

Shift in the Reproductive Strategies of Phragmites australis Under Combined Influences of Salinity and Tidal Level Changes
 ,
,
Abstract
1. Introduction
2. Materials and Methods
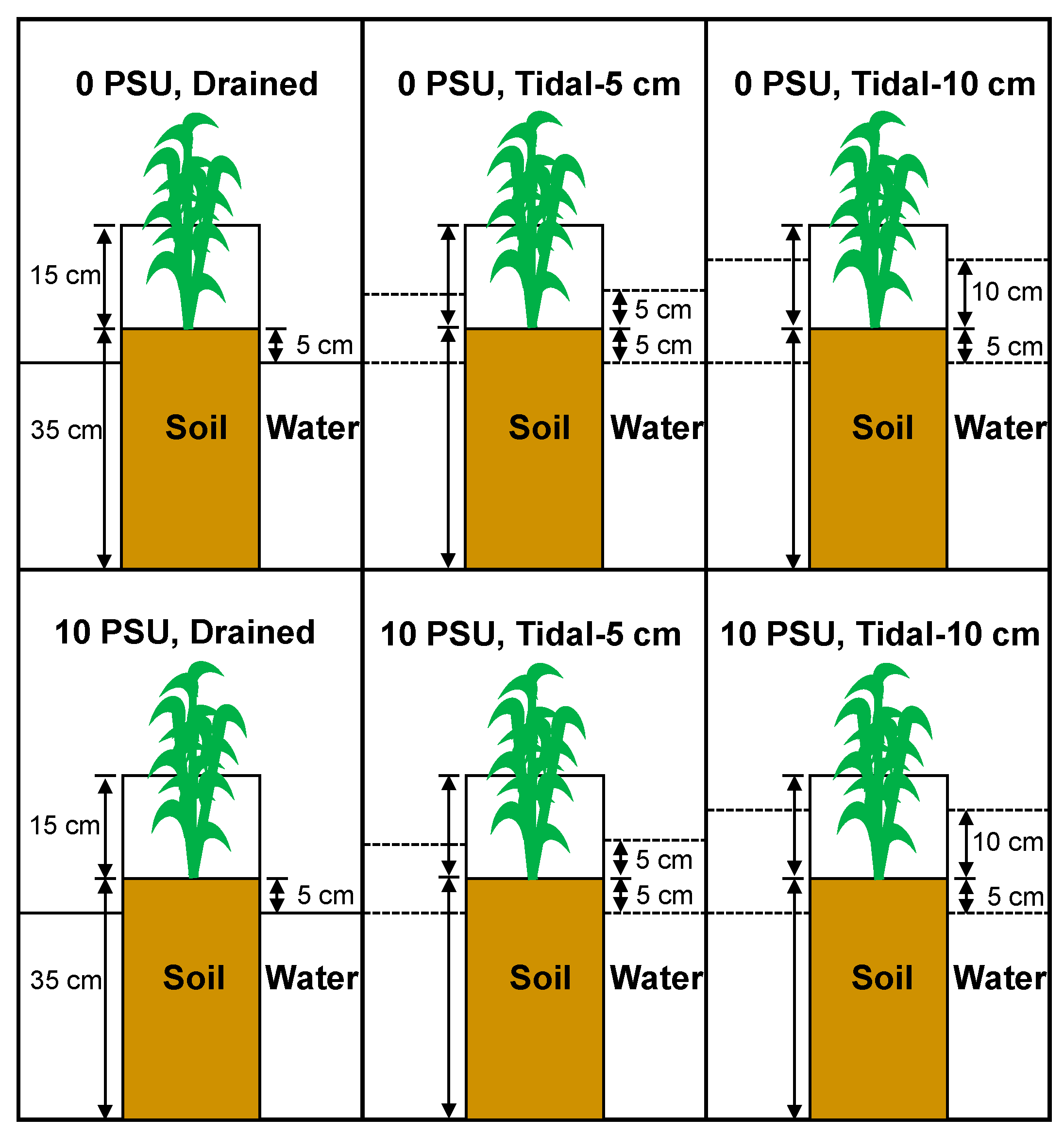
2.1. Plant Material and Experimental Design
2.2. Data Analysis
3. Results
3.1. Effects of Salinity and Tidal Level on Number of Flowering Shoots
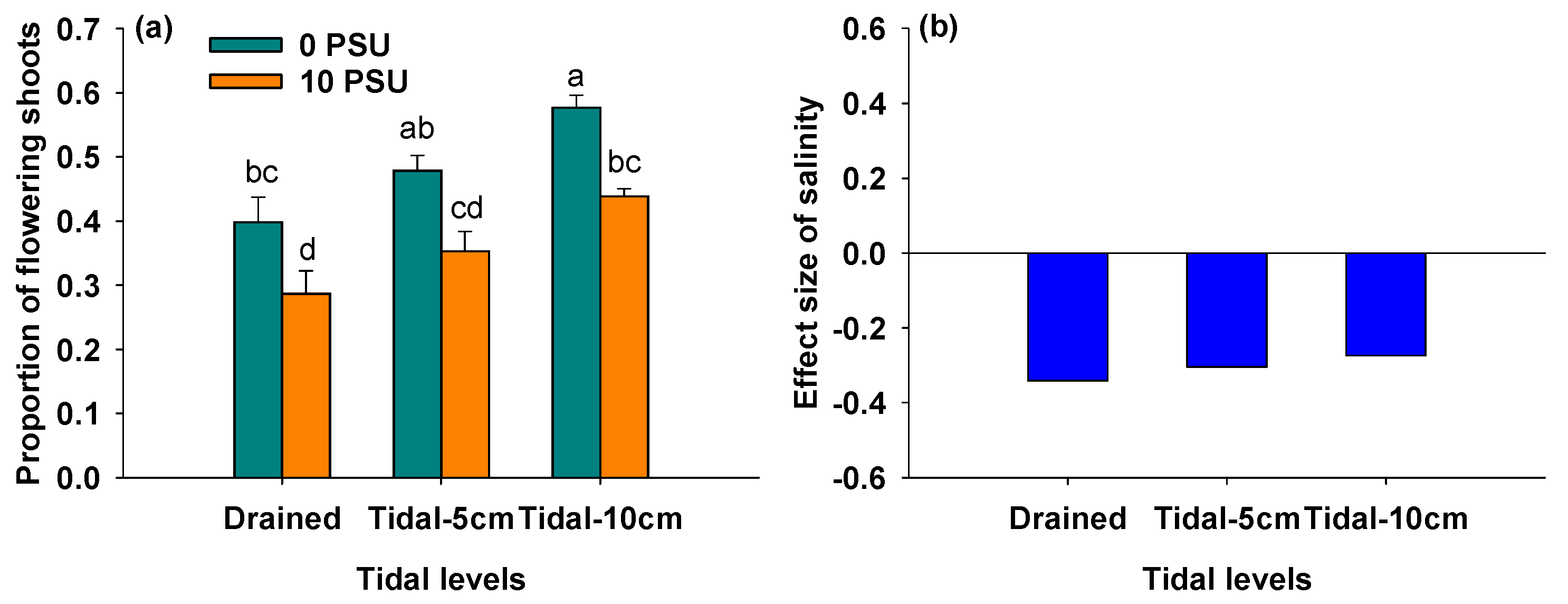
3.2. Effects of Salinity and Tidal Level on Proportion of Flowering Shoots
3.3. Effects of Salinity and Tidal Level on Inflorescence Biomass
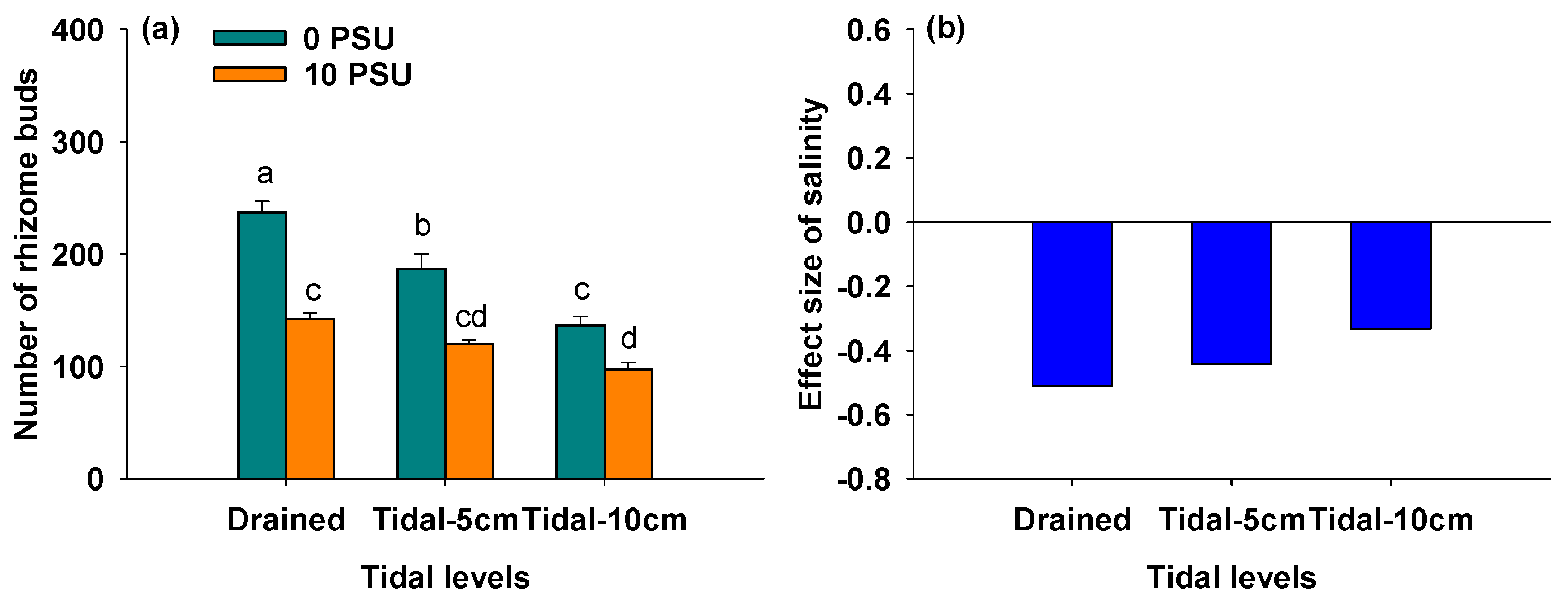
3.4. Effects of Salinity and Tidal Level on Number of Rhizome Buds
3.5. Effects of Salinity and Tidal Level on Rhizome Biomass
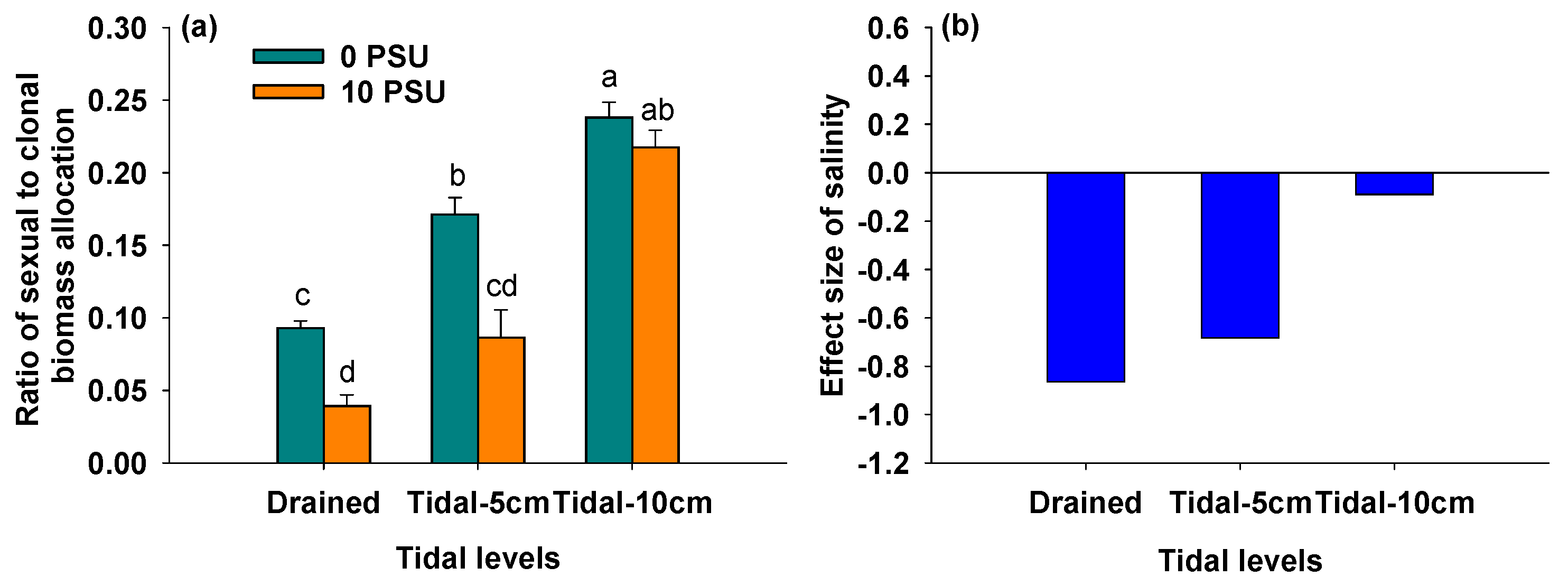
3.6. Effects of Salinity and Tidal Level on Ratio of Sexual to Clonal Biomass Allocation
4. Discussion
4.1. Reproductive Allocation in P. australis Under Varying Salinity Conditions
4.2. Reproductive Allocation in P. australis Across Different Tidal Levels
4.3. Interaction of Salinity and Tidal Level on Reproductive Allocation in P. australis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Milke, J.; Galczynska, M.; Wróbel, J. The importance of biological and ecological properties of Phragmites australis (cav.) Trin. ex Steud., in phytoremendiation of aquatic ecosystems-the review. Water 2020, 12, 1770. [Google Scholar] [CrossRef]
- Haldan, K.; Kuprina, K.; Haase, M.I.; Kieckhäfer, F.; Schade, L.; Schmoldt, J.; Schock, L.S.; Stein, M.; Wille, A.; Schnittler, M.; et al. Choose wisely: Great variation among genotypes of promising paludiculture crop Phragmites australis. Plants 2023, 12, 1045. [Google Scholar] [CrossRef]
- Hansson, P.A.; Fredriksson, H. Use of summer harvested common reed (Phragmites australis) as nutrient source for organic crop production in Sweden. Agric. Ecosyst. Environ. 2004, 102, 365–375. [Google Scholar] [CrossRef]
- Yu, J.H.; Zhong, J.C.; Zhang, Y.S.; Ding, H.; Chen, C.; Zheng, X.Z.; Xu, M.X.; Zhang, Y.L. Fine-scale remobilization of phosphorus by rooted macrophytes (Phragmites australis) growth in lake sediments: Evidence from a holistic growth period simulation study. J. Soils Sediments 2020, 20, 1782–1792. [Google Scholar] [CrossRef]
