
Transcriptome Analysis Reveals the Molecular Mechanisms Through Which Exogenous Methyl Jasmonate Enhances Mercury Stress Tolerance in Maize Seedlings

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Mercury Stress Treatment
2.2. Determination of Physiological Traits
2.3. RNA Isolation and Sequencing
2.4. Assembly Standardization and Quality Assessment
2.5. Differentially Expressed Gene (DEG) Identification
2.6. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
2.7. Quantitative PCR (qRT-PCR) Verification of RNA-Seq Data
2.8. Statistical Analysis
3. Results
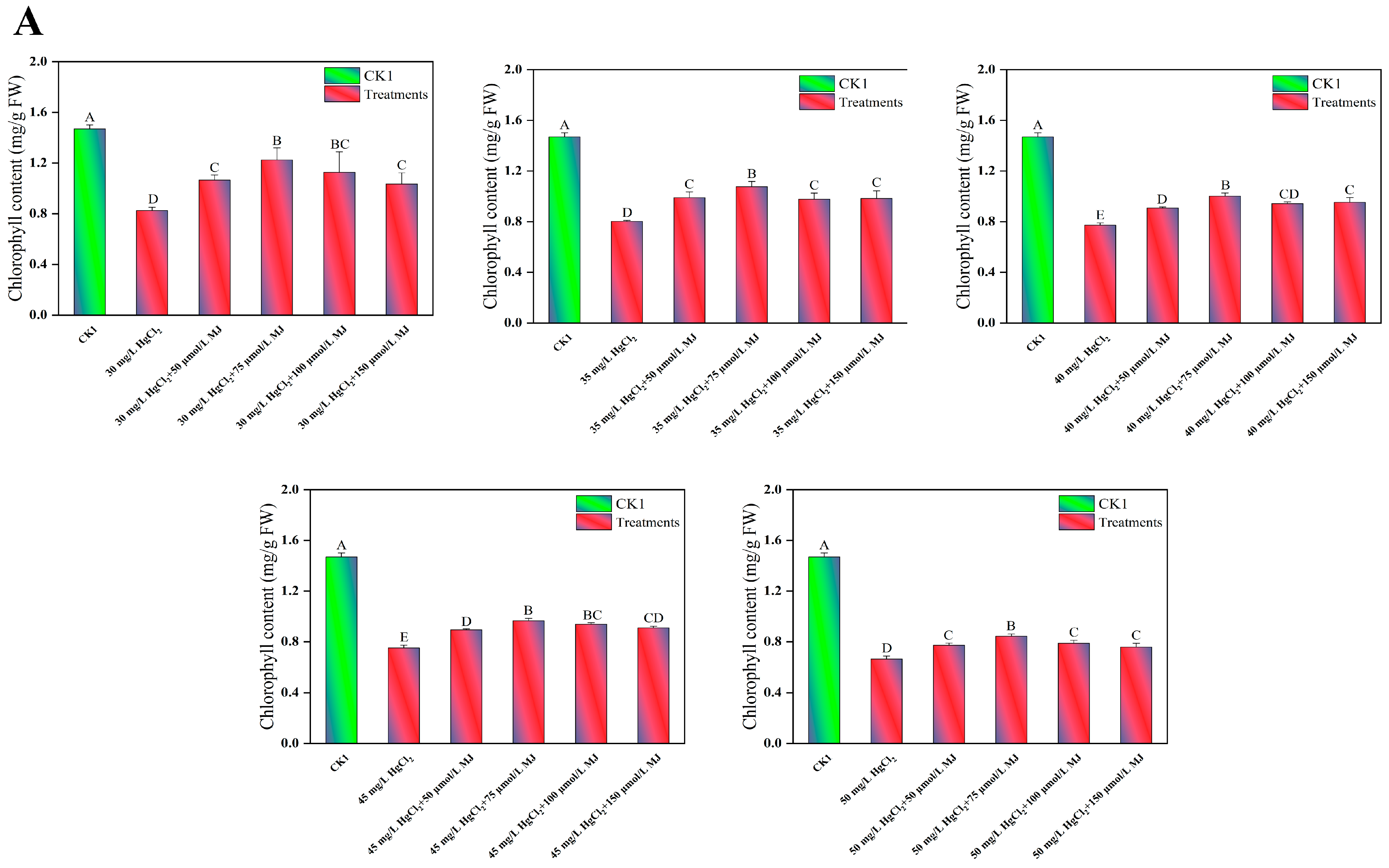
3.1. Effects of MJ Application on the Morphological and Physiological Traits of Maize Seedlings Under Hg Stress
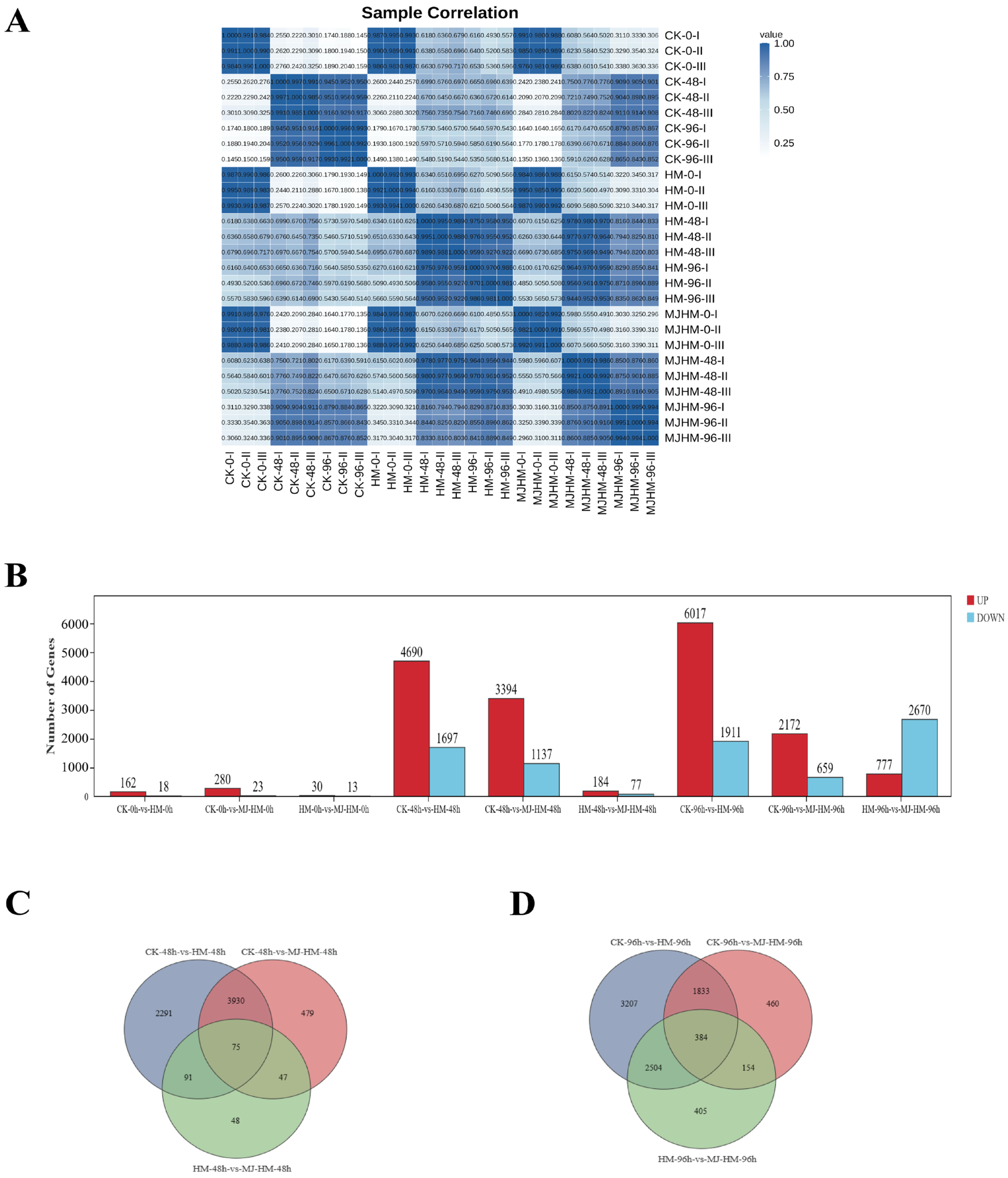
3.2. RNA Sequencing, Assembly, and Annotation
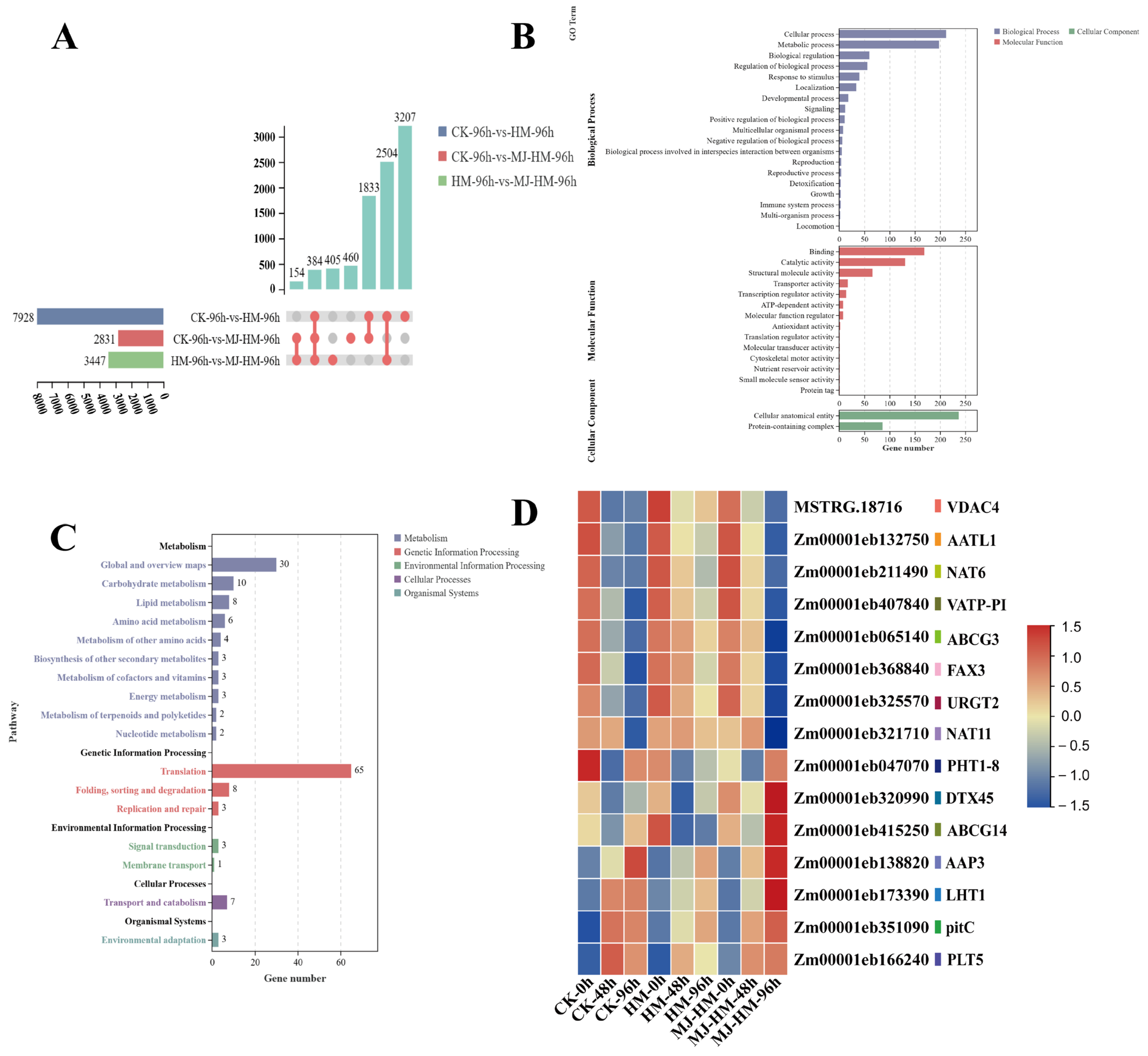
3.3. Analysis of Differentially Expressed Genes in Maize Seedlings Under Different Treatments
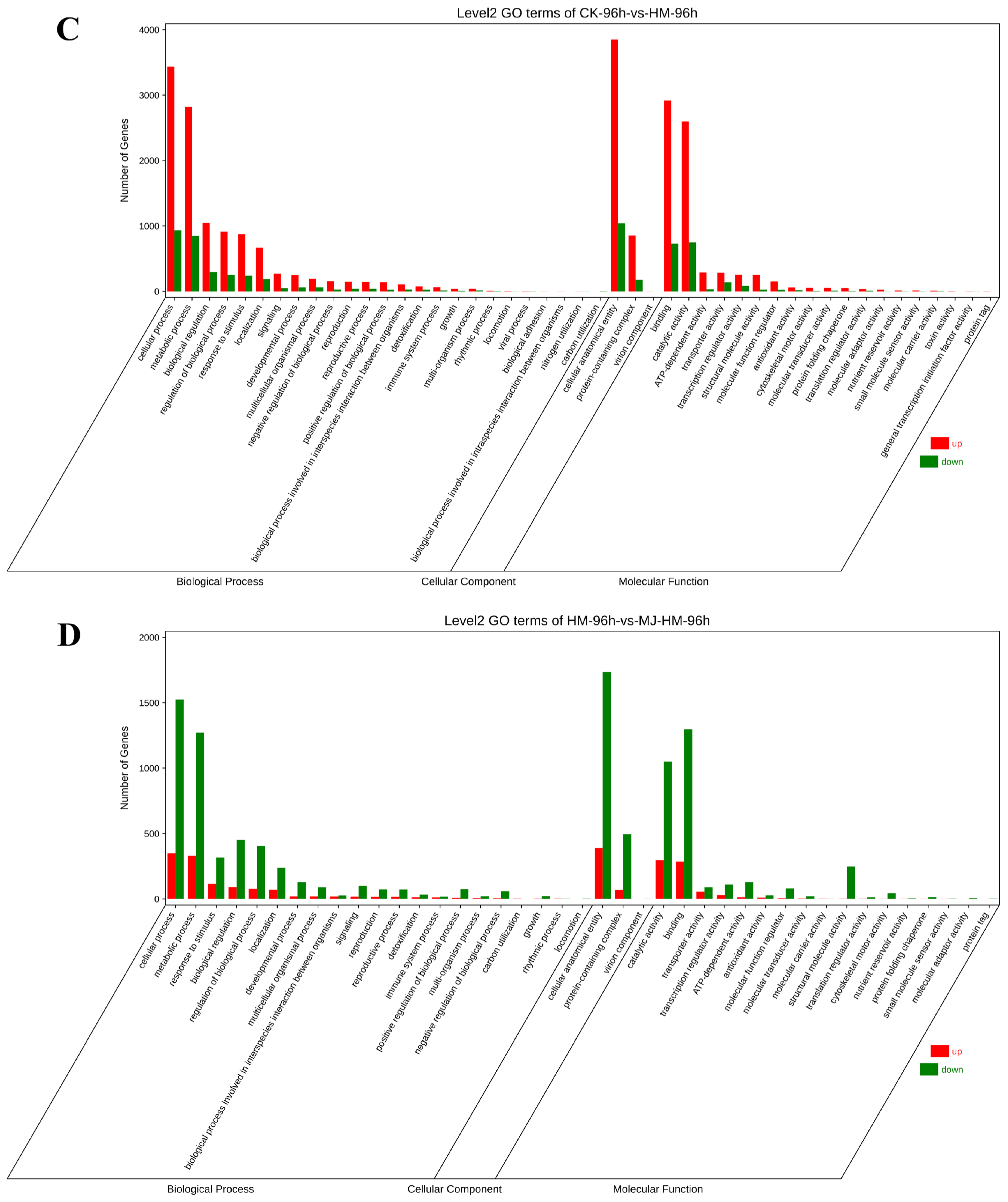
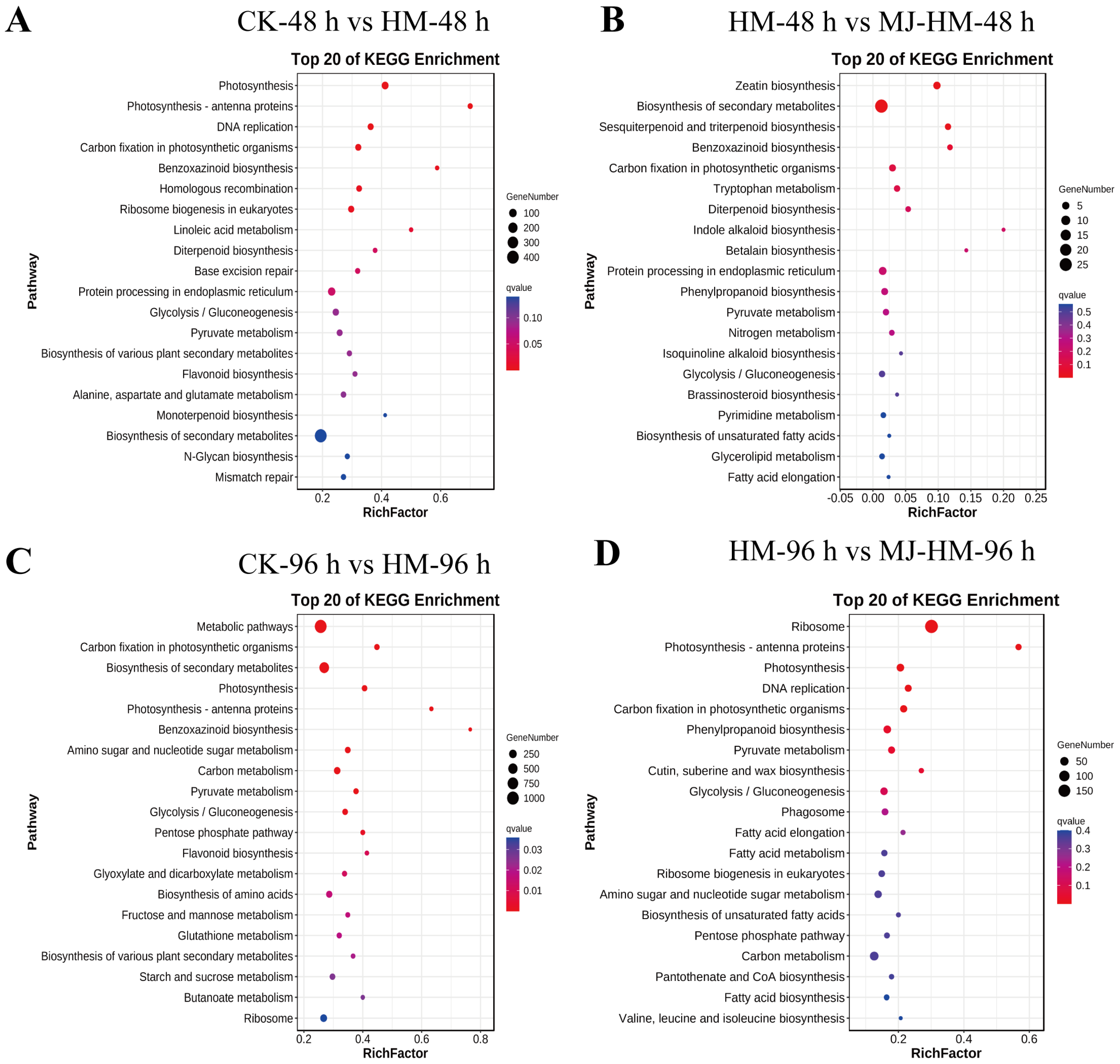
3.4. Gene Ontology and Kyoto Encyclopedia of Genes Assessment of DEGs in Maize Seedlings Under Different Treatments
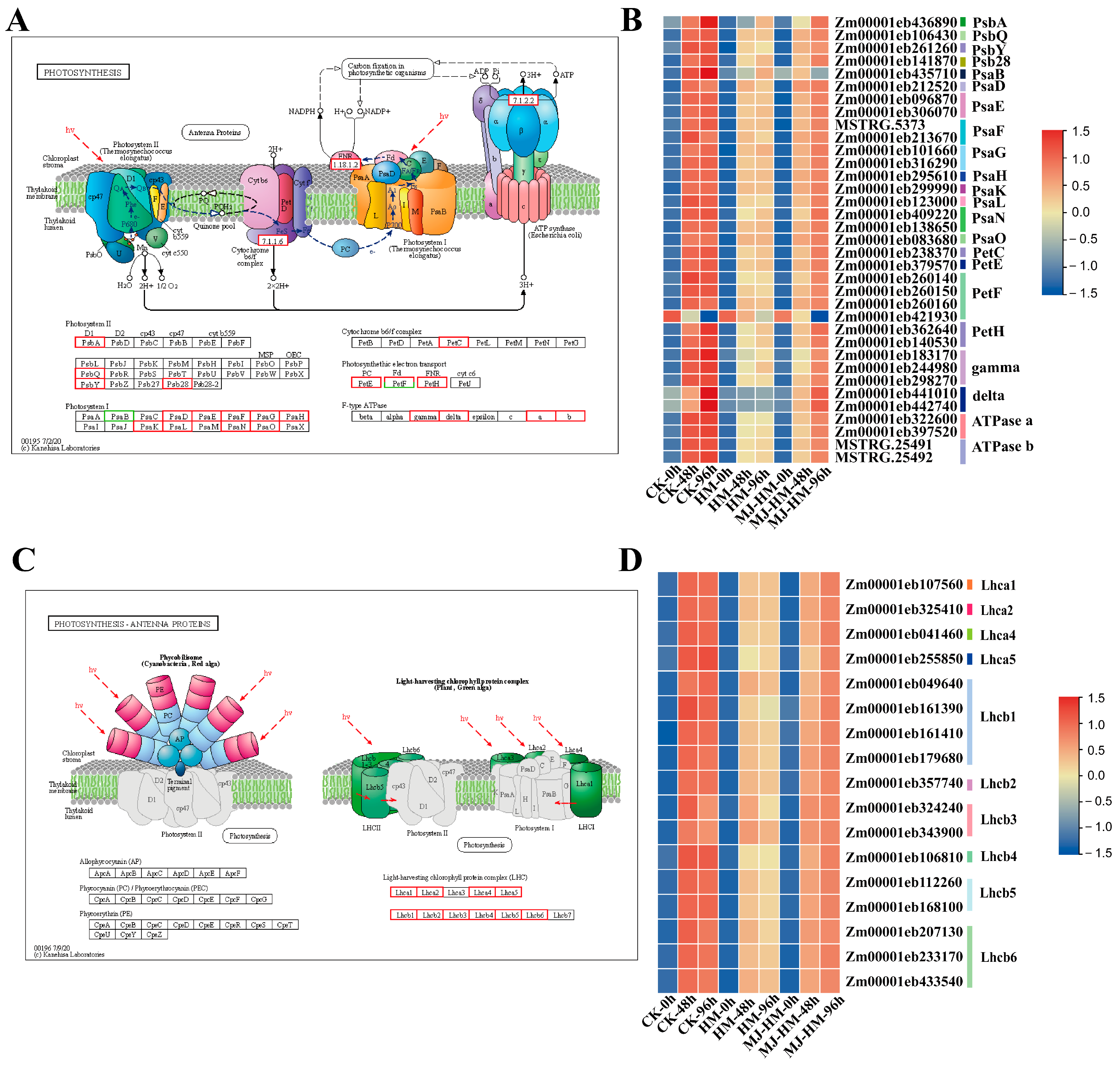
3.5. Effects of MJ Application on the Expression of Photosynthesis-Related Genes in Maize Seedlings Under Hg Stress
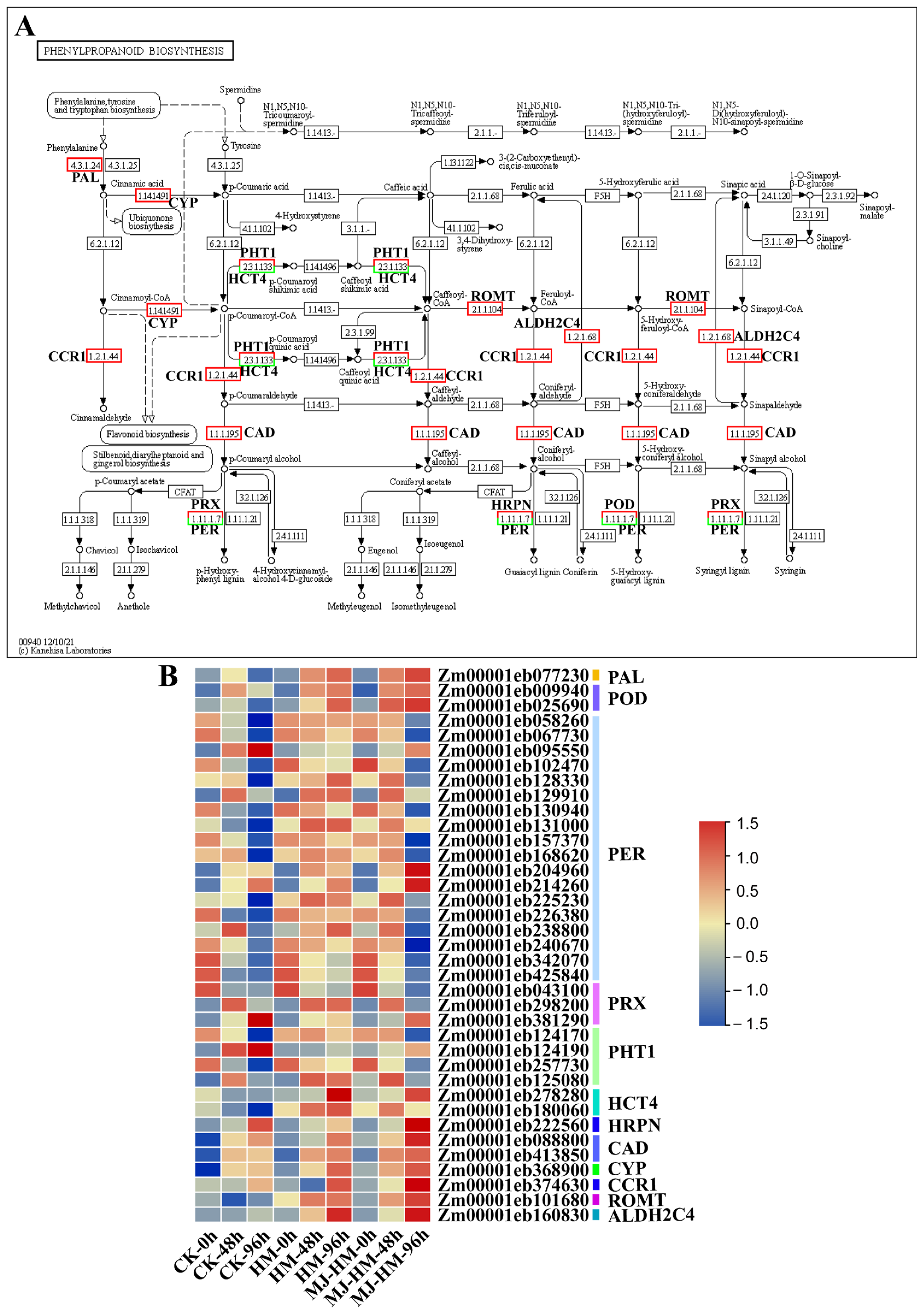
3.6. Effects of MJ Application on the Expression of Phenylpropanoid Biosynthesis-Related Genes in Maize Seedlings Under Hg Stress
3.7. Effects of MJ Application on the Expression of Proline Metabolism-Related Genes in Maize Seedlings Under Hg Stress
3.8. Effects of MJ Application on the Expression of Transport Genes in Maize Seedlings Under Hg Stress
3.9. Verification of RNA-Seq Results by qRT-PCR
4. Discussion
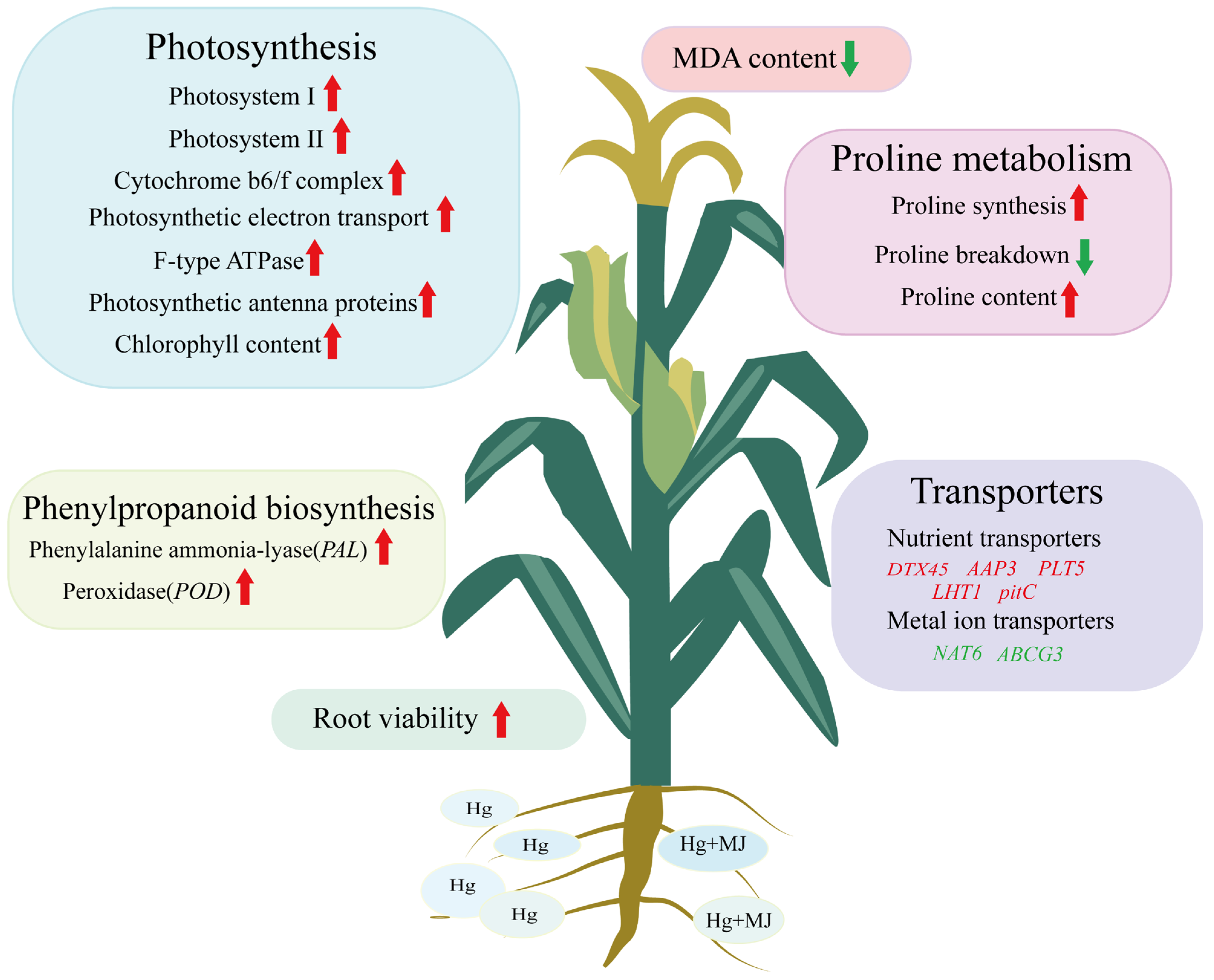
