
Molecular Mutation of the Coat Protein (CP) Gene in Alfalfa Mosaic Virus (AMV) and White Clover Mosaic Virus (WCMV) Combined Infection and the Role of the WCMV CP Gene When Infected with AMV
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Virus Materials
2.2. Virus Inoculation
2.3. Total RNA Isolation
2.4. Cloning of the AMV CP and WCMV CP Sequences During Co-Infection and Single Infection
2.5. Bioinformatic Analysis in Co-Infection and Single Infection of the AMV CP and WCMV CP Sequences
2.6. Determination of the Inhibitory Effect of the WCMV CP on Infection with AMV
2.7. Statistical Analysis
3. Results
3.1. Symptoms of N. benthamiana After Co-Infection with AMV and WCMV
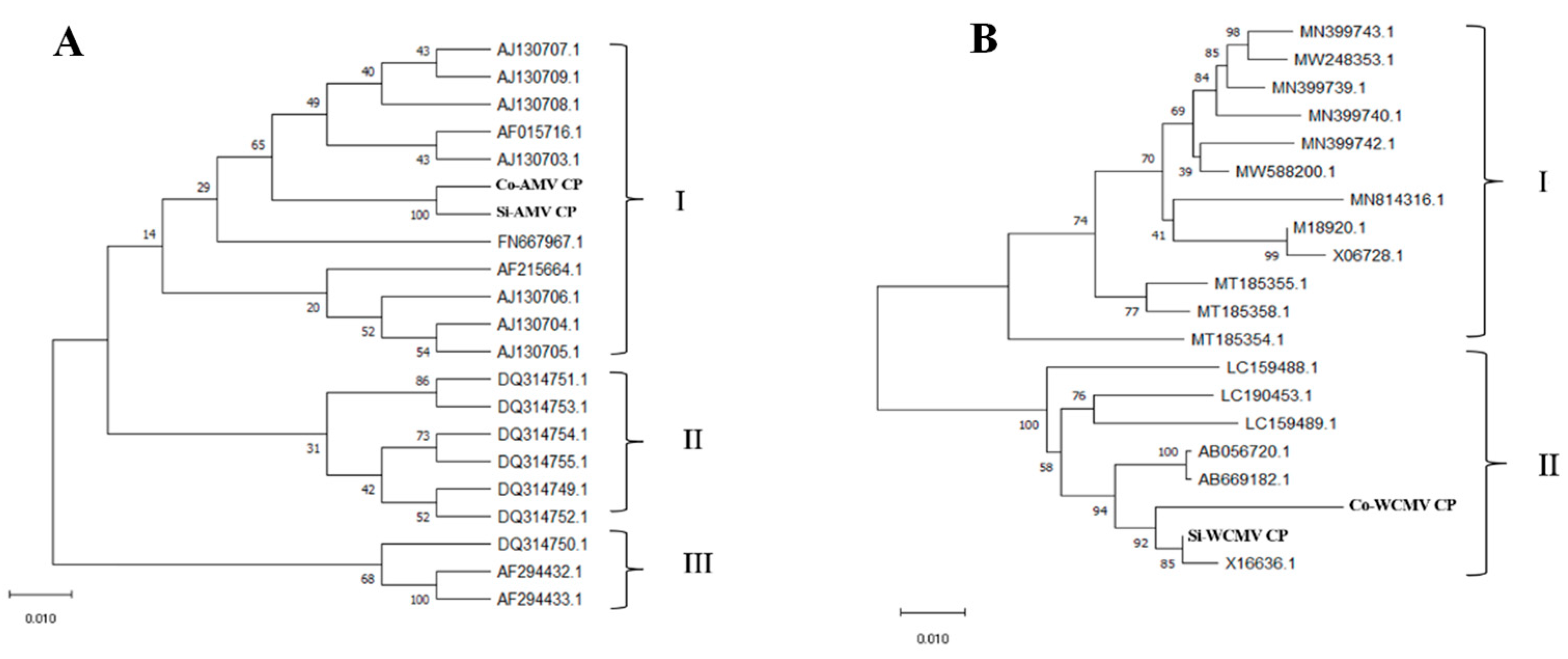
3.2. Mutation Analysis of the AMV CP and WCMV CP During Co-Infection
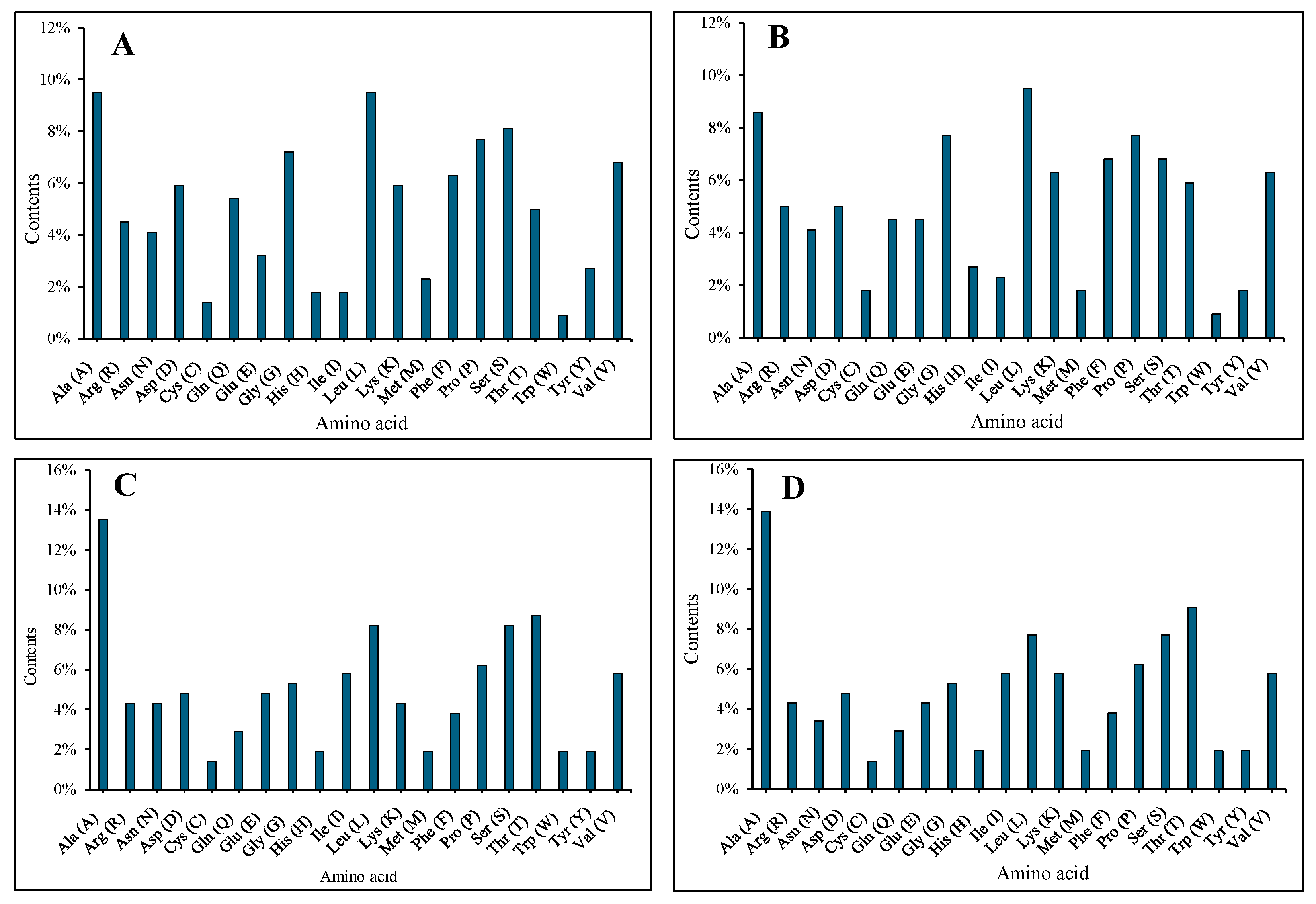
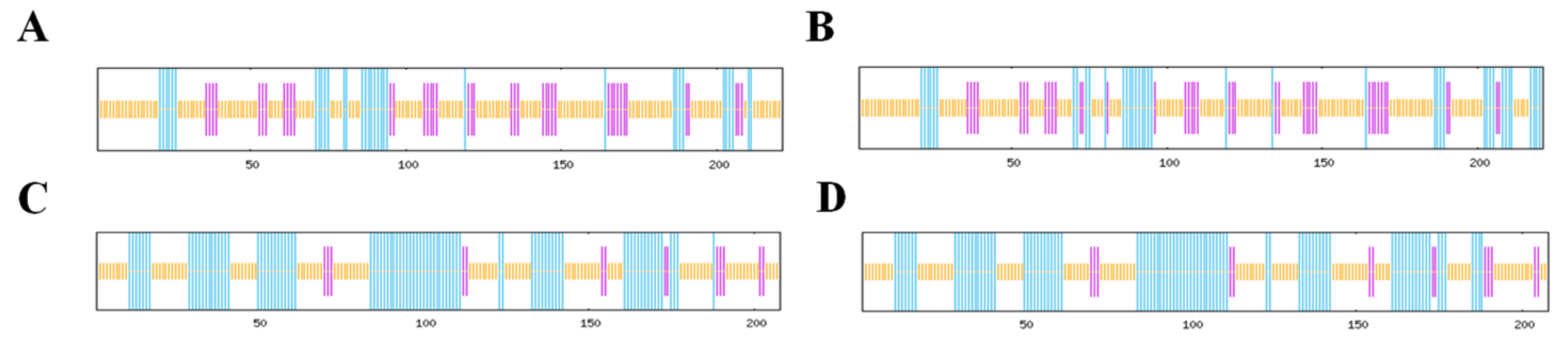
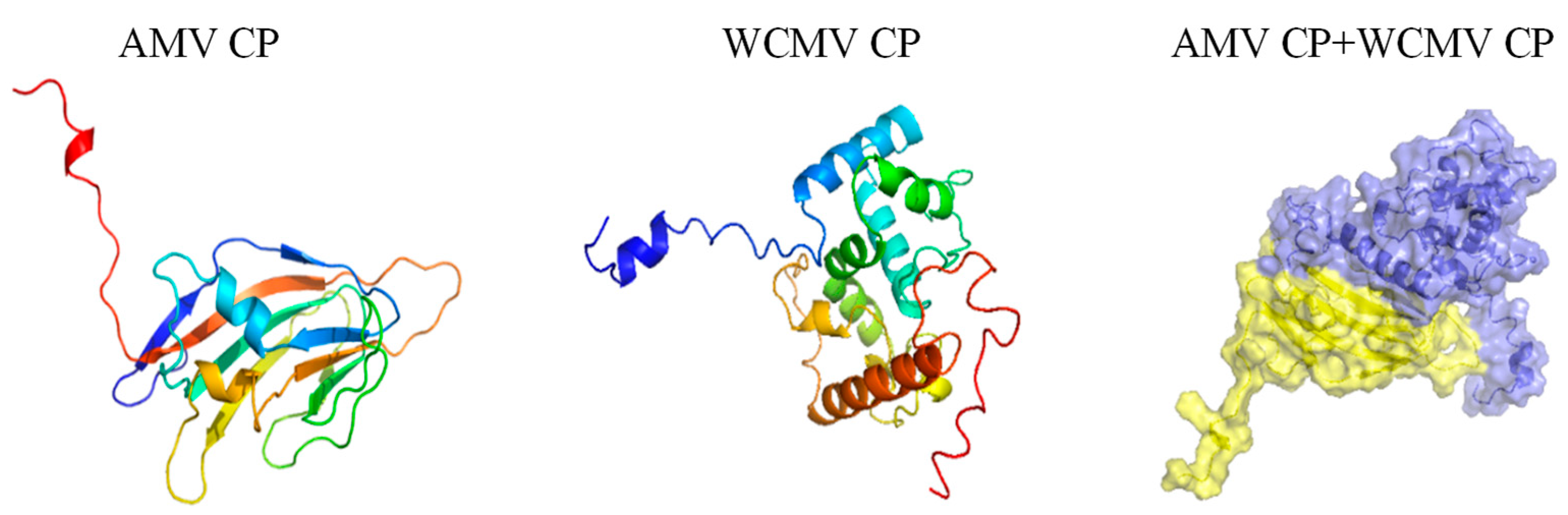
3.3. Nucleotide and Amino Acid Analyses of the AMV CP and WCMV CP DuringCo-Infection and Single Infection
3.4. Diseased Symptoms and the WCMV CP Fluorescence Intensity of N. benthamiana After Infection with Different Proportions of WCMV CP and AMV
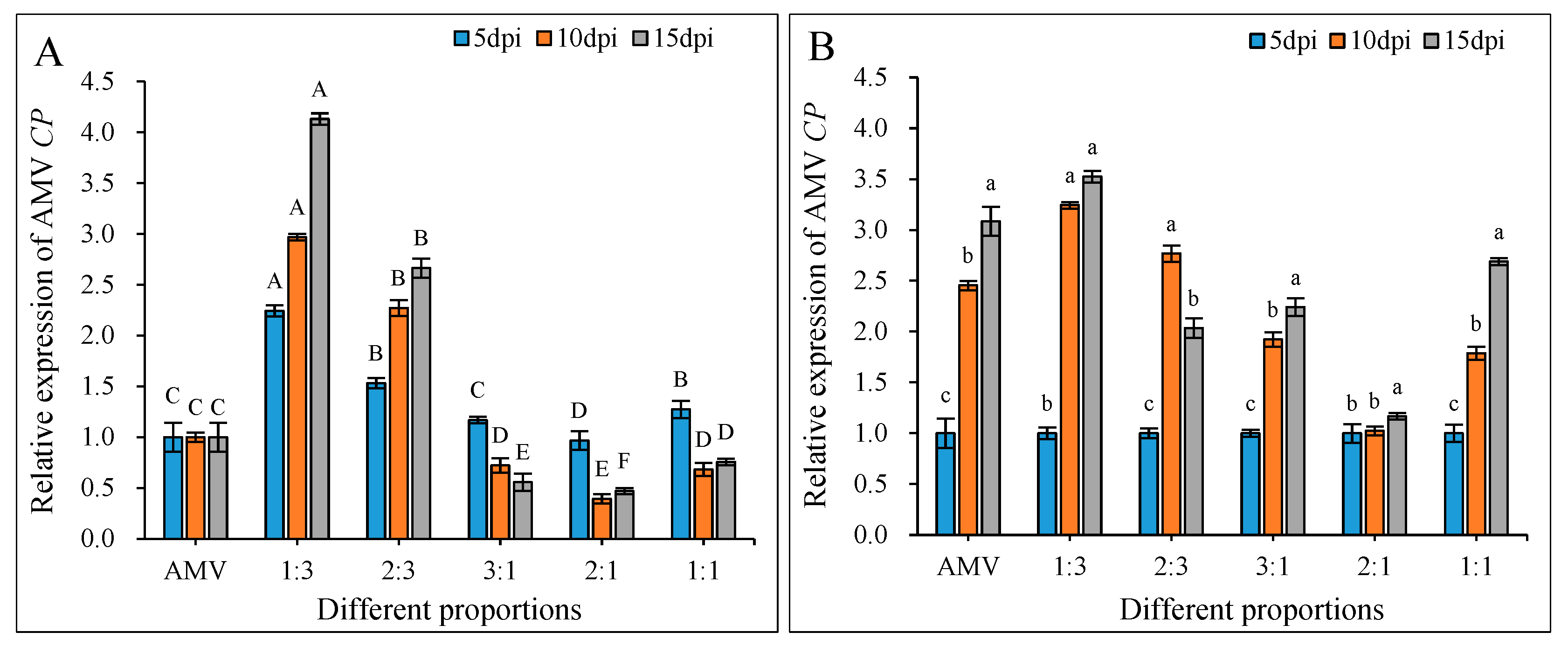
3.5. Changes in the AMV CP Relative Expression After Infection with Different Proportions of WCMV CP and AMV
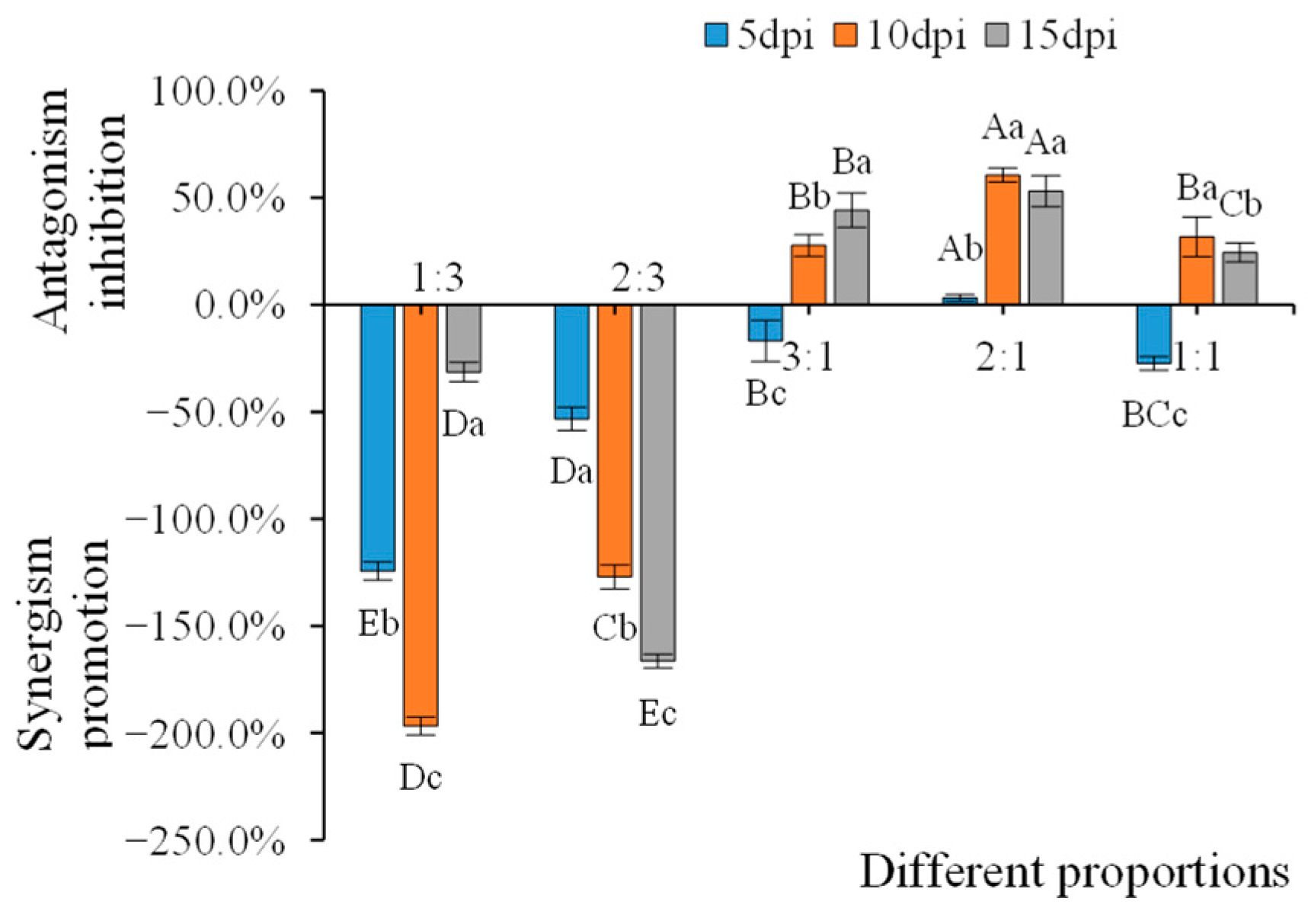
3.6. Inhibitory Effects on AMV Infection of the WCMV CP and AMV Different Proportions of Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, Q.E.; Li, Y.B. Classification and distribution of domestic species of genus Medicago in China. Grassl. Prataculture 2019, 31, 1–7. [Google Scholar]
- Wang, W.X. Countermeasures for the development of alfalfa planting industry in China. J. Beijing Univ. Agric. 2022, 37, 117–120. [Google Scholar]
- Carrasco, J.L.; Sánchez-Navarro, J.A.; Elena, S.F. Exploring the role of cellular homologous of the 30K-superfamily of plant virus movement proteins. Virus Res. 2019, 262, 54–61. [Google Scholar] [CrossRef]