- Cui, L.J.; Pan, X.; Li, W.; Zhang, X.D.; Liu, G.F.; Song, Y.B.; Yu, F.H.; Prinzing, A.; Cornelissen, J.H.C. Phragmites australis meets Suaeda salsa on the “red beach”: Effects of an ecosystem engineer on salt-marsh litter decomposition. Sci. Total Environ. 2019, 693, 133477. [Google Scholar] [CrossRef]
- Yu, X.Y.; Ye, S.Y.; Olsson, L.; Wei, M.J.; Brix, H. In-situ CO2 partitioning measurements in a Phragmites australis wetland: Understanding carbon loss through ecosystem respiration. Wetlands 2020, 40, 901–914. [Google Scholar] [CrossRef]
- Li, X.Y.; Jin, K.; Qin, P.; Liu, C.X.; Zhu, X.Z.; Zhang, Y.Y.; Zong, Q.L. Enhancement effect of Phragmites australis roots on soil shear strength in the Yellow River Delta. Sustainability 2024, 16, 10657. [Google Scholar] [CrossRef]
- Köbbing, J.F.; Beckmann, V.; Thevs, N.; Peng, H.; Zerbe, S. Investigation of a traditional reed economy (Phragmites australis) under threat: Pulp and paper market, values and netchain at Wuliangsuhai Lake, Inner Mongolia, China. Wetl. Ecol. Manag. 2016, 24, 357–371. [Google Scholar] [CrossRef]
- Vazic, T.; Svircek, Z.; Dulic, T.; Krstic, K.; Obreht, I. Potential for energy production from reed biomass in the Vojvodina region (north Serbia). Renew. Sustain. Energy Rev. 2015, 48, 670–680. [Google Scholar] [CrossRef]
- Rezania, S.; Park, J.; Rupani, P.F.; Darajeh, N.; Xu, X.; Shahrokhishahraki, R. Phytoremediation potential and control of Phragmites australis as a green phytomass: An overview. Environ. Sci. Pollut. Res. 2019, 26, 7428–7441. [Google Scholar] [CrossRef]
- Beyzi, S.B.; Ülger, İ.; Konca, Y. Chemical, fermentative, nutritive and anti-nutritive composition of common reed (Phragmites australis) plant and silage. Waste Biomass Valorization 2023, 14, 927–936. [Google Scholar] [CrossRef]
- He, H.Q.; Li, X.Y.; Li, T.L. The sustainable development of wetlands and agriculture: A literature review. Agronomy 2025, 15, 746. [Google Scholar] [CrossRef]