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and lonomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed]
- Sobariu, D.L.; Fertu, D.I.T.; Diaconu, M.; Pavel, L.V.; Hlihor, R.M.; Dragoi, E.N.; Curteanu, S.; Lenz, M.; Corvinid, P.F.X.; Gavrilescu, M. Rhizobacteria and plant symbiosis in heavy metal uptake and its implications for soil bioremediation. New Biotechnol. 2017, 39, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Tan, H.; Wang, M.; Jiang, T.; Wei, H.; Xu, W.; Jiang, Q.; Bao, H.; Ding, Y.; Wang, F.; et al. Research progress of soil microorganisms in response to heavy metals in rice. J. Agric. Food Chem. 2022, 70, 8513–8522. [Google Scholar] [CrossRef] [PubMed]
- Feki, K.; Tounsi, S.; Mrabet, M.; Mhadhbi, H.; Brini, F. Recent advances in physiological and molecular mechanisms of heavy metal accumulation in plants. Environ. Sci. Pollut. Res. 2021, 28, 64967–64986. [Google Scholar] [CrossRef]
- Niu, L.; Hao, R.; Wu, X.; Wang, W. Maize mesocotyl: Role in response to stress and deep-sowing tolerance. Plant Breed. 2020, 139, 466–473. [Google Scholar] [CrossRef]
- Chukwudi, U.P.; Kutu, F.R.; Mavengahama, S. Influence of heat stress, variations in soil type, and soil amendment on the growth of three drought-tolerant maize varieties. Agronomy 2021, 11, 1485. [Google Scholar] [CrossRef]
- Jafari, F.; Wang, B.; Wang, H.; Zou, J. Breeding maize of ideal plant architecture for high-density planting tolerance through modulating shade avoidance response and beyond. J. Integr. Plant Biol. 2024, 66, 849–864. [Google Scholar] [CrossRef]
- Huang, Y.; Yi, J.; Li, X.; Li, F. Transcriptomics and physiological analyses reveal that sulfur alleviates mercury toxicity in rice (Oryza sativa L.). J. Environ. Sci. 2024, 135, 10–25. [Google Scholar] [CrossRef]