- Liu, X.X.; Li, B.Y.; Aziguli, M.; Li, K.M. Harm and control of alfalfa virus disease. Rural. Sci. Technol. 2018, 10, 25–28. [Google Scholar]
- Li, J.; Shang, Q.; Liu, Y.; Dai, W.; Li, X.; Wei, S.; Hu, G.; McNeill, M.R.; Ban, L. Occurrence, distribution, and transmission of alfalfa viruses in China. Viruses 2022, 14, 1519. [Google Scholar] [CrossRef]
- Guo, Z.P.; Feng, C.S.; Zhang, J.X.; Wang, M.L.; Qu, G.; Liu, J.Y.; Guan, Y.Z.; Zhang, X.T.; Guo, Y.X.; Yan, X.B. Field resistance to alfalfa mosaic virus among 30 alfalfa varieties. Acta Prataculturae Sin. 2019, 28, 157–167. [Google Scholar]
- McLaughlin, A.A.; Hanley-Bowdoin, L.; Kennedy, G.G.; Jacobson, A.L. Vector acquisition and co-inoculation of two plant viruses influences transmission, infection, and replication in new hosts. Sci. Rep. 2022, 12, 20355. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Congdon, B.S. Australian cool-season pulse seed-borne virus research: 1. alfalfa and cucumber mosaic vi-ruses and less important viruses. Viruses 2024, 16, 144. [Google Scholar] [CrossRef]
- Bellah, H.; Seiler, N.F.; Croll, D. Divergent outcomes of direct conspecific pathogen strain interaction and plant co-infection suggest consequences for disease dynamics. Microbiol. Spectr. 2023, 11, e0444322. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, T.; Chen, Z.; Niu, J.; Cui, X.; Mao, Y.; Hassan, M.U. Occurrence, distribution, and genetic diversity of alfalfa (Medicago sativa L.) viruses in four major alfalfa producing provinces of China. Front. Microbiol. 2022, 12, 771361. [Google Scholar] [CrossRef]
- Gao, Y.; Fan, G.; Cheng, S.; Zhang, W.; Bai, Y. Evolutionary history and global spatiotemporal pattern of alfalfa mosaic virus. Front. Microbiol. 2022, 13, 1051834. [Google Scholar] [CrossRef]
- Amin, H.A.; Younes, H.A.; Shafie, R.M.; Fathallah, M.M. Molecular characterization and evolution of the resident population of some alfalfa mosaic virus (AMV) isolates in Egypt. BMC Microbiol. 2023, 23, 261. [Google Scholar] [CrossRef]