- Vallejo-Marín, M.; Dorken, M.E.; Barrett, S.C.H. The ecological and evolutionary consequences of clonality for plant mating. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 193–213. [Google Scholar] [CrossRef]
- Yang, P.; Huang, L.; He, S.N.; Zeng, X.H.; Chen, Y.Y.; Wang, H.M. Adaptive strategies employed by clonal plants in heterogeneous patches. Forests 2023, 14, 1648. [Google Scholar] [CrossRef]
- Warren, R.J., II; Mokadam, C. Asexuality and species invasion. Biodivers. Conserv. 2025, 34, 29–43. [Google Scholar] [CrossRef]
- Honnay, O.; Jacquemyn, H. A meta-analysis of the relation between mating system, growth form and genotypic diversity in clonal plant species. Evol. Ecol. 2008, 22, 299–312. [Google Scholar] [CrossRef]
- Lin, C.H.; Miriti, M.N.; Goodell, K. Demographic consequences of greater clonal than sexual reproduction in Dicentra canadensis. Ecol. Evol. 2016, 6, 3871–3883. [Google Scholar] [CrossRef]
- Wang, X.; Feng, Q.H.; Zeng, Z.H.; Zhang, Z.Q.; Cai, J.; Chen, G.; Li, D.Z.; Wang, H.; Zhou, W. Effects of mode of reproduction on genetic polymorphism and divergence in wild yams (Dioscoreaceae: Dioscorea). Plant Divers. 2025, 47, 136–147. [Google Scholar] [CrossRef]
- Barrett, S.C.H. Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 8859–8866. [Google Scholar] [CrossRef]
- Becker, A.; Chen, X.; Dresselhaus, T.; Gutsche, N.; Müller-Schüssele, S.J.; Sprunck, S.; Theissen, G.; de Vries, S.; Zachgo, S. Sexual reproduction in land plants: An evolutionary perspective. Plant Reprod. 2025, 38, 12. [Google Scholar] [CrossRef]
- Boavida, L.C.; Vieira, A.M.; Becker, J.D.; Feijó, J.A. Gametophyte interaction and sexual reproduction: How plants make a zygote. Int. J. Dev. Biol. 2005, 49, 615–632. [Google Scholar] [CrossRef] [PubMed]
- Hubert, B.; Leprince, O.; Buitink, J. Sleeping but not defenceless: Seed dormancy and protection. J. Exp. Bot. 2024, 75, 6110–6124. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, S.; Meirmans, P.G.; Kirkendall, L.R. The costs of sex: Facing real-world complexities. Q. Rev. Biol. 2012, 87, 19–40. [Google Scholar] [CrossRef]