- Goix, S.; Leveque, T.; Xiong, T.T.; Schreck, E.; Baeza-Squiban, A.; Geret, F.; Uzu, G.; Austruy, A.; Dumat, C. Environmental and health impacts of fine and ultrafine metallic particles: Assessment of threat scores. Environ. Res. 2014, 133, 185–194. [Google Scholar] [CrossRef]
- Hu, J.; Chen, G.; Xu, K.; Wang, J. Cadmium in cereal crops: Uptake and transport mechanisms and minimizing strategies. J. Agric. Food Chem. 2022, 70, 5961–5974. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Zia-ur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium stress in rice: Toxic effects, tolerance mechanisms, and management: A critical review. Environ. Sci. Pollut. Res. 2016, 23, 17859–17879. [Google Scholar] [CrossRef]
- Yuan, H.; Liu, Q.; Guo, Z.; Fu, J.; Sun, Y.; Gu, C.; Xing, B.; Dhankher, O.P. Sulfur nanoparticles improved plant growth and reduced mercury toxicity via mitigating the oxidative stress in Brassica napus L. J. Clean. Prod. 2021, 318, 128589. [Google Scholar] [CrossRef]
- Gong, Y.; Zhao, D.; Wang, Q. An overview of field-scale studies on remediation of soil contaminated with heavy metals and metalloids: Technical progress over the last decade. Water Res. 2018, 147, 440–460. [Google Scholar] [CrossRef] [PubMed]
- Roy, S. Function of MYB domain transcription factors in abiotic stress and epigenetic control of stress response in plant genome. Plant Signal. Behav. 2016, 11, e1117723. [Google Scholar] [CrossRef] [PubMed]
- Safeer, R.; Liu, G.; Yousaf, B.; Ashraf, A.; Haider, M.I.S.; Cheema, A.I.; Ijaz, S.; Rashid, A.; Sikandar, A.; Pikon, K. Insights into the biogeochemical transformation, environmental impacts and biochar-based soil decontamination of antimony. Environ. Res. 2024, 251, 118645. [Google Scholar] [CrossRef]