- Chen, Y.E.; Liang, Q.L.; Wei, L.X.; Wang, D.; Tian, L.; Tong, F.Y.; Zhang, G.Y. Study on the epidemic factors of alfalfa mosaic virus and white clover mosaic virus co-infection Nicotiana benthamiana. Pratacultural Sci. 2023, 40, 90–100. [Google Scholar]
- Liang, Q.L.; Wei, L.X.; Xu, B.L.; Calderón-Urrea, A.; Xiang, D. Study of viruses co-infecting white clover (Trifolium repens) in China. J. Integr. Agric. 2017, 16, 1990–1998. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Wang, J.Y.; Xie, K.Z.; Wang, R.Y.; Yang, G.; Guo, Z.H.; Qiu, Y. A study of viral coat protein accumulation in lily chloroplasts from mixed virus infections of Lily mottle virus and cucumber mosaic virus. Plant Pathol. 2019, 68, 261–268. [Google Scholar] [CrossRef]
- Cheng, S.F.; Liang, Q.L.; Wei, L.X.; Sang, X.W.; Jiang, Y.L. Detection of alfalfa mosaic virus and white clover mosaic virus in alfalfa and their effects on physiological and biochemical characteristics of alfalfa plants. Acta Prataculturae Sin. 2020, 29, 140–149. [Google Scholar]
- Chen, Y.E.; Liang, Q.L.; Wei, L.X.; Zhou, X. Double infection of Nicotiana benthamiana with AMV and WCMV increases both virus concentrations and synergistically changes both host organelle ultrastructure and chlorophyll content. Microb. Pathog. 2024, 196, 106956. [Google Scholar] [CrossRef]
- Simón, D.; Cristina, J.; Musto, H. Nucleotide composition and codon usage across viruses and their respective hosts. Front. Microbiol. 2021, 12, 646300. [Google Scholar] [CrossRef]
- González-Pérez, E.; Chiquito-Almanza, E.; Villalobos-Reyes, S.; Canul-Ku, J.; Anaya-López, J.L. Diagnosis and characterization of plant viruses using HTS to support virus management and tomato breeding. Viruses 2024, 16, 888. [Google Scholar] [CrossRef] [PubMed]
- García-Arenal, F.; Fraile, A.; Malpica, J.M. Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 2001, 39, 157–186. [Google Scholar] [CrossRef]
- Lagzian, A.; Ghorbani, A.; Tabein, S.; Riseh, R.S. Genetic variations and gene expression profiles of rice black-streaked dwarf virus (RBSDV) in different host plants and insect vectors: Insights from RNA-Seq analysis. BMC Genom. 2024, 25, 736. [Google Scholar] [CrossRef]