- Stenc, J.; Janosík, L.; Matousková, E.; Hadrava, J.; Mikát, M.; Janovsky, Z. Pollinator visitation closely tracks diurnal patterns in pollen release. Am. J. Bot. 2023, 110, e16179. [Google Scholar] [CrossRef]
- Begum, K.; Hasan, N.; Shammi, M. Selective biotic stressors’ action on seed germination: A review. Plant Sci. 2024, 346, 112156. [Google Scholar] [CrossRef]
- Liu, L.; Zuo, S.N.; Ma, M.Y.; Li, J.H.; Guo, L.Z.; Huang, D. Appropriate nitrogen addition regulates reproductive strategies of Leymus chinensis. Glob. Ecol. Conserv. 2021, 27, e01599. [Google Scholar] [CrossRef]
- Kottler, E.J.; Gedan, K.B. Sexual reproduction is light-limited as marsh grasses colonize maritime forest. Am. J. Bot. 2022, 109, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; He, L.; Zhang, H.; Urrutia-Cordero, P.; Ekvall, M.K.; Hollander, J.; Hansson, L.A. Climate warming and heat waves affect reproductive strategies and interactions between submerged macrophytes. Glob. Change Biol. 2017, 23, 108–116. [Google Scholar] [CrossRef]
- Wang, Z.W.; Xie, L.N.; Prather, C.M.; Guo, H.Y.; Han, G.D.; Ma, C.C. What drives the shift between sexual and clonal reproduction of Caragana stenophylla along a climatic aridity gradient? BMC Plant Biol. 2018, 18, 91. [Google Scholar] [CrossRef]
- Silvertown, J. The evolutionary maintenance of sexual reproduction: Evidence from the ecological distribution of asexual reproduction in clonal plants. Int. J. Plant Sci. 2008, 169, 157–168. [Google Scholar] [CrossRef]
- Nevermann, H.; AghaKouchak, A.; Shokri, N. Sea level rise implications on future inland migration of coastal wetlands. Glob. Ecol. Conserv. 2023, 43, e02421. [Google Scholar] [CrossRef]
- Xian, X.X.; Pang, M.Y.; Zhang, J.L.; Zhu, M.K.; Kong, F.L.; Xi, M. Assessing the effect of potential water and salt intrusion on coastal wetland soil quality: Simulation study. J. Soils Sediments 2019, 19, 2251–2264. [Google Scholar] [CrossRef]