- Xu, D.M.; Fu, R.B.; Liu, H.Q.; Guo, X.P. Current knowledge from heavy metal pollution in Chinese smelter contaminated soils, health risk implications and associated remediation progress in recent decades: A critical review. J. Clean. Prod. 2021, 286, 124989. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, J.; Li, S.; Niinemets, U. Methyl jasmonate-induced emission of biogenic volatiles is biphasic in cucumber: A high-resolution analysis of dose dependence. J. Exp. Bot. 2017, 68, 4679–4694. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, J.; Rasulov, B.; Niinemets, U. Role of stomatal conductance in modifying the dose response of stress-volatile emissions in methyl jasmonate treated leaves of cucumber (Cucumis sativa). Int. J. Mol. Sci. 2020, 21, 1018. [Google Scholar] [CrossRef]
- Li, H.; Guo, Y.; Lan, Z.; Xu, K.; Chang, J.; Ahammed, G.J.; Ma, J.; Wei, C.; Zhang, X. Methyl jasmonate mediates melatonin-induced cold tolerance of grafted watermelon plants. Hortic. Res. 2021, 8, 57. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-responsive mechanisms in plant leaves revealed by proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef]
- Chen, J.; Yan, Z.; Li, X. Effect of methyl jasmonate on cadmium uptake and antioxidative capacity in Kandelia obovata seedlings under cadmium stress. Ecotoxicol. Environ. Saf. 2014, 104, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Anwar, T.; Qureshi, H.; Jabeen, M.; Zaman, W.; Ali, H.M. Mitigation of cadmium-induced stress in maize via synergistic application of biochar and gibberellic acid to enhance morpho-physiological and biochemical traits. BMC Plant Biol. 2024, 24, 192. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhang, W.; Chen, J.; Li, X. Methyl jasmonate alleviates cadmium toxicity in Solanum nigrum by regulating metal uptake and antioxidative capacity. Biol. Plant. 2015, 59, 373–381. [Google Scholar] [CrossRef]