- Yoshida, T.; Ishikawa, M.; Toki, S.; Ishibashi, K. Heritable tissue-culture-free gene editing in Nicotiana benthamiana through viral delivery of spcas9 and SgRNA. Plant Cell Physiol. 2024, 65, 1743–1750. [Google Scholar] [CrossRef]
- Verdin, E.; Wipf-Scheibel, C.; Gognalons, P.; Aller, F.; Jacquemond, M.; Tepfer, M. Sequencing viral siRNAs to identify previously undescribed viruses and viroids in a panel of ornamental plant samples structured as a matrix of pools. Virus Res. 2017, 241, 19–28. [Google Scholar] [CrossRef]
- Gupta, T.; Raj, S.K.; Singhal, T.; Srivastava, A. Phylogenetic and recombination analysis of yellow mosaic disease in soybean plant. J. Plant Dis. Sci. 2022, 17, 28–38. [Google Scholar] [CrossRef]
- Ivanov, K.I.; Mäkinen, K. Coat proteins, host factors and plant viral replication. Curr. Opin. Virol. 2012, 2, 712–718. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Gao, Q.; Wang, H.J.; Yue, J.Y.; An, D.R.; Li, B.; Yan, F.F.; Carmen, S.M.; Zhao, Y.Z.; Zhou, H.Y.; et al. Syn-tasiRNAs targeting the coat protein of potato virus Y confer antiviral resistance in Nicotiana benthamiana. Plant Signal. Behav. 2024, 19, 2358270. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Li, S.; Ji, Y.; Sun, F.; Zou, B. Dicer-like 2 plays a crucial role in rice stripe virus coat protein-mediated virus resistance in Arabidopsis. Viruses 2023, 15, 2239. [Google Scholar] [CrossRef]
- Martínez, C.; López, C.; Pallás, V.; Pallás, F.; Aparicio, L. Role of the coat (CP), movement (MP) and 2b proteins of parietaria motte virus (PMoV) as pathogen determinants in Nicotiana benthamiana plants. Eur. J. Plant Pathol. 2025, 172, 291–304. [Google Scholar] [CrossRef]
- Jiang, J.; Yu, E.; Nihranz, C.T.; Prakash, V.; Varsani, S.; Casteel, C.L. Engineering aphid transmission of foxtail mosaic virus in the presence of potyvirus helper component proteinase through coat protein modifications. J. Gen. Virol. 2023, 104, 10. [Google Scholar] [CrossRef]