- Sadai, S.; Ranganathan, M.; Nauels, A.; Nicholls, Z.; Merner, D.; Dahl, K.; Licker, R.; Ekwurzel, B. Estimating the sea level rise responsibility of industrial carbon producers. Environ. Res. Lett. 2025, 20, 044012. [Google Scholar] [CrossRef]
- IPCC. IPCC Summary for policymakers. In Climate Change 2023; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Talke, S.A.; Jay, D.A. Changing tides: The role of natural and anthropogenic factors. Annu. Rev. Mar. Sci. 2020, 12, 121–151. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Biemond, B.; van Keulen, D.; Huismans, Y.; van Westen, R.M.; de Swart, H.E.; Dijkstra, H.A.; Kranenburg, W.M. Global increases of salt intrusion in estuaries under future environmental conditions. Nat. Commun. 2025, 16, 3444. [Google Scholar] [CrossRef] [PubMed]
- Solohin, E.; Widney, S.E.; Craft, C.B. Declines in plant productivity drive loss of soil elevation in a tidal freshwater marsh exposed to saltwater intrusion. Ecology 2020, 101, e03148. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Pennings, S.C. Response and recovery of low-salinity marsh plant communities to presses and pulses of elevated salinity. Estuaries Coasts 2019, 42, 708–718. [Google Scholar] [CrossRef]
- Ghirardelli, A.; Straffelini, E.; Park, E.; D’Agostino, V.; Masin, R.; Tarolli, P. Global impact of seawater intrusion on coastal agriculture. Environ. Res. Lett. 2025, 20, 013005. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Gorai, M.; Ennajeh, M.; Khemira, H.; Neffati, M. Influence of NaCl-salinity on growth, photosynthesis, water relations and solute accumulation in Phragmites australis. Acta Physiol. Plant. 2011, 33, 963–971. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Matthew, C.; Uddin, M.J.; Bayazid, K.N. Salinity-induced reduction in root surface area and changes in major root and shoot traits at the phytomer level in wheat. J. Exp. Bot. 2016, 67, 3719–3729. [Google Scholar] [CrossRef] [PubMed]
- Rolletschek, H.; Hartzendorf, T. Effects of salinity and convective rhizome ventilation on amino acid and carbohydrate patterns of Phragmites australis populations in the Neusiedler See region of Austria and Hungary. New Phytol. 2000, 146, 95–105. [Google Scholar] [CrossRef]