- Kaushik, S.; Sharma, P.; Kaur, G.; Singh, A.K.; Al-Misned, F.A.; Shafik, H.M.; Sirhindi, G. Seed priming with methyl jasmonate mitigates copper and cadmium toxicity by modifying biochemical attributes and antioxidants in Cajanus cajan. Saudi J. Biol. Sci. 2022, 29, 721–729. [Google Scholar] [CrossRef]
- Nazir, F.; Jahan, B.; Iqbal, N.; Rajurkar, A.B.; Siddiqui, M.H.; Khan, M.I.R. Methyl jasmonate influences ethylene formation, defense systems, nutrient homeostasis and carbohydrate metabolism to alleviate arsenic-induced stress in rice (Oryza sativa). Plant Physiol. Biochem. 2023, 202, 107990. [Google Scholar] [CrossRef]
- Salavati, J.; Fallah, H.; Niknejad, Y.; Barari Tari, D. Methyl jasmonate ameliorates lead toxicity in Oryza sativa by modulating chlorophyll metabolism, antioxidative capacity and metal translocation. Physiol. Mol. Biol. Plants 2021, 27, 1089–1104. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, H.; Zhang, Y.; Su, Z.; Hu, T.; Liu, J.; Ding, Q.; Niu, N.; Ma, L. Methyl jasmonate enhances the safe production ability of Cd-stressed wheat by regulating the antioxidant capacity, Cd absorption, and distribution in wheat. Plant Physiol. Biochem. 2024, 212, 108788. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Wang, R.; Zhang, D.; Chu, S.; Yang, X.; Hayat, K.; Fan, Z.; Cao, X.; Ok, Y.S.; et al. Insights into growth-promoting effect of nanomaterials: Using transcriptomics and metabolomics to reveal the molecular mechanisms of MWCNTs in enhancing hyperaccumulator under heavy metal(loid)s stress. J. Hazard. Mater. 2022, 439, 129640. [Google Scholar] [CrossRef]
