Water and Nitrogen Dynamics of Mungbean as a Summer Crop in Temperate Environments

 , , , , and
, , , , and
Abstract
1. Introduction
- (i)
- Increasing water supply to mungbean during the early podding period will be more beneficial to yield than increasing water supply during the vegetative growth phase.
- (ii)
- Nitrogen fixation will be strongly affected by water availability at key growth stages of mungbean grown in southern Australia.
2. Materials and Methods
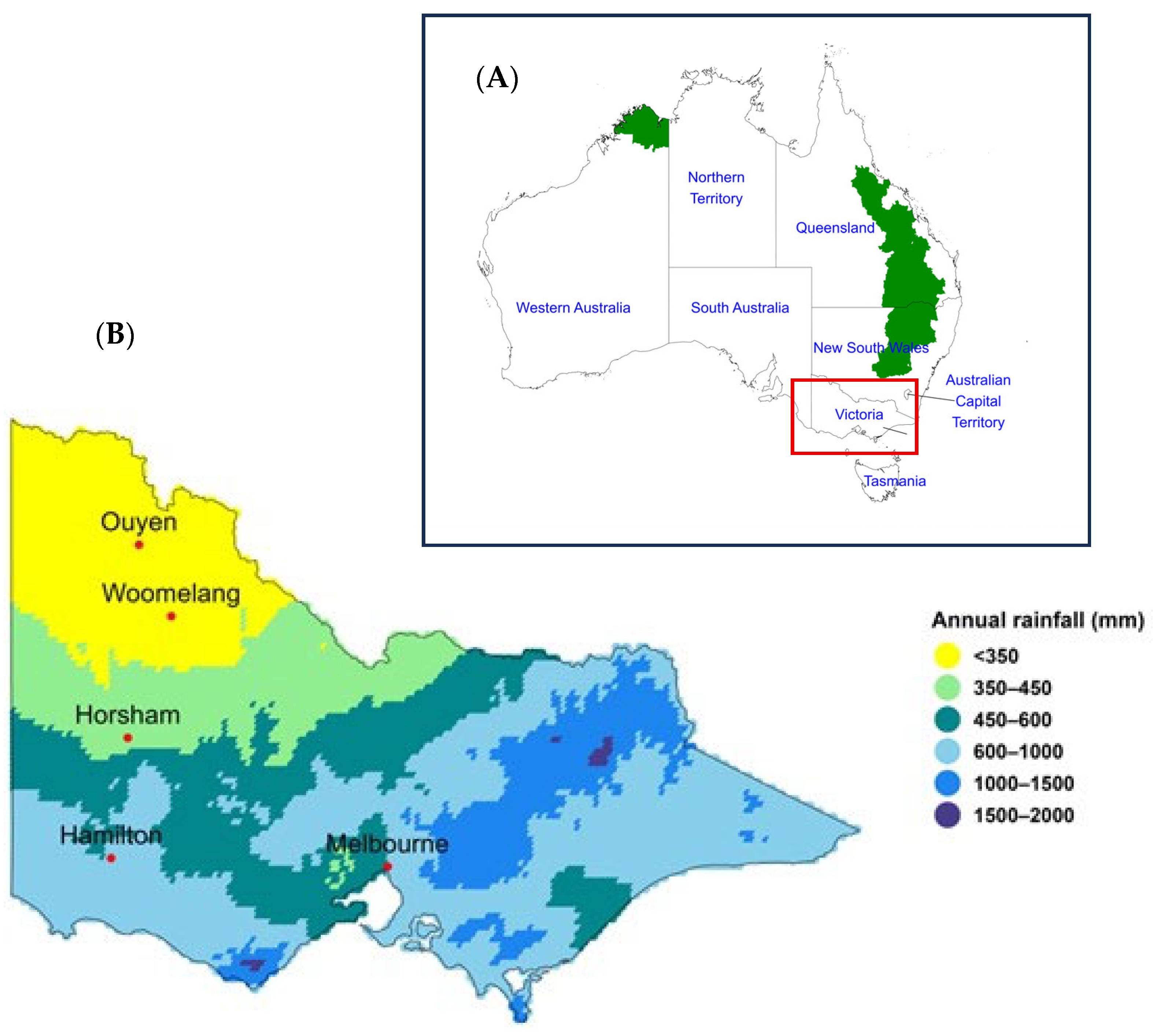
2.1. Site Description
2.2. Experimental Design
2.3. Crop Management
2.4. Measurement of Nitrogen Fixation
2.5. Data Collection
2.5.1. Soil Water
2.5.2. Phenology
2.5.3. Crop and Yield Components
2.6. Data Analysis
3. Results
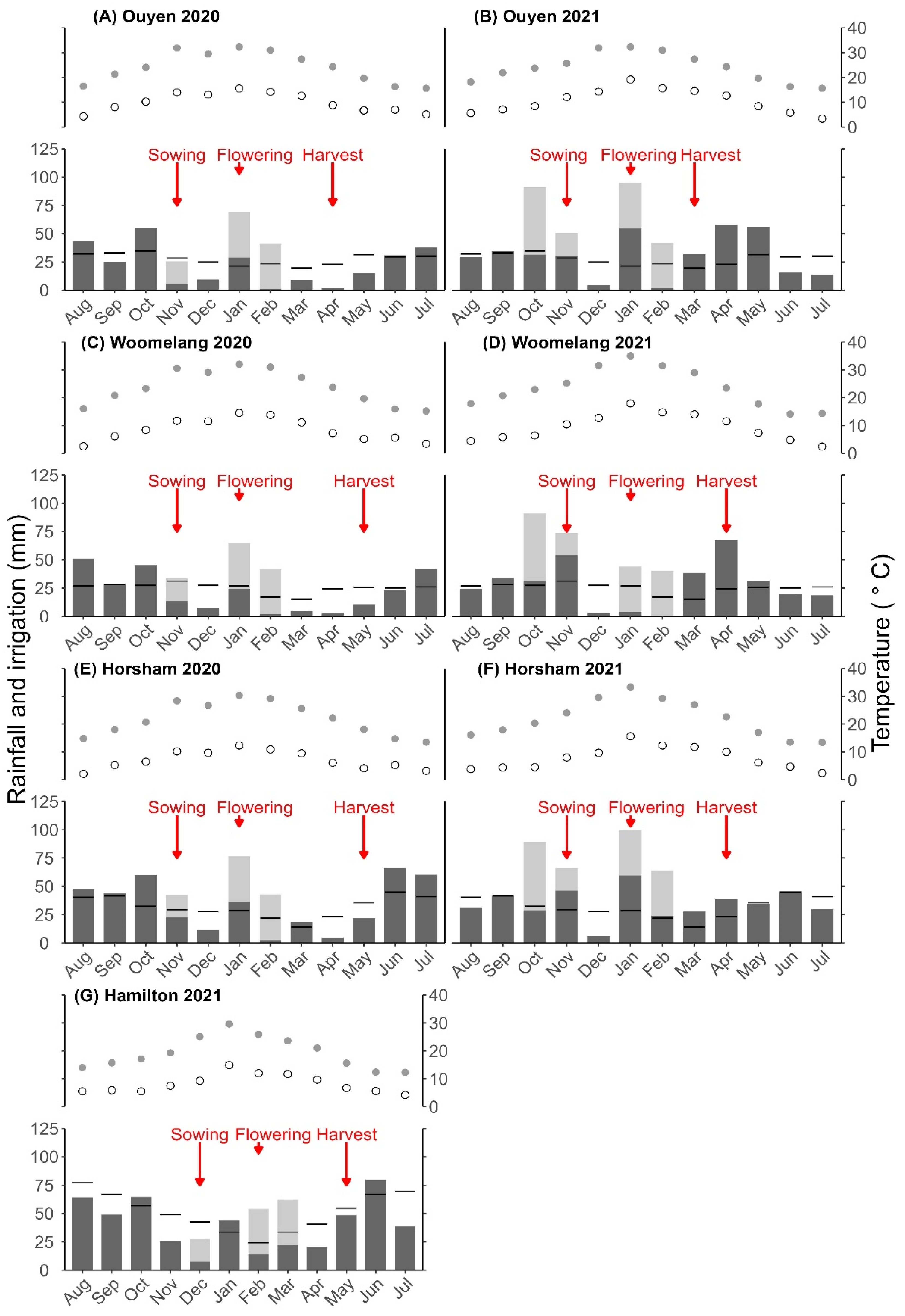
3.1. Weather
3.2. Days to Flowering
3.3. Biomass and Yield Components
3.3.1. Hamilton 2021
3.3.2. Horsham 2020
3.3.3. Horsham 2021
3.3.4. Ouyen 2020
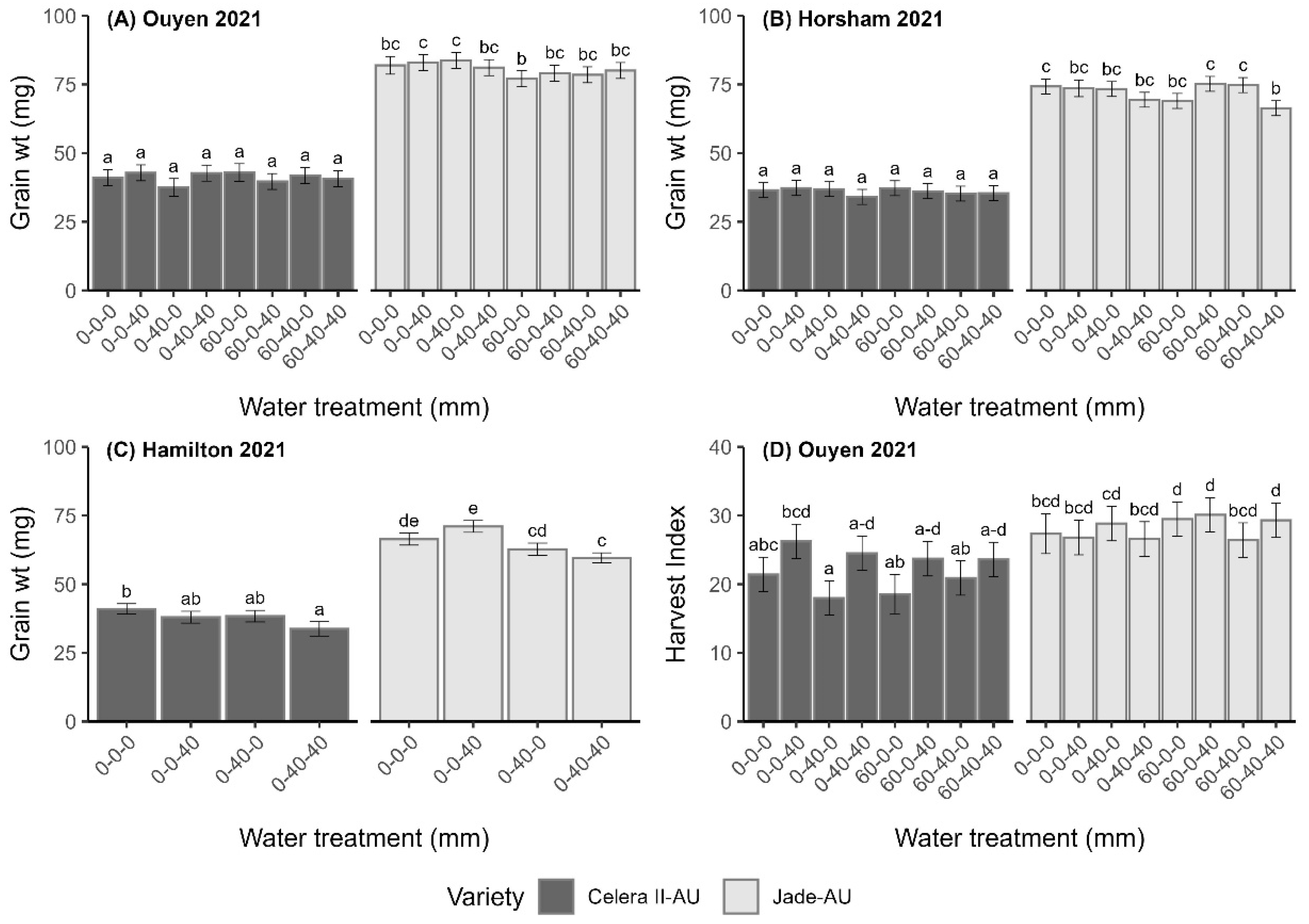
3.3.5. Ouyen 2021
3.3.6. Woomelang 2020
3.3.7. Woomelang 2021
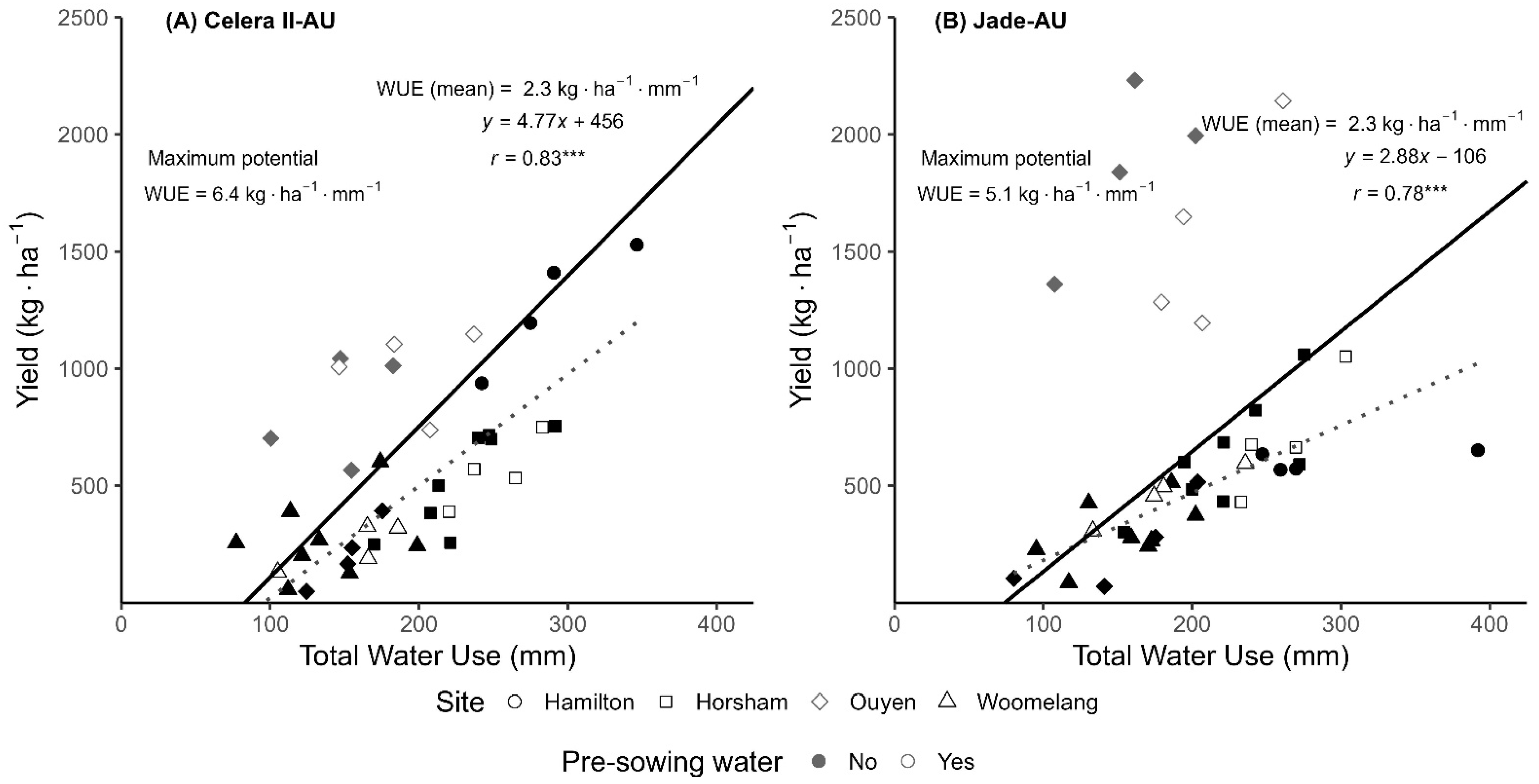
3.4. Water Use and Water Use Efficiency (WUE)
3.5. Nitrogen Derived from the Atmosphere (%Ndfa)
3.6. Total N Fixation
4. Discussion
4.1. Relationship of Weather, Water Availability and Phenology to Mungbean Yield
4.2. The Relationship Between Water Availability and Yield-Determining Attributes
4.3. Relationship Between Yield and Water Use
4.4. Effect of Water Availability at Different Growth Stages on %Ndfa and Total N Fixation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, Y.-S.; Sun, S.; FitzGerald, R. Mung bean proteins and peptides: Nutritional, functional and bioactive properties. Food Nutr. Res. 2018, 62, 1290. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Schafleitner, R.; Kenyon, L.; Srinivasan, R.; Easdown, W.; Ebert, A.; Hanson, P. Genetic improvement of mungbean. SABRAO J. Breed. Genet. 2012, 44, 177–190. [Google Scholar]
- Nair, R.; Schreinemachers, P. Global status and economic importance of mungbean. In The Mungbean Genome; Nair, R.M., Schafleitner, R., Lee, S.-H., Kole, C., Eds.; Compendium of Plant Genomes; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–8. [Google Scholar]
