turabian StyleGao, Chunhua, Weilin Kong, Fengtao Zhao, Feiyan Ju, Ping Liu, Zongxin Li, Kaichang Liu, and Haijun Zhao. 2025. "Enhancing Soil Phosphorus Availability in Intercropping Systems: Roles of Plant Growth Regulators" Agronomy 15, no. 7: 1748. https://doi.org/10.3390/agronomy15071748
APA StyleGao, C., Kong, W., Zhao, F., Ju, F., Liu, P., Li, Z., Liu, K., & Zhao, H. (2025). Enhancing Soil Phosphorus Availability in Intercropping Systems: Roles of Plant Growth Regulators. Agronomy, 15(7), 1748. https://doi.org/10.3390/agronomy15071748