- Chen, X.; Shi, X.; Ai, Q.; Han, J.; Wang, H.; Fu, Q. Transcriptomic and metabolomic analyses reveal that exogenous strigolactones alleviate the response of melon root to cadmium stress. Hortic. Plant J. 2022, 8, 637–649. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, J.; Liang, Y.; Liu, J.; Jia, J.; Huo, H.; Wu, Z.; Yang, R.; Gong, H. Transcriptomic dynamics provide an insight into the mechanism for silicon mediated alleviation of salt stress in cucumber plants. Ecotoxicol. Environ. Saf. 2019, 174, 245–254. [Google Scholar] [CrossRef]
- Nie, G.; Zhou, J.; Jiang, Y.; He, J.; Wang, Y.; Liao, Z.; Appiah, C.; Li, D.; Feng, G.; Huang, L.; et al. Transcriptome characterization of candidate genes for heat tolerance in perennial ryegrass after exogenous methyl Jasmonate application. BMC Plant Biol. 2022, 22, 68. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Rainer, J.; Gatto, L.; Weichenberger, C.X. ensembldb: An R package to create and use Ensembl-based annotation resources. Bioinformatics 2019, 35, 3151–3153. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Aubert, J.; Bar-Hen, A.; Daudin, J.J.; Robin, S. Determination of the differentially expressed genes in microarray experiments using local FDR. BMC Bioinform. 2004, 5, 125. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, C.; Lan, H.; Gao, S.; Liu, H.; Liu, J.; Cao, M.; Pan, G.; Rong, T.; Zhang, S. Validation of potential reference genes for qPCR in maize across abiotic stresses, hormone treatments, and tissue types. PLoS ONE 2014, 9, e95445. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant-environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Sun, F.L.; Wang, J.H.; Geng, S.W.; Liang, Y.J.; Gong, Z.L.; Yang, N.; Qian, S.S.; Zhang, N.L.; Li, X.Y.; Wang, J.D.; et al. Comprehensive transcriptomic and metabolomic analysis revealed drought tolerance regulatory pathways in upland cotton. Front. Plant Sci. 2025, 16, 1571944. [Google Scholar] [CrossRef]