- Negrao, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ding, Z.; Bachofen, C.; Lou, Y.J.; Jiang, M.; Tang, X.G.; Lu, X.G.; Buchmann, N. The effect of saline-alkaline and water stresses on water use efficiency and standing biomass of Phragmites australis and Bolboschoenus planiculmis. Sci. Total Environ. 2018, 644, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.; Yu, J.B.; Hou, A.X.; Han, G.X.; Wang, G.M.; Qu, F.Z.; Xia, J.B.; Wang, X.H. The ecological adaptability of Phragmites australis to interactive effects of water level and salt stress in the Yellow River Delta. Aquat. Ecol. 2017, 51, 107–116. [Google Scholar] [CrossRef]
- Ryu, H.; Cho, Y.G. Plant hormones in salt stress tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Yu, Z.P.; Duan, X.B.; Luo, L.; Dai, S.J.; Ding, Z.J.; Xia, G.M. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Song, H.J.; Guo, X.; Yang, J.C.; Liu, L.L.; Li, M.Y.; Wang, J.F.; Guo, W.H. Phenotypic plasticity variations in Phragmites australis under different plant-plant interactions influenced by salinity. J. Plant Ecol. 2024, 17, rtae035. [Google Scholar] [CrossRef]
- Kobayashi, K. Sexual reproduction and diversity: Connection between sexual selection and biological communities via population dynamics. Popul. Ecol. 2019, 61, 135–140. [Google Scholar] [CrossRef]
- Hualpa-Ramirez, E.; Carrasco-Lozano, E.C.; Madrid-Espinoza, J.; Tejos, R.; Ruiz-Lara, S.; Stange, C.; Norambuena, L. Stress salinity in plants: New strategies to cope with in the foreseeable scenario. Plant Physiol. Biochem. 2024, 208, 108507. [Google Scholar] [CrossRef]
- Pagter, M.; Bragato, C.; Brix, H. Tolerance and physiological responses of Phragmites australis to water deficit. Aquat. Bot. 2005, 81, 285–299. [Google Scholar] [CrossRef]
- Yang, Z.F.; Xie, T.; Liu, Q. Physiological responses of Phragmites australis to the combined effects of water and salinity stress. Ecohydrology 2014, 7, 420–426. [Google Scholar] [CrossRef]