- Australian Mungbean Association. Industry Size and Value. Available online: http://www.mungbean.org.au/about-us.html (accessed on 25 March 2021).
- Nair, R.M.; Alam, A.M.; Douglas, C.; Gowda, A.; Pratap, A.; Win, M.M.; Karimi, R.; Emmanuel, M.K.; Binagwa, P.; Boddepalli, V.; et al. Establishing the International Mungbean Improvement Network; Australian Centre for International Agricultural Research: Canberra, Australia, 2022; p. 118.
- Tarahi, M. The potential application of mung bean (Vigna radiata L.) protein in plant- based food analogs: A review. Legume Sci. 2024, 6, e70011. [Google Scholar] [CrossRef]
- Diego, R. Multidisciplinary Research: A Key to Unlocking Effective Innovation for Pulse Crops. Available online: https://iyp2016.org/news/320-multidisciplinary-research-a-key-to-unlocking-effective-innovation-for-pulse-crops (accessed on 11 April 2024).
- Research and Markets. Mung Beans Market. Available online: https://www.researchandmarkets.com/report/mung-bean?srsltid=AfmBOopkR7J8AKnj2bPOPr_d8o66eVjRjbes1vvvCTgxICjv21Ke8yXW (accessed on 27 June 2025).
- Wells, L. Rain, Firming Prices Buoy Outlook for CQ Mungbeans. Available online: https://www.graincentral.com/cropping/pulses/rain-firming-prices-buoy-outlook-for-cq-mungbeans/ (accessed on 30 June 2023).
- Wells, L. Pulse update: Prices steady, demand strong. Grain Cent. 2021. [Google Scholar]
- McCarrol, A.P. Exporting Food Quality Mungbeans to Taiwan: The Importance of Supply Chain Entities; Seoul National University: Seoul, Republic of Korea, 2024. [Google Scholar]
- AEGIC. Australian Pulses. Available online: https://www.aegic.org.au/wp-content/uploads/2024/11/AEGIC-Australian-pulses-brochure-2024-LR.pdf (accessed on 4 June 2025).
- Delahunty, A.; Wallace, A.; Norton, S.; Henry, F.; Riffkin, P.; Christy, B.; Brand, J.; Clancy, A.; Silwal, S.; Partington, D.; et al. New legume species as opportunistic summer crops for southern Australia–Part 2 Exploring global germplasm for increasing crop adaptation. In Proceedings of the 20th Australian Agronomy Conference, Toowoomba, QLD, Australia, 18–22 September 2022. [Google Scholar]
- Harris, R.H.; Armstrong, R.D.; Wallace, A.J.; Belyaeva, O.N. Delaying nitrogen fertiliser application improves wheat 15N recovery from high rainfall cropping soils in south eastern Australia. Nutr. Cycl. Agroecosyst. 2016, 106, 113–128. [Google Scholar] [CrossRef]