- Yu, W.; Zhou, X.R.; Meng, J.H.; Zhou, X.F.; Xu, H.W. Multi-omics research reveals the effects of the ABA-regulated phenylpropanoid biosynthesis pathway on the UV-B response in Rhododendron chrysanthum Pall. Plants 2025, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Mu, J.X.; Yang, T.; Shen, Q.; Wang, Y.K. Integrated transcriptome and metabolome analyses reveal candidate genes associate with phenolic compound biosynthesis in different varieties of Perilla frutescens. Int. J. Mol. Sci. 2025, 26, 2841. [Google Scholar] [CrossRef] [PubMed]
- Adamipour, N.; Nazari, F.; Nalousi, A.M.; da Silva, J.A.T. Evaluation of the molecular mechanism underlying proline metabolic and catabolic pathways and some morpho-physiological traits of tobacco (Nicotiana tabacum L.) plants under arsenic stress. BMC Plant Biol. 2025, 25, 258. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, R.; Bourdès, A.; Schuller, M.; Jorrin, B.; Ahel, I.; Poole, P.S. Aspartate aminotransferase of Rhizobium leguminosarum has extended substrate specificity and metabolizes aspartate to enable N2 fixation in pea nodules. Microbiol. Sgm 2024, 170, 001471. [Google Scholar] [CrossRef]
- Qiu, Z.T.; Liu, X.H.; Yu, J.; Zhao, Y.S.; Zhao, G.R.; Li, S.Y.; Liu, K.; Du, L.; Ma, L. Efficient conversion of aromatic and phenylpropanoid alcohols to acids by the cascade biocatalysis of alcohol and aldehyde dehydrogenases. Synth. Syst. Biotechnol. 2024, 9, 187–195. [Google Scholar] [CrossRef]
- Li, B.; Liang, J.; Baniasadi, H.R.; Kurihara, S.; Phillips, M.A.; Michael, A.J. Functional identification of bacterial spermine, thermospermine, norspermine, norspermidine, spermidine, and N1-aminopropylagmatine synthases. J. Biol. Chem. 2024, 300, 107281. [Google Scholar] [CrossRef]
