

Variation in Root Traits and Root-Endophyte Interactions in Primary Synthetic Wheat Derived from Aegilops tauschii Collected from Diverse Soil Types

 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experiment Setup
2.3. Harvest and Data Collection
2.3.1. Shoot Traits
2.3.2. Root Traits
2.3.3. Root Biotic Traits
2.4. Statistical Analysis
3. Results
3.1. Variation in Traits Among Wheat PS Lines
3.1.1. Root Traits
3.1.2. Shoot Traits
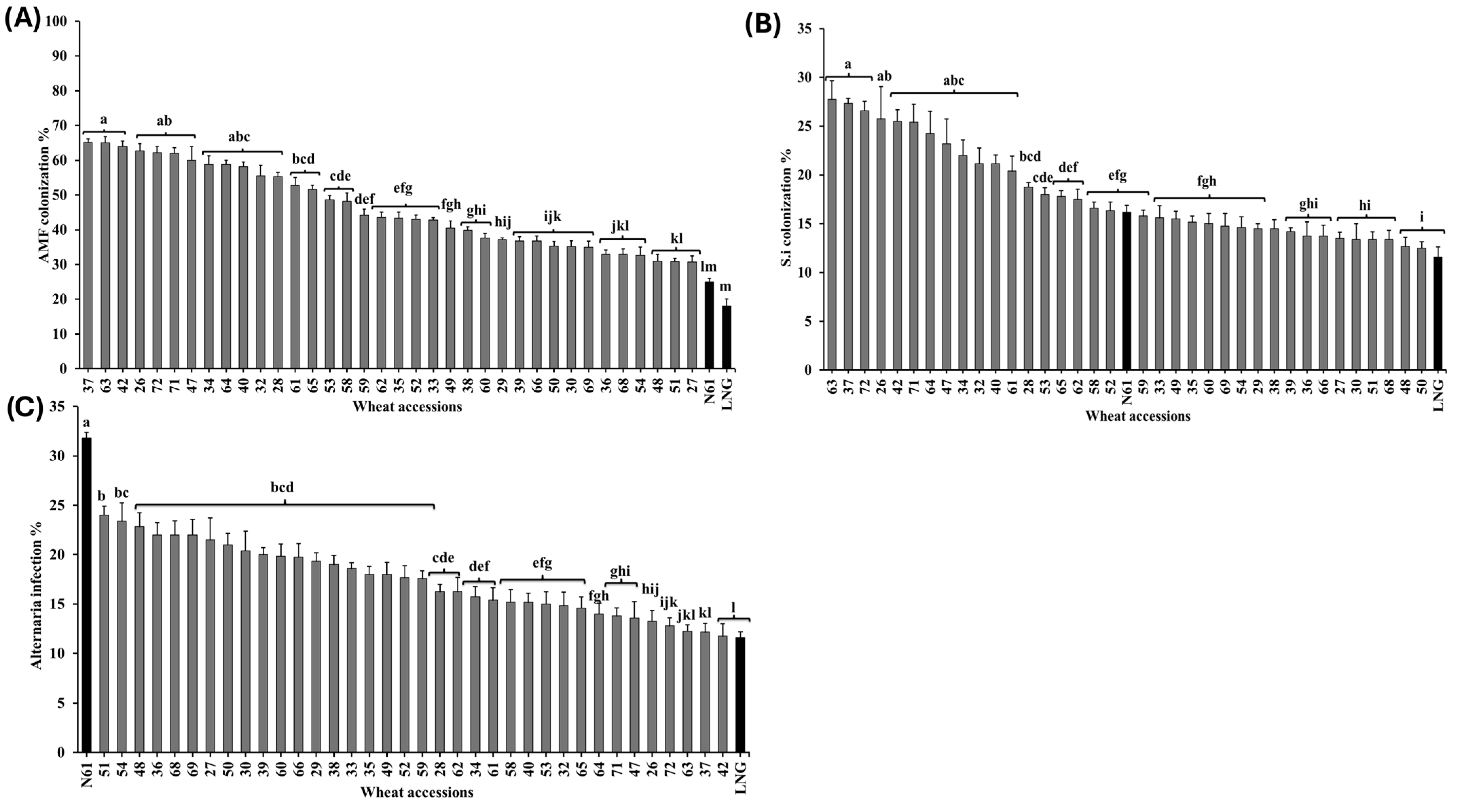
3.1.3. Biotic Traits
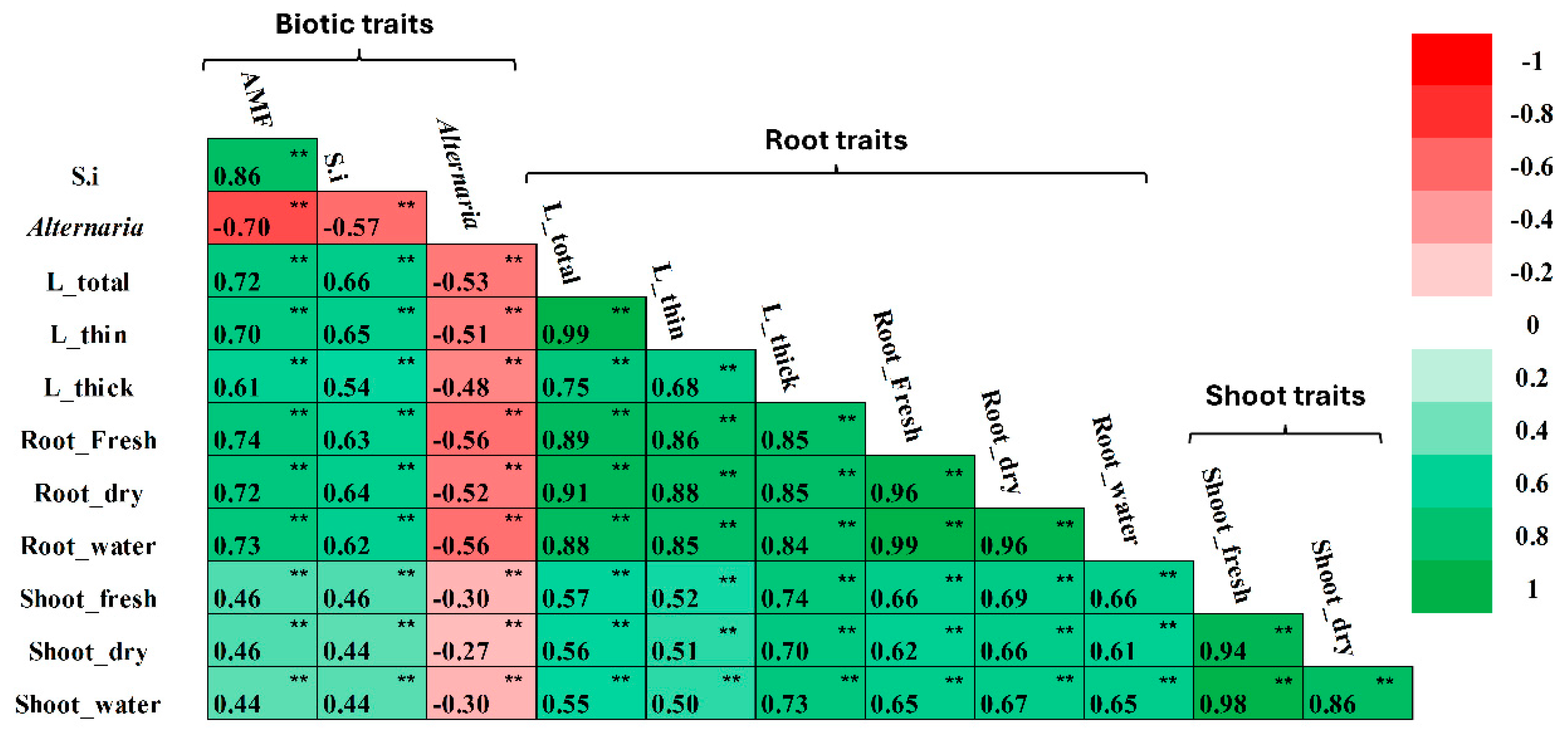
3.1.4. Association Among Wheat Accession Traits
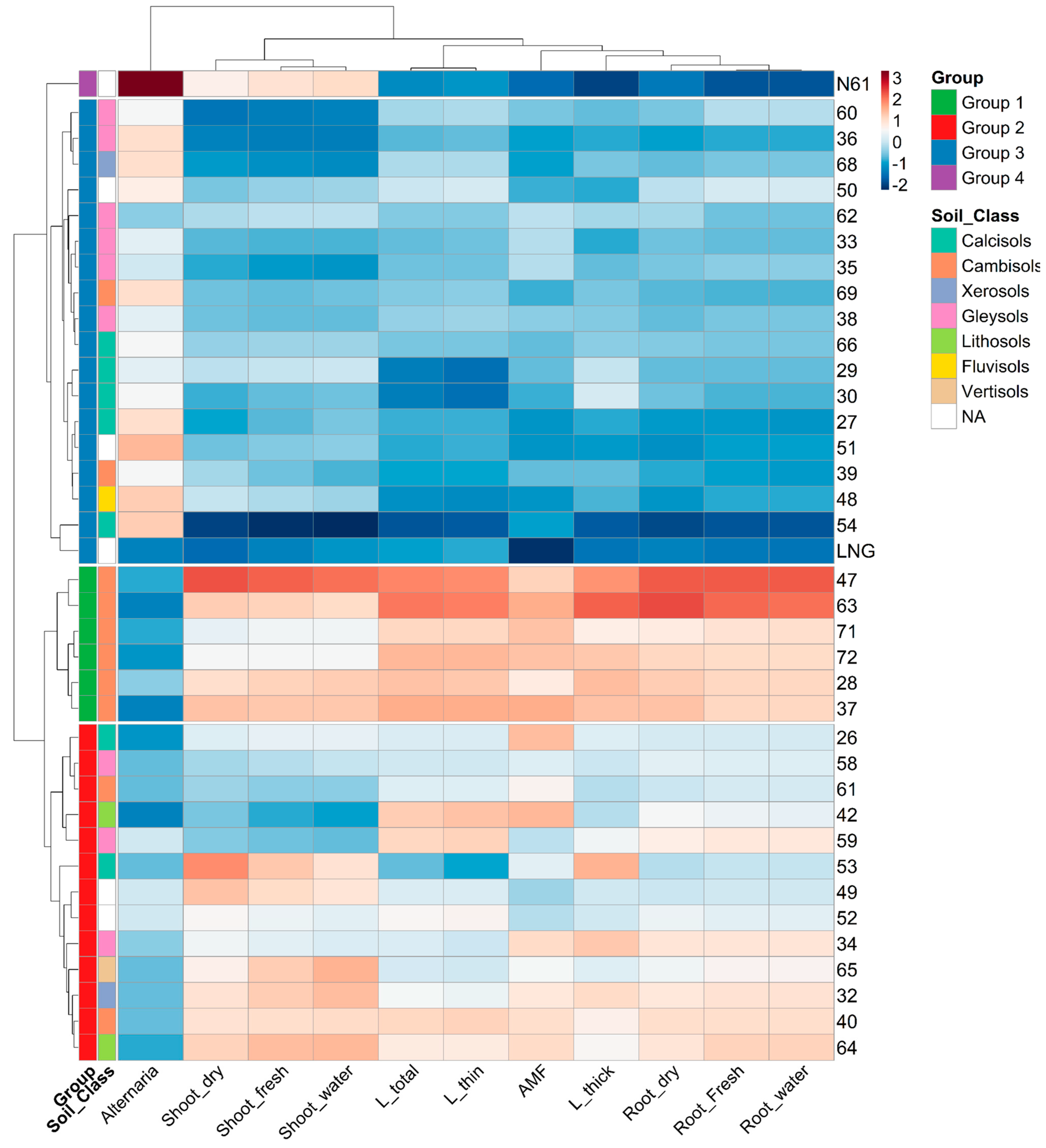
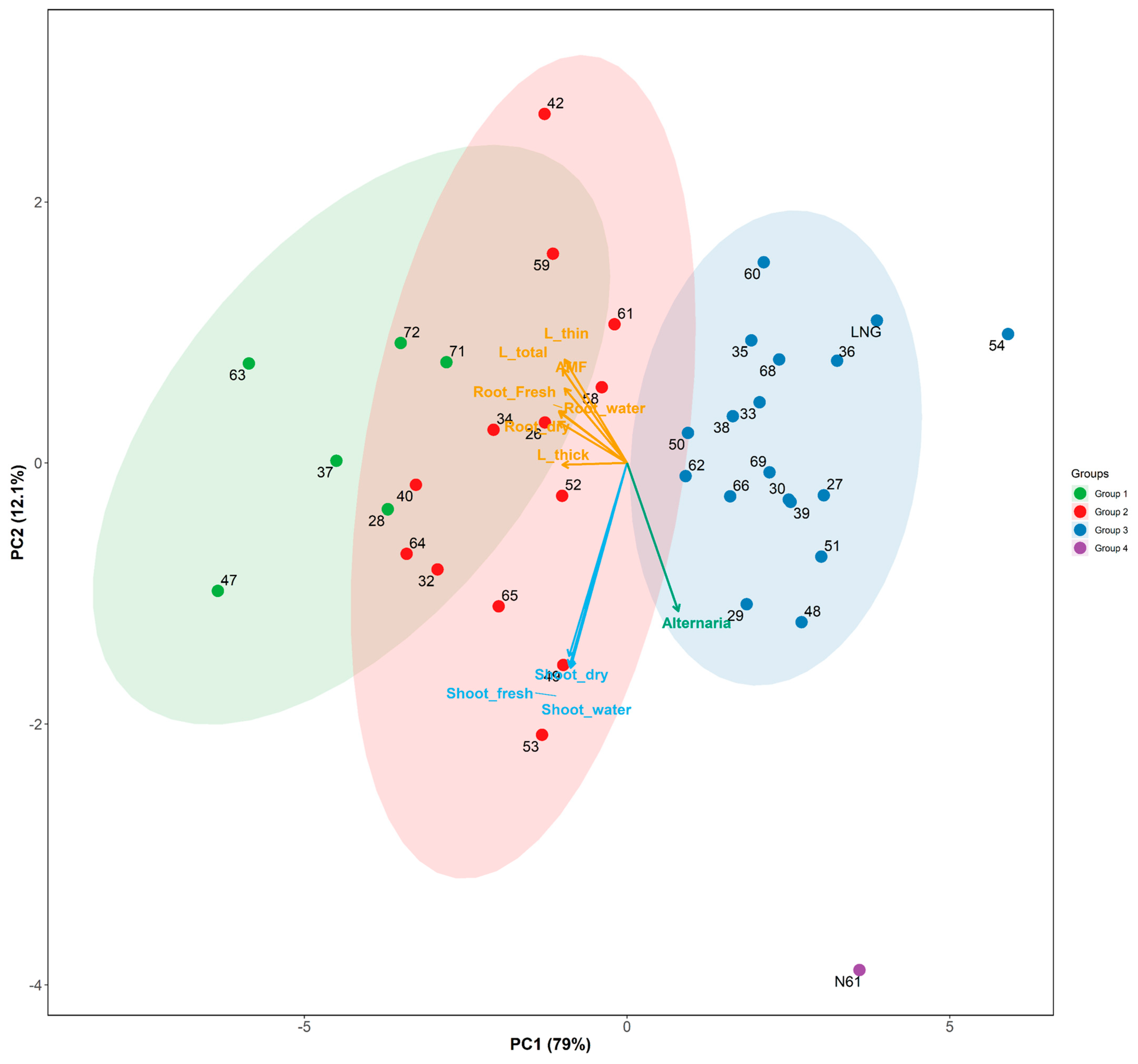
3.2. Wheat Accession Grouping and Traits Association
4. Discussion
- Biotic traits variability among PS lines: AMF, S. indica, and Alternaria resistance
- Root-biotic trait associations
- Soil classification affects wheat root traits grouping
- Impact of domesticated LNG on PS lines
- Limitations and future directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.W.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K.; Blanco, G.; et al. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2023. [Google Scholar]
- Constas, M.A.; d’Errico, M.; Hoddinott, J.F.; Pietrelli, R. Resilient Food Systems—A Proposed Analytical Strategy for Empirical Applications. Available online: https://ageconsearch.umn.edu/record/319840 (accessed on 28 March 2025).