- Bendahmane, M.; Szecsi, J.; Chen, I.; Berg, R.H.; Beachy, R.N. Characterization of mutant tobacco mosaic virus coat protein that interferes with virus cell-to-cell movement. Proc. Natl. Acad. Sci. USA 2002, 99, 3645–3650. [Google Scholar] [CrossRef]
- Shi, J.; Wang, X.; Wang, E. Mycorrhizal symbiosis in plant growth and stress adaptation: From genes to ecosystems. Annu. Rev. Plant Biol. 2023, 74, 569–607. [Google Scholar] [CrossRef]
- Lukhovitskaya, N.; Brown, K.; Hua, L.; Pate, A.E.; Carr, J.P.; Firth, A.E. A novel ilarvirus protein CP-RT is expressed via stop codon readthrough and suppresses RDR6-dependent RNA silencing. PLoS Pathog. 2024, 20, e1012034. [Google Scholar] [CrossRef]
- Suman, R.; Rani, A.; Rishi, N.; Dhir, S.; Hallan, V.; Chandel, V. First report of apple stem grooving virus infection in loquat from India. Virus Dis. 2022, 33, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Asurmendi, S.; Berg, R.H.; Smith, T.J.; Bendahmane, M.; Beachy, R.N. Aggregation of TMV CP plays a role in CP functions and in coat-protein-mediated resistance. Virology 2007, 366, 98–106. [Google Scholar] [CrossRef]
- Hosseini, H.; Mehrvar, M.; Zakiaghl, M.; Siampour, M. Comparative genetic diversity of potato virus Y populations based on coat protein gene. Acta Virol. 2017, 61, 161–174. [Google Scholar] [CrossRef]
- Pinczés, D.; Sáray, R.; Nemes, K.; Palkovics, L.; Salánki, K. Viral coat proteins decrease the gene silencing activity of cognate and heterologous viral suppressors. Sci. Rep. 2024, 14, 31008. [Google Scholar] [CrossRef]
- Villar-Álvarez, D.; Pallás, V.; Elena, S.F.; Sánchez-Navarro, J.A. An evolved 5’ untranslated region of alfalfa mosaic virus allows the RNA transport of movement-defective variants. J. Virol. 2022, 96, e0098822. [Google Scholar] [CrossRef]
- Jin, L.L. Complete genomic sequence and biological characteristics of a new alfalfa mosaic virus isolate. Zhejiang Univ. Technol. 2011. [Google Scholar]
- Wetter, C. Partial purification of some elongated plant viruses and their use as antigens in immunization by means of Freund’s adjuvant. Arch. Mikrobiol. 1960, 37, 278. [Google Scholar]