- Bureau of Meteorology. Rainfall Variability Maps. Available online: http://www.bom.gov.au/climate/maps/averages/rainfall-variability/?period=dec (accessed on 18 April 2025).
- Shanice, V.H.; Dudley, C.; Kang, Y.; Smith, D.; Nair, R.M.; Douglas, C.A.; Potgieter, A.; Robinson, H.; Hickey, L.T.; Smith, M.R. Building a better Mungbean: Breeding for reproductive resilience in a changing climate. Food Energy Secur. 2023, 12, e467. [Google Scholar] [CrossRef]
- Grains Research and Development Corporation. Mungbeans. Grownotes 2017, 282. [Google Scholar]
- Gentry, J. Mungbean Management Guide; The state of Queensland, Department of Employment, Economic Development and Innovation: Brisbane, Australia, 2010.
- Sadasivan, R.; Natrajaratnam, N.; Dabu, R.; Muralidharan, V.; Rangasmay, S. Response of mungbean cultivars to soil moisture stress at different growth phases. In Proceedings of the Mungbean, second international symposium, Bangkok, Thailand, 16–20 November 1987; pp. 260–262. [Google Scholar]
- Nielsen, D.C.; Nelson, N.O. Black bean sensitivity to water stress at various growth stages. Crop Sci. 1998, 38, 422–427. [Google Scholar] [CrossRef]
- Pannu, R.K.; Singh, D.P. Effect of irrigation on water use, water-use efficiency, growth and yield of mungbean. Field Crops Res. 1993, 31, 87–100. [Google Scholar] [CrossRef]
- Thomas; Robertson, M.J.; Fukai, S.; Peoples, M.B. The effect of timing and severity of water deficit on growth, development, yield accumulation and nitrogen fixation of mungbean. Field Crops Res. 2004, 86, 67–80. [Google Scholar] [CrossRef]
- El-Nakhlawy, F.S.; Ismail, S.M.; Basahi, J.M. Optimizing mungbean productivity and irrigation water use efficiency through the use of low water-consumption during plant growth stages. Legume Res. 2018, 41, 108–113. [Google Scholar] [CrossRef]
- Wenham, K.; Williams, A.; Rachaputi, R.; Rossignol, T.; Collins, M. Critical Period of Moisture Vulnerability in Mungbeans. GRDC Grains Research Update. 2020. Available online: https://grdc.com.au/__data/assets/pdf_file/0030/430887/GRDC-Update-Paper-Wenham-Kylie-July-2020.pdf (accessed on 9 June 2025).