- Reif, J.C.; Zhang, P.; Dreisigacker, S.; Warburton, M.L.; van Ginkel, M.; Hoisington, D.; Bohn, M.; Melchinger, A.E. Wheat Genetic Diversity Trends during Domestication and Breeding. TAG Theor. Appl. Genet. Theor. Angew. Genet. 2005, 110, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Maeght, J.-L.; Rewald, B.; Pierret, A. How to Study Deep Roots—And Why It Matters. Front. Plant Sci. 2013, 4, 299. [Google Scholar] [CrossRef] [PubMed]
- Pierret, A.; Maeght, J.-L.; Clément, C.; Montoroi, J.-P.; Hartmann, C.; Gonkhamdee, S. Understanding Deep Roots and Their Functions in Ecosystems: An Advocacy for More Unconventional Research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef]
- Ober, E.S.; Alahmad, S.; Cockram, J.; Forestan, C.; Hickey, L.T.; Kant, J.; Maccaferri, M.; Marr, E.; Milner, M.; Pinto, F.; et al. Wheat Root Systems as a Breeding Target for Climate Resilience. Theor. Appl. Genet. 2021, 134, 1645–1662. [Google Scholar] [CrossRef]
- Jaster-Keller, J.; Müller, M.E.H.; El-Khatib, A.H.; Lorenz, N.; Bahlmann, A.; Mülow-Stollin, U.; Bunzel, M.; Scheibenzuber, S.; Rychlik, M.; von der Waydbrink, G.; et al. Root Uptake and Metabolization of Alternaria Toxins by Winter Wheat Plants Using a Hydroponic System. Mycotoxin Res. 2023, 39, 109–126. [Google Scholar] [CrossRef]
- Qiao, X.; Sun, T.; Lei, J.; Xiao, L.; Xue, L.; Zhang, H.; Jia, J.; Bei, S. Arbuscular Mycorrhizal Fungi Contribute to Wheat Yield in an Agroforestry System with Different Tree Ages. Front. Microbiol. 2022, 13, 1024128. [Google Scholar] [CrossRef]
- Crespo-Herrera, L.A.; Govindan, V.; Stangoulis, J.; Hao, Y.; Singh, R.P. QTL Mapping of Grain Zn and Fe Concentrations in Two Hexaploid Wheat RIL Populations with Ample Transgressive Segregation. Front. Plant Sci. 2017, 8, 1800. [Google Scholar] [CrossRef]
- Elbashir, A.A.E.; Gorafi, Y.S.A.; Tahir, I.S.A.; Kim, J.-S.; Tsujimoto, H. Wheat Multiple Synthetic Derivatives: A New Source for Heat Stress Tolerance Adaptive Traits. Breed. Sci. 2017, 67, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Itam, M.O.; Mega, R.; Gorafi, Y.S.A.; Yamasaki, Y.; Tahir, I.S.A.; Akashi, K.; Tsujimoto, H. Genomic Analysis for Heat and Combined Heat–Drought Resilience in Bread Wheat under Field Conditions. Theor. Appl. Genet. 2022, 135, 337–350. [Google Scholar] [CrossRef]
- Mahjoob, M.M.M.; Chen, T.-S.; Gorafi, Y.S.A.; Yamasaki, Y.; Kamal, N.M.; Abdelrahman, M.; Iwata, H.; Matsuoka, Y.; Tahir, I.S.A.; Tsujimoto, H. Traits to Differentiate Lineages and Subspecies of Aegilops Tauschii, the D Genome Progenitor Species of Bread Wheat. Diversity 2021, 13, 217. [Google Scholar] [CrossRef]
- Jones, A.; Caon, L.; Yigini, Y.; Verbeke, I.; Konyushkova, M.; Vargas, R.; Viatkin, K.; Michéli, E. Soil Atlas of Asia; European Commission, Ed.; Publications Office: Luxembourg, 2024; ISBN 978-92-68-03116-2. [Google Scholar]
- Mauer, O.; Palátová, E. Mountain Ash (Sorbus aucuparia L.) Root System Morphogenesis. J. For. Sci. 2002, 48, 342–350. [Google Scholar] [CrossRef]
- Wang, H.; Hu, Z.; Huang, K.; Han, Y.; Zhao, A.; Han, H.; Song, L.; Fan, C.; Li, R.; Xin, M.; et al. Three Genomes Differentially Contribute to the Seedling Lateral Root Number in Allohexaploid Wheat: Evidence from Phenotype Evolution and Gene Expression. Plant J. 2018, 95, 976–987. [Google Scholar] [CrossRef]
- Mohammedali, A.K.H.; Gorafi, Y.S.A.; Kamal, N.M.; Tahir, I.S.A.; Tsujimoto, H.; Taniguchi, T. Wheat Root Traits and Endophytic Fungal Association Differ with Ploidy. Front. Agron. 2025; under review. [Google Scholar]
- Maccaferri, M.; El-Feki, W.; Nazemi, G.; Salvi, S.; Canè, M.A.; Colalongo, M.C.; Stefanelli, S.; Tuberosa, R. Prioritizing Quantitative Trait Loci for Root System Architecture in Tetraploid Wheat. J. Exp. Bot. 2016, 67, 1161–1178. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Harnessing Root Architecture to Address Global Challenges. Plant J. 2022, 109, 415–431. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Takumi, S.; Kawahara, T. Natural Variation for Fertile Triploid F1 Hybrid Formation in Allohexaploid Wheat Speciation. Theor. Appl. Genet. 2007, 115, 509–518. [Google Scholar] [CrossRef]
- FAO. Fertilizer Use by Crop in the Islamic Republic of Iran. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/a59738f1-92fc-4bac-bbd9-00e6eadb0483/content (accessed on 30 April 2025).