- Guo, H.Y.; Gao, F.L.; Pang, J.L.; Wang, H.H.; Wang, H.D.; Wang, Y.H.; Whitt, A.A.; Ma, C.C. Plant-plant interactions of Phragmites australis and Suaeda salsa as mediated by combined influences of salinity and tidal level changes. Plant Soil. 2022, 474, 141–161. [Google Scholar] [CrossRef]
- Gao, F.L.; Wang, Y.H.; Whitt, A.A.; Wang, H.D.; Ma, C.C.; Guo, H.Y. Belowground responses of Phragmites australis and Suaeda salsa to salinity and water depth changes. Pak. J. Bot. 2018, 50, 853–861. [Google Scholar]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shao, R.Y.; Zhu, P.C.; Wang, R.Q. The reproductive strategy of the clonal helophyte Leersia oryzoides (L.) Swartz. in response to variable submergence conditions and different harvest times. Evol. Ecol. 2021, 35, 27–40. [Google Scholar] [CrossRef]
- Yu, X.; Xu, B.Y.; Yao, R.J.; Wei, J.H.; Tu, T.B.; Chen, Z. Temporal dynamics of soil salinization due to vertical and lateral saltwater intrusion at an onshore aquaculture farm. Agric. Water Manag. 2024, 306, 109179. [Google Scholar] [CrossRef]
- Kurepa, J.; Smalle, J.A. Plant hormone modularity and the survival-reproduction trade-off. Biology 2023, 12, 1143. [Google Scholar] [CrossRef]
- Pagnussat, G.C.; Gomez-Casati, D.F. Plant development and reproduction in a changing environment. J. Exp. Bot. 2024, 75, 4167–4170. [Google Scholar] [CrossRef]
- Elsey-Quirk, T.; Lynn, A.; Jacobs, M.D.; Diaz, R.; Cronin, J.T.; Wang, L.; Huang, H.; Justic, D. Vegetation dieback in the Mississippi River Delta triggered by acute drought and chronic relative sea-level rise. Nat. Commun. 2024, 15, 3518. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Jiang, L.M.; Chen, Y.D.; Tian, X.M.; Lv, G.H. Abiotic stress-by-competition interactions drive hormone and nutrient changes to regulate Suaeda salsa growth. Glob. Ecol. Conserv. 2021, 31, e01845. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | df | F | p |
|---|---|---|---|
| Number of flowering shoots of P. australis | |||
| Salinity | 1, 24 | 57.791 | <0.001 *** |
| Tidal level | 2, 24 | 135.075 | <0.001 *** |
| Salinity × Tidal level | 2, 24 | 3.970 | 0.032 * |
| Proportion of flowering shoots of P. australis | |||