- De Costa, W.A.J.M.; Shanmugathasan, K.N.; Joseph, K.D.S.M. Physiology of yield determination of mung bean (Vigna radiata (L.) Wilczek) under various irrigation regimes in the dry and intermediate zones of Sri Lanka. Field Crops Res. 1999, 61, 1–12. [Google Scholar] [CrossRef]
- Bangar, P.; Chaudhury, A.; Tiwari, B.; Kumar, S.; Kumari, R.; Bhat, K.V. Morphophysiological and biochemical response of mungbean [Vigna radiata (L.) Wilczek] varieties at different developmental stages under drought stress. Turk. J. Biol. 2019, 43, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Geetika, G.; Collins, M.; Singh, V.; Hammer, G.; Mellor, V.; Smith, M.; Rachaputi, R.C.N. Canopy and reproductive development in mungbean (Vigna radiata). Crop Pasture Sci. 2022, 73, 1142–1155. [Google Scholar] [CrossRef]
- Rachaputi, R.C.N.; Sands, D.; McKenzie, K.; Agius, P.; Lehane, J.; Seyoum, S. Eco-physiological drivers influencing mungbean [Vigna radiata (L.) Wilczek] productivity in subtropical Australia. Field Crops Res. 2019, 238, 74–81. [Google Scholar] [CrossRef]
- Lamichaney, A.; Parihar, A.K.; Hazra, K.K.; Dixit, G.P.; Katiyar, P.K.; Singh, D.; Singh, A.K.; Kumar, N.; Singh, N.P. Untangling the influence of heat stress on crop phenology, seed set, seed weight, and germination in field pea (Pisum sativum L.). Front. Plant Sci. 2021, 12, 635868. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Sarker, B.C.; Alam, M.A.; Javed, T.; Alam, M.J.; Zaman, M.S.U.; Azam, M.G.; Shabbir, R.; Raza, A.; Habib-ur-Rahman, M.; et al. Yield stability and genotype environment interaction of water deficit stress tolerant mung bean (Vigna radiata l. Wilczak) genotypes of bangladesh. Agronomy 2021, 11, 2136. [Google Scholar] [CrossRef]
- Parihar, A.K.; Gupta, S.; Hazra, K.K.; Lamichaney, A.; Sen Gupta, D.; Singh, D.; Kumar, R.; Singh, A.K.; Vaishnavi, R.; Jaberson, M.S.; et al. Multi-location evaluation of mungbean (Vigna radiata L.) in Indian climates: Ecophenological dynamics, yield relation, and characterization of locations. Front. Plant Sci. 2022, 13, 984912. [Google Scholar] [CrossRef] [PubMed]
- Unkovich, M.J.; Baldock, J.; Peoples, M.B. Prospects and problems of simple linear models for estimating symbiotic N2 fixation by crop and pasture legumes. Plant Soil 2010, 329, 75–89. [Google Scholar] [CrossRef]
- Gentry, J.; Sands, D.J.; Silburn, C.; Dunn, M.; Seymour, N.P.; Bell, K.L. Mungbeans: Response to applied nitrogen. In Proceedings of the 20th Agronomy Australia Conference, Toowoomba, Queensland, 18–22 September 2022. [Google Scholar]
- Martin, R.; Montgomery, S.; Phan, S.; Im, S. Mungbean Production Guide for Cambodian Conditions; ACIAR Monograph No. 162; ACIAR: Bruce, ACT, Australia, 2015; pp. 1–52. [Google Scholar]
- McIntosh, P. Protecting Mungbean Inoculant. Available online: http://pulseaus.com.au/blog/post/protecting-mungbean-inoculant (accessed on 28 March 2021).
- Hauser, S.; Grams, C.M.; Reeder, M.J.; McGregor, S.; Fink, A.H.; Quinting, J.F. A weather system perspective on winter–spring rainfall variability in southeastern Australia during El Niño. Q. J. R. Meteorol. Soc. 2020, 146, 2614–2633. [Google Scholar] [CrossRef]
- Bureau of Meteorology. Climate Data Online. Available online: https://reg.bom.gov.au/climate/data/index.shtml (accessed on 13 November 2023).
- Australian Mungbean Association. Australian Mungbean Association Strategic Plan 2020–2025; Australian Mungbean Association: Dalby, QLD, Australia, 2021; pp. 1–26. [Google Scholar]
- Bureau of Meteorology. Average Annual, Seasonal and Monthly Rainfall. Available online: http://www.bom.gov.au/jsp/ncc/climate_averages/rainfall/index.jsp?period=an&area=vc#maps (accessed on 27 March 2021).
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984; p. 680. [Google Scholar]
- NSW DPI. Summer Cropping Options for Northern and Central NSW. Available online: https://www.dpi.nsw.gov.au/__data/assets/pdf_file/0003/1415388/summer-crop-options-2022.pdf (accessed on 19 March 2025).
- Rachaputi, R.C.N.; Sands, D.; McKenzie, K.; Lehane, J.; Agius, P.; Seyoum, S.; Peak, A. Water extraction patterns of mungbean (Vigna radiata) in diverse subtropical environments. Agric. Water Manag. 2019, 219, 109–116. [Google Scholar] [CrossRef]
- Unkovich, M.; Herridge, D.; Peoples, M.; Cadisch, G.; Boddey, B.; Giller, K.; Alves, B.; Chalk, P. Measuring Plant-Associated Nitrogen Fixation in Agricultural Systems; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 2008.
- Diatta, A.A.; Thomason, W.E.; Abaye, O.; Thompson, T.L.; Battaglia, M.L.; Vaughan, L.J.; Lo, M.; Filho, J.F.D.C.L. Assessment of nitrogen fixation by mungbean genotypes in different soil textures using 15N natural abundance method. J. Soil Sci. Plant Nutr. 2020, 20, 2230–2240. [Google Scholar] [CrossRef]
- Gan, Y.; Hamel, C.; O’Donovan, J.T.; Cutforth, H.; Zentner, R.P.; Campbell, C.A.; Niu, Y.; Poppy, L. Diversifying crop rotations with pulses enhances system productivity. Sci. Rep. 2015, 5, 14625. [Google Scholar] [CrossRef] [PubMed]
- The VSNi Team. Asreml: Fits Linear Mixed Models Using REML, R package version 4.2.0.355; VSN International: Hemel Hempstead, UK, 2023.
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Rogers, S.; Conway, A. Biometryassist: Functions to Assist Design and Analysis of Agronomic Experiments, R package version 1.3.0; The University of Adelaide: Adelaide, Australia, 2025.
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests, R package version 0.7.2. 2023. Available online: https://rpkgs.datanovia.com/rstatix/authors.html#citation (accessed on 10 June 2025).