- Martín, M.; Rubio, A.; Remesal, E.; Cano, C.; Bago, A. Application of the Ultimate Arbuscular Mycorrhizal Inoculant MYCOGEL® in Japan: Results and Prospects. J. Integr. Field Sci. 2018, 15, 31–40. Available online: http://hdl.handle.net/10097/00124000 (accessed on 4 June 2025).
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- McGONIGLE, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A New Method Which Gives an Objective Measure of Colonization of Roots by Vesicular—Arbuscular Mycorrhizal Fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Smith, F.A. Plant Roots. Growth, Activity and Interaction with Soils. Ann. Bot. 2007, 100, 151–152. [Google Scholar] [CrossRef]
- Lynch, J.P. Root Phenotypes for Improved Nutrient Capture: An Underexploited Opportunity for Global Agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, A.; Ali, S.; Javed, M.A.; Iqbal, R.; Khan, M.N.; Çiğ, F.; Sabagh, A.E.; Abujamel, T.; Harakeh, S.; Ercisli, S.; et al. Breeding for Water-Use Efficiency in Wheat: Progress, Challenges and Prospects. Mol. Biol. Rep. 2024, 51, 429. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Long, L.; Sun, X.; Parsons, D.; Zhou, Z. Responsive Root Traits and Mitigating Strategies for Wheat Production under Single or Combined Abiotic Stress. Eur. J. Agron. 2025, 162, 127393. [Google Scholar] [CrossRef]
- Cheng, S.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Elucidating the Mechanisms Underlying Enhanced Drought Tolerance in Plants Mediated by Arbuscular Mycorrhizal Fungi. Front. Microbiol. 2021, 12, 809473. [Google Scholar] [CrossRef]
- Hosseini, F.; Mosaddeghi, M.R.; Zarebanadkouki, M. Enhancing Rhizosphere Soil Water Retention in Wheat through Colonization with Endophytic Fungus Serendipita indica. Rhizosphere 2023, 26, 100709. [Google Scholar] [CrossRef]
- Duan, H.-X.; Luo, C.-L.; Zhu, Y.; Zhao, L.; Wang, J.; Wang, W.; Xiong, Y.-C. Arbuscular Mycorrhizal Fungus Activates Wheat Physiology for Higher Reproductive Allocation under Drought Stress in Primitive and Modern Wheat. Eur. J. Agron. 2024, 161, 127376. [Google Scholar] [CrossRef]
- Tkacz, A.; Pini, F.; Turner, T.R.; Bestion, E.; Simmonds, J.; Howell, P.; Greenland, A.; Cheema, J.; Emms, D.M.; Uauy, C.; et al. Agricultural Selection of Wheat Has Been Shaped by Plant-Microbe Interactions. Front. Microbiol. 2020, 11, 132. [Google Scholar] [CrossRef]
- Lattacher, A.; Le Gall, S.; Rothfuss, Y.; Gao, C.; Harings, M.; Pagel, H.; Giraud, M.; Alahmad, S.; Hickey, L.T.; Kandeler, E.; et al. Rooting for Microbes: Impact of Root Architecture on the Microbial Community and Function in Top- and Subsoil. Plant Soil 2025, 1–19. [Google Scholar] [CrossRef]
- Klimek-Kopyra, A.; Kulig, B.; Głąb, T.; Zając, T.; Skowera, B.; Kopcińska, J. Effect of plant intercropping and soil type on specific root length. Rom. Agric. Res. 2015, 32, 1–10. [Google Scholar]
- Schneider, F.; Don, A. Root-Restricting Layers in German Agricultural Soils. Part I: Extent and Cause. Plant Soil 2019, 442, 433–451. [Google Scholar] [CrossRef]