- Janeczko, A.; Dziurka, M.; Gullner, G.; Kocurek, M.; Rys, M.; Saja, D.; Skoczowski, A.; Tóbiás, I.; Kornas, A.; Barna, B. Comparative studies of compatible and incompatible pepper tobamovirus interactions and the evaluation of effects of 24-epibrassinolide. Photosynthetica 2018, 56, 763–775. [Google Scholar] [CrossRef]
- Niu, E.B.; Liu, H.; Zhou, H.; Luo, L.; Wu, Y.; Andika, I.B.; Sun, L. Autophagy inhibits intercellular transport of citrus leaf blotch virus by targeting viral movement protein. Viruses 2021, 13, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Alcaide, C.; Sardanyés, J.; Elena, S.F.; Gómez, P. Increasing temperature alters the within-host competition of viral strains and influences virus genetic variability. Virus Evol. 2021, 7, veab017. [Google Scholar] [CrossRef]
- Agüero, J.; Gómez-Aix, C.; Sempere, R.N.; García-Villalba, J.; García-Núñez, J.; Hernando, Y.; Aranda, M.A. Stable and broad spectrum cross-protection against pepino mosaic virus attained by mixed infection. Front. Plant Sci. 2018, 9, 1810. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.; Millar, A.H. Proteomics in evolutionary ecology. J. Proteom. 2016, 135, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Dillenberger, M.S.; Wei, N.; Tennessen, J.A.; Ashman, T.L.; Liston, A. Plastid genomes reveal recurrent formation of allopolyploid Fragaria. Am. J. Bot. 2018, 105, 862–874. [Google Scholar] [CrossRef]
- Sánchez-Martín, J.; Steuernagel, B.; Ghosh, S.; Herren, G.; Hurni, S.; Adamski, N.; Vrána, J. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 2016, 17, 221. [Google Scholar] [CrossRef]
- Weber, P.H.; Bujarski, J.J. Multiple functions of capsid proteins in (+) stranded RNA viruses during plant-virus interactions. Virus Res. 2015, 196, 140–149. [Google Scholar] [CrossRef]
- Agranovsky, A. Enhancing capsid proteins capacity in plant virus-vector interactions and virus transmission. Cells 2021, 10, 90. [Google Scholar] [CrossRef]