- Chauhan, Y.; Williams, R. Physiological and agronomic strategies to increase mungbean yield in climatically variable environments of northern Australia. Agronomy 2018, 8, 83. [Google Scholar] [CrossRef]
- French, R.; Schultz, J. Water use efficiency of wheat in a Mediterranean-type environment. I. The relation between yield, water use and climate. Aust. J. Agric. Res. 1984, 35, 743–764. [Google Scholar] [CrossRef]
- Bashandi, M.M.H.; Poehlman, J.M. Photoperiod response in mungbeans (Vigna radiata (L.) Wilczek). Euphytica 1974, 23, 691–697. [Google Scholar] [CrossRef]
- Imrie, B.C.; Lawn, R.J. Time to flowering of mung bean (Vigna radiata) genotypes and their hybrids in response to photoperiod and temperature. Exp. Agric. 1990, 26, 307–318. [Google Scholar] [CrossRef]
- Lawn, R. Agronomic studies on Vigna spp. in south-eastern Queensland. I. Phenological response of cultivars to sowing date. Aust. J. Agric. Res. 1979, 30, 855–870. [Google Scholar] [CrossRef]
- Ellis, R.H.; Lawn, R.J.; Summerfield, R.J.; Qi, A.; Roberts, E.H.; Chay, P.M.; Brouwer, J.B.; Rose, J.L.; Yeates, S.J.; Sandover, S. Towards the reliable prediction of time to flowering in six annual crops. IV. cultivated and wild mung bean. Exp. Agric. 1994, 30, 31–43. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, K.D. Physiological responses and dry matter partitioning of summer mungbean (Vigna radiata L.) genotypes subjected to drought conditions. J. Agron. Crop Sci. 2009, 195, 270–277. [Google Scholar] [CrossRef]
- Kaur, R.; Bains, T.S.; Bindumadhava, H.; Nayyar, H. Responses of mungbean (Vigna radiata L.) genotypes to heat stress: Effects on reproductive biology, leaf function and yield traits. Sci. Hortic. 2015, 197, 527–541. [Google Scholar] [CrossRef]
- Jha, U.C.; Shafi, S.; Tallury, S.; Nayyar, H.; Ciampitti, I.A.; Siddique, K.H.M.; Prasad, P.V.V. Differential physiological and yield responses of selected mung bean (Vigna radiata (L.) R. Wilczek) genotypes to various high-temperature stress regimes. Sci. Rep. 2025, 15, 1034. [Google Scholar] [CrossRef] [PubMed]
- Geetika, G.; Hammer, G.; Smith, M.; Singh, V.; Collins, M.; Mellor, V.; Wenham, K.; Rachaputi, R.C. Quantifying physiological determinants of potential yield in mungbean (Vigna radiata (L.) Wilczek). Field Crops Res. 2022, 287, 108648. [Google Scholar] [CrossRef]
- Patriyawaty, N.R.; Rachaputi, R.C.N.; George, D. Physiological mechanisms underpinning tolerance to high temperature stress during reproductive phase in mungbean (Vigna radiata (L.) Wilczek). Environ. Exp. Bot. 2018, 150, 188–197. [Google Scholar] [CrossRef]
- Iqbal, J.; Ahsan, M.; Saleem, M.; Ali, A. Appraisal of gene action for indeterminate growth in mungbean [Vigna radiata (L.) Wilczek]. Front. Plant Sci. 2015, 6, 665. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.; Kwon, H.; Cho, K.-H.; Yoon, M.Y.; Kim, M.Y.; Lee, S.-H. Identification of epigenetic variation associated with synchronous pod maturity in mungbean (Vigna radiata L.). Sci. Rep. 2020, 10, 17414. [Google Scholar] [CrossRef] [PubMed]
- Marwiyah, S.; Suwarno, W.B.; Wirnas, D.; Trikoesoemaningtyas; Sutjahjo, S.H. Genotype by environment interaction on phenology and synchronous maturity of mungbean. Agron. J. 2021, 113, 2321–2334. [Google Scholar] [CrossRef]
- Christy, B.; Delahunty, A.; Norton, S.; Wallace, A.; Riffkin, P.; O’Leary, G.J.; Nuttall, J. New legume species as opportunistic summer crops for southern Australia–Part 1: Environmental suitability. In Proceedings of the 20th Australian Agronomy Conference, Toowoomba, QLD, Australia, 18–22 September 2022. [Google Scholar]
- Gölgül, İ.; Kırnak, H.; Ali İrik, H. Yield components and crop water stress index (CWSI) of mung bean grown under deficit irrigations. Gesunde Pflanz. 2023, 75, 271–281. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Squire, G.R. The Physiology of Tropical Crop Production; C. A. B. International: Wallingford, UK, 1990; p. 236. [Google Scholar]
- Singh, D.P.; Singh, P.; Sharma, H.C.; Turner, N.C. Influence of water deficits on the water relations, canopy gas exchange, and yield of chickpea (Cicer arietinum). Field Crops Res. 1987, 16, 231–241. [Google Scholar] [CrossRef]
- Jamieson, P.D.; Martin, R.J.; Francis, G.S.; Wilson, D.R. Drought effects on biomass production and radiation-use efficiency in barley. Field Crops Res. 1995, 43, 77–86. [Google Scholar] [CrossRef]
- Goel, K.; Kundu, P.; Sharma, P.; Zinta, G. Thermosensitivity of pollen: A molecular perspective. Plant Cell Rep. 2023, 42, 843–857. [Google Scholar] [CrossRef] [PubMed]
- Iovane, M.; Aronne, G. High temperatures during microsporogenesis fatally shorten pollen lifespan. Plant Reprod 2022, 35, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Y.; Nakayama, N.; Saneoka, H.; Fujita, K. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol. Plant. 2006, 50, 138–141. [Google Scholar] [CrossRef]
- Zeleke, K.; Luckett, D.J. Irrigated agricultural production dynamics in response to rainfall variability and water policy reforms in the southern Murray-Darling Basin of Australia. Agric. Water Manag. 2025, 315, 109539. [Google Scholar] [CrossRef]
- Goulburn-Murray Water. Goulburn-Murray Water at a Glance. Available online: https://www.g-mwater.com.au/about/waterclass/resources/videos/the-gmid.html (accessed on 29 June 2025).
- Delahunty, A.; Henry, F.; Wallace, A.; Christy, B.; Brand, J.; Norton, S.; Perry, E.; Riffkin, P.; Walker, C.; O’Leary, G.; et al. Alternative Legume Crop Options for Southern Australia—A Review; Agriculture Victoria, Department of Jobs, Precincts and Regions (DJPR): Melbourne, VIC, USA, 2020; pp. 1–152.
- Matthews, R.B.; Harris, D.; Williams, J.H.; Rao, R.C.N. The physiological basis for yield differences between four genotypes of groundnut (Arachis hypogaea) in response to drought. II. Solar radiation interception and leaf movement. Exp. Agric. 1988, 24, 203–213. [Google Scholar] [CrossRef]
- Silwal, S.; Bhattarai, S.P.; Midmore, D.J. Aerobic rice with or without strategic irrigation in the subtropics. Agronomy 2020, 10, 1831. [Google Scholar] [CrossRef]
- Collins, M.; Anderson, B.; Bell, L. Yield Gaps in Mungbean Crops Across the Northern Grains Region. GRDC Grains Research Update 2020, pp. 100–106. Available online: https://grdc.com.au/__data/assets/pdf_file/0023/433823/GRDC-Grains-Research-Update-Toowoomba-2020.pdf (accessed on 31 July 2022).