- Labidi, S.; Calonne, M.; Ben Jeddi, F.; Debiane, D.; Rezgui, S.; Laruelle, F.; Tisserant, B.; Grandmougin-Ferjani, A.; Lounès-Hadj Sahraoui, A. Calcareous Impact on Arbuscular Mycorrhizal Fungus Development and on Lipid Peroxidation in Monoxenic Roots. Phytochemistry 2011, 72, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Sediqui, N.; Amin, M.W.; Dawlatzai, N.; Gulab, G.; Poyesh, D.S.; Terada, N.; Sanada, A.; Kamata, A.; Koshio, K. Elucidation of Shoot and Root Growth, Physiological Responses, and Quality Traits of Tomato (Solanum lycopersicon L.) Exposed to Elevated Calcium Carbonate Concentrations. Horticulturae 2024, 10, 573. [Google Scholar] [CrossRef]
- Bektas, H.; Hohn, C.E.; Waines, J.G. Characteristics of the Root System in the Diploid Genome Donors of Hexaploid Wheat (Triticum aestivum L.). Genet. Resour. Crop Evol. 2017, 64, 1641–1650. [Google Scholar] [CrossRef]
- Li, A.; Liu, D.; Yang, W.; Kishii, M.; Mao, L. Synthetic Hexaploid Wheat: Yesterday, Today, and Tomorrow. Engineering 2018, 4, 552–558. [Google Scholar] [CrossRef]
- Gorafi, Y.S.A.; Kim, J.-S.; Elbashir, A.A.E.; Tsujimoto, H. A Population of Wheat Multiple Synthetic Derivatives: An Effective Platform to Explore, Harness and Utilize Genetic Diversity of Aegilops tauschii for Wheat Improvement. Theor. Appl. Genet. 2018, 131, 1615–1626. [Google Scholar] [CrossRef]
- Ma, J.; Zhao, D.; Tang, X.; Yuan, M.; Zhang, D.; Xu, M.; Duan, Y.; Ren, H.; Zeng, Q.; Wu, J.; et al. Genome-Wide Association Study on Root System Architecture and Identification of Candidate Genes in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2022, 23, 1843. [Google Scholar] [CrossRef]
- Xu, F.; Chen, S.; Yang, X.; Zhou, S.; Wang, J.; Zhang, Z.; Huang, Y.; Song, M.; Zhang, J.; Zhan, K.; et al. Genome-Wide Association Study on Root Traits Under Different Growing Environments in Wheat (Triticum aestivum L.). Front. Genet. 2021, 12, 646712. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammedali, A.K.H.; Kamal, N.M.; Gorafi, Y.S.A.; Tahir, I.S.A.; Tsujimoto, H.; Taniguchi, T. Variation in Root Traits and Root-Endophyte Interactions in Primary Synthetic Wheat Derived from Aegilops tauschii Collected from Diverse Soil Types. Agronomy 2025, 15, 1443. https://doi.org/10.3390/agronomy15061443
Mohammedali AKH, Kamal NM, Gorafi YSA, Tahir ISA, Tsujimoto H, Taniguchi T. Variation in Root Traits and Root-Endophyte Interactions in Primary Synthetic Wheat Derived from Aegilops tauschii Collected from Diverse Soil Types. Agronomy. 2025; 15(6):1443. https://doi.org/10.3390/agronomy15061443
Chicago/Turabian StyleMohammedali, Ahmed Khaled Hassan, Nasrein Mohamed Kamal, Yasir Serag Alnor Gorafi, Izzat Sidahmed Ali Tahir, Hisashi Tsujimoto, and Takeshi Taniguchi. 2025. "Variation in Root Traits and Root-Endophyte Interactions in Primary Synthetic Wheat Derived from Aegilops tauschii Collected from Diverse Soil Types" Agronomy 15, no. 6: 1443. https://doi.org/10.3390/agronomy15061443
APA StyleMohammedali, A. K. H., Kamal, N. M., Gorafi, Y. S. A., Tahir, I. S. A., Tsujimoto, H., & Taniguchi, T. (2025). Variation in Root Traits and Root-Endophyte Interactions in Primary Synthetic Wheat Derived from Aegilops tauschii Collected from Diverse Soil Types. Agronomy, 15(6), 1443. https://doi.org/10.3390/agronomy15061443