- He, C.; Xing, F.; Zhao, X.; Li, S.; Zhan, B.; Liu, Z.; Xu, T. The coat protein of the ilarvirus prunus necrotic ringspot virus mediates long-distance movement. J. Gen. Virol. 2023, 104, 1009. [Google Scholar] [CrossRef]
- Maachi, A.; Nagata, T.; Silva, J.M.F. Date palm virus A: First plant virus found in date palm trees. Virus Genes. 2020, 56, 792–795. [Google Scholar] [CrossRef]
- Vinaykumar, H.D.; Hiremath, S.; Nandan, M.; Muttappagol, M.; Reddy, M.; Venkataravanappa, V.; Shankarappa, K.S. Genome sequencing of cucumber mosaic virus (CMV) isolates infecting chilli and its interaction with host ferredoxin protein of different host for causing mosaic symptoms. 3 Biotech 2023, 13, 361. [Google Scholar] [CrossRef] [PubMed]
- Purohit, R.; Kumar, S.; Hallan, V. Screening of potential inhibitor against coat protein of apple chlorotic leaf spot virus. Cell Biochem. Biophys. 2018, 76, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Alcaide, C.; Aranda, M.A. Determinants of persistent patterns of pepino mosaic virus mixed infections. Front. Microbiol. 2021, 12, 694492. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.J.; Yang, X.; Yang, L.L.; Li, Q.L.; Liu, X.M.; Han, X.Y.; Gu, Q.S. Interaction between cucumber green mottle mosaic virus MP and CP promotes virus systemic infection. Mol. Plant Pathol. 2023, 24, 208–220. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, S.; Jiang, L.; Yang, Q.; Luo, L.; Jiang, J.; Malichan, S. Pitaya virus x coat protein acts as an RNA silencing suppressor and can be used as a specific target for detection using RT-LAMP. Plant Dis. 2023, 107, 3378–3382. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Primer Sequences (5′-3′) | Product Size (bp) | Amplification Efficiency | R2 | Linear Regression Equation | Description |
|---|---|---|---|---|---|---|---|
| AMV CP | AMV CP-F | ATGAGTTCTTCACAAAAGAAAGCTGGTG | 666 | Primers for cloning | |||
| AMV CP-R | TGACGATCAAGATCGTCAGCTTCG | ||||||
| Si-AMV CP | qSi-AMV CP-F | GCATCCCTAGGGGCATTCATGCA | 208 | 0.9849 | 0.9939 | Y = −3.3586X + 33.941 | Primers for RT-qPCR |