- Bourgault, M.; Madramootoo, C.A.; Webber, H.A.; Stulina, G.; Horst, M.G.; Smith, D.L. Effects of deficit irrigation and salinity stress on common bean (Phaseolus vulgaris L.) and mungbean (Vigna radiata (L.) wilczek) grown in a controlled environment. J. Agron. Crop Sci. 2010, 196, 262–272. [Google Scholar] [CrossRef]
- Lu, S.; Liu, M.; Yi, J.; Li, S.; Xu, Y.; Zhang, H.; Ding, F. Lateral partition patterns and controlling factors of soil infiltration at a steep, near-stream, and humid hillslope scale. Catena 2024, 239, 107917. [Google Scholar] [CrossRef]
- Peoples, M.B.; Brockwell, J.; Herridge, D.F.; Rochester, I.J.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M.; Dakora, F.D.; Bhattarai, S.; Maskey, S.L.; et al. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Shah, Z.; Shah, S.H.; Peoples, M.B.; Schwenke, G.D.; Herridge, D.F. Crop residue and fertiliser N effects on nitrogen fixation and yields of legume–cereal rotations and soil organic fertility. Field Crops Res. 2003, 83, 1–11. [Google Scholar] [CrossRef]
- Djekoun, A.; Planchon, C. Water status effect on dinitrogen fixation and photosynthesis in soybean. Agron. J. 1991, 83, 316–322. [Google Scholar] [CrossRef]
- Raji, S.G.; Tzanakakis, V.; Dörsch, P. Bradyrhizobial inoculation and P application effects on haricot and mung beans in the Ethiopian Rift Valley. Plant Soil 2019, 442, 271–284. [Google Scholar] [CrossRef]
- Herridge, D.F.; Robertson, M.J.; Cocks, B.; Peoples, M.B.; Holland, J.F.; Heuke, L. Low nodulation and nitrogen fixation of mungbean reduce biomass and grain yields. Aust. J. Exp. Agric. 2005, 45, 269–277. [Google Scholar] [CrossRef]
- Diatta, A.A.; Thomason, W.E.; Abaye, O.; Vaughan, L.J.; Thompson, T.L.; Lo, M.; Chim, B.K.; Bateman, S. Inoculation and soil texture effects on yield and yield components of mungbean. J. Agric. Sci. 2018, 10, 6. [Google Scholar] [CrossRef]
- Mott, J.; Abaye, O.; Reiter, M.; Maguire, R. Evaluating effects of Bradyrhizobium and arbuscular mycorrhizal fungi inoculation on yield components of mung bean (Vigna radiata (L.) wilczek) and nitrogen fixation. Agronomy 2022, 12, 2358. [Google Scholar] [CrossRef]
- Christopher, M.; Macdonald, B.; Yeates, S.; Ziegler, D.; Seymour, N. Wild bradyrhizobia that occur in the Burdekin region of Queensland are as effective as commercial inoculum for mungbean (Vigna radiata (L.)) and black gram (Vigna mungo (L.)) in fixing nitrogen and dry matter production. Appl. Soil Ecol. 2018, 124, 88–94. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Location | |||
|---|---|---|---|---|
| Hamilton | Horsham | Ouyen | Woomelang | |
| Soil type (Australian Soil Classification) | Chromosol | Vertosol | Calcarosol | Calcarosol |
| Soil pH (CaCl2), 2020 | – | 8.2 | 8.3 | 8.4 |
| Soil pH (CaCl2), 2021 | 5.8 | 8.1 | 8.3 | 8.0 |
| Soil nitrate (mg·kg−1), 2020 | – | 7.8 | 5.0 | 5.2 |
| Soil nitrate (mg·kg−1), 2021 | 6.0 | 4.12 | 8.3 | 8.0 |
| GSR (mm), (2020) | – | 93 | 52 | 24 |
| GSR (mm), (2021) | 133 | 127 | 108 | 137 |
| Sowing (2020) | – | 5 November 2020 | 3 November 2020 | 4 November 2020 |
| Sowing (2021) | 23 December 2021 | 19 November 2021 | 4 November 2021 | 12 November 2021 |
| Harvest (2020) | – | 11 May 2021 | 21 April 2021 | 19 May 2021 |
| Harvest (2021) | 4 May 2022 | 28 April 2022 | 31 March 2022 | 4 May 2022 |
| Treatment | Growth Stages and Water Application | |||
|---|---|---|---|---|
| Pre-Sowing (mm) | Sowing (mm) | Vegetative (mm) | Reproductive (mm) | |
| 0-0-0 | 0 | 20 | 0 | 0 |
| 0-0-40 | 0 | 20 | 0 | 40 |
| 0-40-0 | 0 | 20 | 40 | 0 |
| 0-40-40 | 0 | 20 | 40 | 40 |
| 60-0-0 | 60 | 20 | 0 | 0 |
| 60-0-40 | 60 | 20 | 0 | 40 |
| 60-40-0 | 60 | 20 | 40 | 0 |
| 60-40-40 | 60 | 20 | 40 | 40 |
| Site | Year | Days to Flowering | Sowing to Flowering GDDs | One Week to Flowering Average Photoperiod (h) |
|---|---|---|---|---|
| Hamilton | 2021 | 38 | 495 | 14.15 |
| Horsham | 2020 | 69 | 829 | 14.50 |
| Horsham | 2021 | 67 | 913 | 14.22 |
| Ouyen | 2020 | 78 | 1170 | 14.18 |
| Ouyen | 2021 | 68 | 974 | 14.37 |
| Woomelang | 2020 | 71 | 985 | 14.38 |
| Woomelang | 2021 | 78 | 1195 | 14.03 |
| Treatment | Podding Biomass (kg·ha−1) | Yield Component | ||||
|---|---|---|---|---|---|---|
| Grain Number (grains·m−2) | Grain Weight (mg) | Yield (kg·ha−1) | HI (%) | |||
| Water | 0-0-0 | 1564 | 1599 | 54 bc | 755 | 19 |
| 0-40-0 | 2564 | 2505 | 51 b | 989 | 16 | |
| 0-0-40 | 1243 | 2048 | 54 c | 915 | 21 | |
| 0-40-40 | 2117 | 1818 | 47 a | 1079 | 17 | |
| Variety | Celera II-AU | 1698 | 3224 b | 38 a | 1262 b | 26 b |
| Jade-AU | 2047 | 761 a | 65 b | 607 a | 11 a | |
| Variety | p-value | <0.001 | <0.001 | <0.001 | <0.001 | |
| Year | Treatment | Podding Biomass (kg·ha−1) | Yield Component | ||||
|---|---|---|---|---|---|---|---|
| Grain Number (grains·m−2) | Grain Weight (mg) | Yield (kg·ha−1) | HI (%) | ||||
| 2020 | Water | 0-0-0 | 1460 a | 497 a | 59 a | 276 a | 26 a |
| 0-40-0 | 2104 bc | 749 b | 59 a | 387 ab | 26 a | ||
| 0-0-40 | 1809 ab | 800 b | 65 b | 502 b | 30 b | ||