| Salinity | 1, 24 | 29.574 | <0.001 *** |
| Tidal level | 2, 24 | 17.253 | <0.001 *** |
| Salinity × Tidal level | 2, 24 | 0.110 | 0.896 |
| Inflorescence biomass of P. australis | |||
| Salinity | 1, 24 | 71.162 | <0.001 *** |
| Tidal level | 2, 24 | 28.068 | <0.001 *** |
| Salinity × Tidal level | 2, 24 | 3.601 | 0.043 * |
| Number of rhizome buds of P. australis | |||
| Salinity | 1, 24 | 95.656 | <0.001 *** |
| Tidal level | 2, 24 | 37.663 | <0.001 *** |
| Salinity × Tidal level | 2, 24 | 5.602 | 0.011 * |
| Rhizome biomass of P. australis | |||
| Salinity | 1, 24 | 46.182 | <0.001 *** |
| Tidal level | 2, 24 | 9.695 | <0.001 *** |
| Salinity × Tidal level | 2, 24 | 7.238 | 0.003 ** |
| Ratio of sexual to clonal biomass allocation of P. australis | |||
| Salinity | 1, 24 | 30.329 | <0.001 *** |
| Tidal level | 2, 24 | 95.603 | <0.001 *** |
| Salinity × Tidal level | 2, 24 | 3.728 | 0.039 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Lyu, X.; Tian, X.; Guo, C.; Li, Y.; Yang, C.; Li, N.; Guo, H. Shift in the Reproductive Strategies of Phragmites australis Under Combined Influences of Salinity and Tidal Level Changes. Agronomy 2025, 15, 1587. https://doi.org/10.3390/agronomy15071587
Wang Y, Lyu X, Tian X, Guo C, Li Y, Yang C, Li N, Guo H. Shift in the Reproductive Strategies of Phragmites australis Under Combined Influences of Salinity and Tidal Level Changes. Agronomy. 2025; 15(7):1587. https://doi.org/10.3390/agronomy15071587
Chicago/Turabian StyleWang, Yinhua, Xin Lyu, Xinyi Tian, Changcheng Guo, Yifan Li, Chen Yang, Ningning Li, and Hongyu Guo. 2025. "Shift in the Reproductive Strategies of Phragmites australis Under Combined Influences of Salinity and Tidal Level Changes" Agronomy 15, no. 7: 1587. https://doi.org/10.3390/agronomy15071587
APA StyleWang, Y., Lyu, X., Tian, X., Guo, C., Li, Y., Yang, C., Li, N., & Guo, H. (2025). Shift in the Reproductive Strategies of Phragmites australis Under Combined Influences of Salinity and Tidal Level Changes. Agronomy, 15(7), 1587. https://doi.org/10.3390/agronomy15071587