| qSi-AMV CP-R | ATCATTGATCGGTAATGGGCCGTT | ||||||
| WCMV CP | WCMV CP-F | ATGGCAACCACCACAGCAAC | 621 | Primers for cloning | |||
| WCMV CP-R | GGGATAGGTAATAAGGGGACC | ||||||
| Si-WCMV CP | qSi-WCMV CP-F | AAACTCGAGCATGGACTTCACTACTTTA | 198 | 0.9904 | 0.9898 | Y = −3.3452X + 32.064 | Primers for RT-qPCR |
| qSi-WCMV CP-R | CAGGTACCCTGAAATTTTATTAAACAGAAAGCACACAC | ||||||
| OESi-WCMV CP-R | AGAACACGGGGGACGAGCTCATGGCAACCACCACAGCAACCAC | 621 | Primers for pCEGFP | ||||
| OESi-WCMV CP-R | ACCATGGTGTCGACTCTAGAGGGATAGGTAATAAGGGGACCAAT | ||||||
| 25S rRNA | q25S rRNA-F | AAGGCCGAAGAGGAGAAAGGT | 206 | 0.9986 | 0.9882 | Y = −3.3252X + 27.819 | Primers for RT-qPCR |
| q25S rRNA-R | CGTCCCTTAGGATCGGCTTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Liang, Q.; Wei, L.; Zhou, X.; Lai, S. Molecular Mutation of the Coat Protein (CP) Gene in Alfalfa Mosaic Virus (AMV) and White Clover Mosaic Virus (WCMV) Combined Infection and the Role of the WCMV CP Gene When Infected with AMV. Agronomy 2025, 15, 1646. https://doi.org/10.3390/agronomy15071646
Chen Y, Liang Q, Wei L, Zhou X, Lai S. Molecular Mutation of the Coat Protein (CP) Gene in Alfalfa Mosaic Virus (AMV) and White Clover Mosaic Virus (WCMV) Combined Infection and the Role of the WCMV CP Gene When Infected with AMV. Agronomy. 2025; 15(7):1646. https://doi.org/10.3390/agronomy15071646
Chicago/Turabian StyleChen, Yinge, Qiaolan Liang, Liexin Wei, Xin Zhou, and Shiyu Lai. 2025. "Molecular Mutation of the Coat Protein (CP) Gene in Alfalfa Mosaic Virus (AMV) and White Clover Mosaic Virus (WCMV) Combined Infection and the Role of the WCMV CP Gene When Infected with AMV" Agronomy 15, no. 7: 1646. https://doi.org/10.3390/agronomy15071646
APA StyleChen, Y., Liang, Q., Wei, L., Zhou, X., & Lai, S. (2025). Molecular Mutation of the Coat Protein (CP) Gene in Alfalfa Mosaic Virus (AMV) and White Clover Mosaic Virus (WCMV) Combined Infection and the Role of the WCMV CP Gene When Infected with AMV. Agronomy, 15(7), 1646. https://doi.org/10.3390/agronomy15071646