| 0-40-40 | 2455 c | 1299 c | 64 b | 769 c | 31 b | ||
| Variety | Celera II-AU | 1894 | 996 a | 44 a | 438 | 27 a | |
| Jade-AU | 2019 | 677 b | 80 b | 529 | 30 b | ||
| Water | p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Variety | p-value | <0.001 | <0.001 | <0.001 | |||
| 2021 | Water | 0-0-0 | 984 | 655 a | 55 b | 345 a | 26 ab |
| 0-40-0 | 1407 | 1442 bc | 55 b | 695 ab | 29 ab | ||
| 0-0-40 | 1073 | 1327 abc | 55 b | 645 ab | 27 ab | ||
| 0-40-40 | 1308 | 1871 c | 52 ab | 907 b | 30 b | ||
| 60-0-0 | 1011 | 836 ab | 53 ab | 410 a | 25 a | ||
| 60-40-0 | 1461 | 1194 abc | 54 ab | 598 ab | 27 ab | ||
| 60-0-40 | 958 | 1248 abc | 56 b | 623 ab | 27 ab | ||
| 60-40-40 | 1672 | 1611 c | 51 a | 811 b | 29 ab | ||
| Variety | Celera II-AU | 1067 a | 1568 a | 36 a | 582 | 29 a | |
| Jade-AU | 1401 b | 978 b | 72 b | 676 | 27 b | ||
| Water | p-value | <0.001 | 0.003 | <0.001 | 0.027 | ||
| Variety | p-value | 0.014 | <0.001 | <0.001 | 0.004 | ||
| Year | Treatment | Podding Biomass (kg·ha−1) | Yield Component | ||||
|---|---|---|---|---|---|---|---|
| Grain Number (grains·m−2) | Grain Weight (mg) | Yield (kg·ha−1) | HI (%) | ||||
| 2020 | Water | 0-0-0 | 383 a | 135 a | 53 ab | 77 a | 18 a |
| 0-40-0 | 815 bc | 272 ab | 49 a | 118 ab | 20 a | ||
| 0-0-40 | 519 ab | 459 b | 61 b | 258 b | 26 ab | ||
| 0-40-40 | 962 c | 846 c | 56 ab | 455 c | 29 b | ||
| Variety | Celera II-AU | 588 | 529 b | 40 a | 211 | 25 | |
| Jade-AU | 752 | 327 a | 70 b | 243 | 22 | ||
| Water | p-value | 0.001 | <0.001 | 0.006 | <0.001 | 0.006 | |
| Variety | p-value | 0.005 | <0.001 | ||||
| 2021 | Water | 0-0-0 | 1331 a | 1671 ab | 62 | 1032 ab | 24 |
| 0-40-0 | 1442 ab | 1989 ab | 61 | 1399 ab | 23 | ||
| 0-0-40 | 1452 ab | 2344 b | 63 | 1441 ab | 27 | ||
| 0-40-40 | 1511 ab | 2420 b | 62 | 1504 b | 26 | ||
| 60-0-0 | 1597 ab | 1366 a | 60 | 839 a | 24 | ||
| 60-40-0 | 2148 b | 1644 ab | 60 | 967 ab | 24 | ||
| 60-0-40 | 1586 ab | 2061 ab | 59 | 1376 ab | 27 | ||
| 60-40-40 | 2035 ab | 2379 b | 60 | 1412 ab | 26 | ||
| Variety | Celera II-AU | 1472 a | 1927 | 41 a | 839 a | 22 a | |
| Jade-AU | 1804 b | 2042 | 81 b | 1654 b | 28 b | ||
| Water | p-value | 0.02 | 0.004 | 0.004 | |||
| Variety | p-value | 0.012 | <0.001 | ||||
| Year | Treatment | Podding Biomass (kg·ha−1) | Yield Component | ||||
|---|---|---|---|---|---|---|---|
| Grain Number (grains·m−2) | Grain Weight (mg) | Yield (kg·ha−1) | HI (%) | ||||
| 2020 | Water | 0-0-0 | 1197 a | 456 a | 59 | 242 a | 17 |
| 0-40-0 | 1373 ab | 520 a | 59 | 283 a | 19 | ||
| 0-0-40 | 1317 ab | 723 ab | 62 | 408 ab | 20 | ||
| 0-40-40 | 2317 b | 1058 b | 60 | 558 b | 22 | ||
| Variety | Celera II-AU | 1405 | 916 b | 42 a | 384 b | 22 b | |
| Jade-AU | 1697 | 463 a | 77 b | 362 a | 16 a | ||
| Water | p-value | 0.024 | <0.001 | <0.001 | |||
| Variety | p-value | <0.001 | <0.001 | 0.003 | |||
| 2021 | Water | 0-0-0 | 727 a | 88 a | 50 a | 16 a | 11 a |
| 0-40-0 | 1328 abc | 372 abc | 56 ab | 168 ab | 21 b | ||
| 0-0-40 | 942 ab | 281 abc | 54 ab | 135 ab | 16 ab | ||
| 0-40-40 | 1507 bcd | 514 bc | 61 b | 251 ab | 24 b | ||
| 60-0-0 | 1032 ab | 192 ab | 54 ab | 98 ab | 20 ab | ||
| 60-40-0 | 1835 cd | 623 bc | 61 b | 320 b | 23 b | ||
| 60-0-40 | 1257 abc | 274 abc | 58 ab | 151 ab | 24 b | ||
| 60-40-40 | 2054 d | 640 c | 61 b | 360 b | 24 b | ||
| Variety | Celera II-AU | 1111 a | 380 | 40 a | 119 a | 20 | |
| Jade-AU | 1559 b | 366 | 74 b | 255 b | 21 | ||
| Water | p-value | <0.001 | <0.001 | 0.005 | 0.001 | <0.001 | |
| Variety | p-value | 0.004 | <0.001 | 0.001 | |||
| Water Treatment | Hamilton 2021 | Horsham 2020 | Horsham 2021 | Ouyen 2020 | Ouyen 2021 | Woomelang 2020 | Woomelang 2021 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | ||||||||
| 0-0-0 | 285 | 270 | 278 a | 170 | 154 | 162 a | 244 | 221 | 233 a | 124 | 80 | 102 a | 100 | 108 | 104 a | 77 | 95 | 86 a | 112 | 117 | 115 a | |||||||
| 0-40-0 | 357 | 259 | 308 ab | 208 | 200 | 204 b | 240 | 246 | 243 a | 152 | 141 | 147 b | 155 | 161 | 158 b | 133 | 159 | 146 b | 121 | 172 | 147 ab | |||||||
| 0-0-40 | 275 | 247 | 261 a | 213 | 195 | 204 b | 249 | 272 | 260 ab | 155 | 175 | 165 bc | 147 | 151 | 150 b | 114 | 130 | 122 b | 153 | 170 | 162 ab | |||||||
| 0-40-40 | 346 | 392 | 369 b | 247 | 243 | 245 c | 292 | 275 | 284 b | 175 | 204 | 190 c | 183 | 202 | 193 c | 174 | 186 | 180 c | 199 | 202 | 207 b | |||||||
| 60-0-0 | 235 | 233 | 234 a | 146 | 179 | 163 b | 105 | 133 | 119 a | |||||||||||||||||||
| 60-40-0 | 265 | 270 | 267 ab | 207 | 207 | 207 c | 186 | 181 | 183 b | |||||||||||||||||||
| 60-0-40 | 237 | 240 | 238 a | 183 | 194 | 189 c | 166 | 174 | 170 ab | |||||||||||||||||||
| 60-40-40 | 283 | 303 | 293 b | 237 | 243 | 240 d | 165 | 236 | 200 b | |||||||||||||||||||
| Mean | 316 | 292 | 304 | 210 | 198 | 204 | 256 | 258 | 257 | 152 | 150 | 151 | 170 | 181 | 175 | 125 | 143 | 134 | 151 | 173 | 162 | |||||||
| Water × Variety | p-value | 0.003 | ||||||||||||||||||||||||||
| Water | p-value | 0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | ||||||||||||||||||||
| Variety | p-value | 0.003 | 0.039 | 0.017 | ||||||||||||||||||||||||
| Water Treatment | Hamilton 2021 | Horsham 2020 | Horsham 2021 | Ouyen 2020 | Ouyen 2021 | Woomelang 2020 | Woomelang 2021 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | ||||||||
| 0-0-0 | 3.9 | 1.9 | 2.9 | 1.5 | 2.0 | 1.7 a | 1.3 | 2.0 | 1.6 a | 0.4 | 1.2 | 0.8 a | 7.5 | 12.9 | 10.2 c | 3.6 | 2.2 | 2.9 | 0.5 | 0.7 | 0.6 a | |||||||
| 0-40-0 | 5.1 | 2.1 | 3.6 | 1.8 | 2.4 | 2.1 ab | 2.9 | 3.2 | 3.1 b | 1.1 | 0.5 | 0.8 a | 3.6 | 14.0 | 8.8 bc | 2.2 | 1.9 | 2.0 | 1.8 | 1.4 | 1.6 ab | |||||||
| 0-0-40 | 4.4 | 2.6 | 3.5 | 2.3 | 3.2 | 2.7 bc | 2.8 | 2.2 | 2.5 ab | 1.5 | 1.6 | 1.6 ab | 7.3 | 14.5 | 10.9 c | 3.7 | 3.3 | 3.5 | 0.6 | 0.7 | 0.7 ab | |||||||
| 0-40-40 | 4.7 | 1.7 | 3.2 | 3.0 | 3.4 | 3.2 c | 2.6 | 3.9 | 3.2 b | 2.3 | 2.6 | 2.4 b | 5.6 | 9.9 | 7.7 abc | 3.6 | 2.8 | 3.2 | 1.1 | 1.8 | 1.4 ab | |||||||
| 60-0-0 | 1.8 | 1.8 | 1.8 ab | 2.8 | 7.1 | 5.0 a | 0.6 | 2.1 | 1.3 ab | |||||||||||||||||||
| 60-40-0 | 2.0 | 2.3 | 2.2 ab | 3.6 | 5.8 | 4.7 a | 1.6 | 1.8 | 1.7 ab | |||||||||||||||||||
| 60-0-40 | 2.4 | 2.9 | 2.6 ab | 6.0 | 8.5 | 7.2 abc | 0.6 | 1.8 | 1.2 ab | |||||||||||||||||||
| 60-40-40 | 2.7 | 2.7 | 2.7 ab | 4.8 | 8 | 6.4 ab | 1.8 | 2.2 | 2.0 b | |||||||||||||||||||
| Mean | 4.5 | 2.1 | 3.3 | 2.2 | 2.8 | 2.5 | 2.3 | 2.6 | 2.5 | 1.3 | 1.5 | 1.4 | 5.2 | 10.1 | 7.6 | 3.3 | 2.6 | 2.9 | 1.1 | 1.6 | 1.3 | |||||||
| Water × Variety | p-value | 0.012 | ||||||||||||||||||||||||||
| Water | p-value | 0.001 | 0.006 | 0.001 | <0.001 | 0.036 | ||||||||||||||||||||||
| Variety | p-value | <0.001 | 0.027 | 0.002 | <0.001 | 0.030 | ||||||||||||||||||||||
| Water Treatment | Hamilton 2021 | Horsham 2021 | Ouyen 2021 | Woomelang 2021 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | |||||
| 0-0-0 | 46.2 | 35.0 | 40.6 | 20.0 | 19.6 | 19.8 a | 60.4 | 65.2 | 62.8 | 63.6 | 71.1 | 67.4 ab | ||||
| 0-40-0 | 40.0 | 34.4 | 37.2 | 21.2 | 32.8 | 27 ab | 56.8 | 57.3 | 57.1 | 67.0 | 69.3 | 68.1 ab | ||||
| 0-0-40 | 41.5 | 34.3 | 37.9 | 25.6 | 26.8 | 26.2 ab | 60.2 | 65.2 | 62.7 | 62.4 | 69.4 | 65.9 ab | ||||
| 0-40-40 | 38.9 | 26.7 | 32.8 | 39.0 | 32.3 | 35.6 b | 56.3 | 63.4 | 59.9 | 76.8 | 74.8 | 75.8 b | ||||
| 60-0-0 | 9.6 | 23.2 | 16.4 a | 65.9 | 67.5 | 66.7 | 76.4 | 62.1 | 69.2 ab | |||||||
| 60-40-0 | 21.3 | 16.1 | 18.7 a | 59.9 | 61.9 | 60.9 | 65.0 | 63.2 | 64.1 a | |||||||
| 60-0-40 | 18.0 | 20.5 | 19.2 a | 59.0 | 62.5 | 60.7 | 72.5 | 75.9 | 74.2 ab | |||||||
| 60-40-40 | 25.3 | 25.2 | 25.2 a | 58.7 | 58.5 | 58.6 | 70.1 | 72.6 | 71.3 ab | |||||||
| Mean | 41.6 | 32.6 | 37.1 | 22.5 | 24.6 | 23.5 | 59.7 | 62.7 | 61.2 | 69.2 | 69.8 | 69.5 | ||||
| Water × Variety | p-value | 0.041 | ||||||||||||||
| Water | p-value | <0.001 | 0.025 | |||||||||||||
| Water Treatment | Hamilton 2021 | Horsham 2021 | Ouyen 2021 | Woomelang 2021 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | Celera II-AU | Jade-AU | Mean | |||||
| 0-0-0 | 20.0 | 33.0 | 26.5 | 2.1 | 2.8 | 2.5 a | 33.9 | 81.9 | 57.9 b | 2.8 | 10.7 | 6.7 a | ||||
| 0-40-0 | 32.4 | 27.1 | 29.8 | 3.4 | 7.5 | 5.4 ab | 35.8 | 72.3 | 54.1 ab | 8.2 | 10.6 | 9.4 ab | ||||
| 0-0-40 | 22.6 | 35.2 | 28.9 | 5.3 | 6.2 | 5.8 ab | 43.4 | 80.9 | 62.1 b | 6.4 | 18.2 | 12.3 ab | ||||
| 0-40-40 | 31.2 | 23.6 | 27.4 | 4.2 | 10.0 | 7.1 b | 48.8 | 73.6 | 61.2 b | 9.2 | 20.1 | 14.7 b | ||||
| 60-0-0 | 1.5 | 5.7 | 3.6 a | 21.3 | 44.8 | 33.0 a | 4.3 | 13.4 | 8.8 ab | |||||||
| 60-40-0 | 3.3 | 3.0 | 3.1 a | 28.0 | 39.5 | 33.7 a | 9.3 | 15.4 | 12.3 ab | |||||||
| 60-0-40 | 2.9 | 5.4 | 4.2 ab | 39.0 | 60.9 | 49.9 ab | 7.4 | 12.0 | 9.7 ab | |||||||
| 60-40-40 | 3.7 | 7.1 | 5.4 ab | 38.8 | 47.6 | 43.2 ab | 10.1 | 20.4 | 15.2 b | |||||||
| Mean | 26.6 | 29.7 | 28.1 | 3.3 a | 6.0 b | 4.6 | 36.1 a | 62.7 b | 49.4 | 7.2 a | 15.1 b | 11.2 | ||||
| Water | p-value | 0.001 | <0.001 | 0.005 | ||||||||||||
| Variety | p-value | <0.001 | <0.001 | <0.001 | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silwal, S.; Delahunty, A.J.; Wallace, A.J.; Norton, S.; Pang, A.; Nuttall, J.G. Water and Nitrogen Dynamics of Mungbean as a Summer Crop in Temperate Environments. Agronomy 2025, 15, 1711. https://doi.org/10.3390/agronomy15071711
Silwal S, Delahunty AJ, Wallace AJ, Norton S, Pang A, Nuttall JG. Water and Nitrogen Dynamics of Mungbean as a Summer Crop in Temperate Environments. Agronomy. 2025; 15(7):1711. https://doi.org/10.3390/agronomy15071711
Chicago/Turabian StyleSilwal, Sachesh, Audrey J. Delahunty, Ashley J. Wallace, Sally Norton, Alexis Pang, and James G. Nuttall. 2025. "Water and Nitrogen Dynamics of Mungbean as a Summer Crop in Temperate Environments" Agronomy 15, no. 7: 1711. https://doi.org/10.3390/agronomy15071711
APA StyleSilwal, S., Delahunty, A. J., Wallace, A. J., Norton, S., Pang, A., & Nuttall, J. G. (2025). Water and Nitrogen Dynamics of Mungbean as a Summer Crop in Temperate Environments. Agronomy, 15(7), 1711. https://doi.org/10.3390/agronomy15071711