- Chen, X.Q.; Zhao, Y.C.; Zhong, Y.Q.; Chen, J.J.; Qi, X. Deciphering the functional roles of transporter proteins in subcellular metal transportation of plants. Planta 2023, 258, 17. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.; Zhu, L.Z. Amino acid transporter as a potential carrier protein for the root-to-shoot translocation of polybrominated diphenyl ethers in rice. Environ. Sci. Technol. 2023, 57, 9722–9731. [Google Scholar] [CrossRef]
- Bittner, A.J.; Huntley, R.B.; Mourad, G.S.; Schultes, N.P. An Erwinia amylovora uracil transporter mutant retains virulence on immature apple and pear fruit. Microb. Pathog. 2020, 147, 104363. [Google Scholar] [CrossRef]
- Li, C.H.; Yue, Z.; Newstead, S.; Voth, G.A. Proton coupling and the multiscale kinetic mechanism of a peptide transporter. Biophys. J. 2022, 121, 2266–2278. [Google Scholar] [CrossRef]
- Elejalde-Palmett, C.; San Segundo, I.M.; Garroum, I.; Charrier, L.; De Bellis, D.; Mucciolo, A.; Guerault, A.; Liu, J.; Zeisler-Diehl, V.; Aharoni, A.; et al. ABCG transporters export cutin precursors for the formation of the plant cuticle. Curr. Biol. 2021, 31, 2111–2123. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.T.; Chou, J.C.C.; Oo, H.; Dassama, L.M.K. Structures and mechanisms of a novel bacterial transport system for fatty acids. Chembiochem 2023, 24, e202300156. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Newstead, S. Gateway to the Golgi: Molecular mechanisms of nucleotide sugar transporters. Curr. Opin. Struct. Biol. 2019, 57, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kourkoulou, A.; Pittis, A.A.; Diallinas, G. Evolution of substrate specificity in the nucleobase-ascorbate transporter (NAT) protein family. Microb. Cell 2018, 5, 280–292. [Google Scholar] [CrossRef]
- Lee, Y.; Nishizawa, T.; Takemoto, M.; Kumazaki, K.; Yamashita, K.; Hirata, K.; Minoda, A.; Nagatoishi, S.; Tsumoto, K.; Ishitani, R.; et al. Structure of the triose-phosphate/phosphate translocator reveals the basis of substrate specificity. Nat. Plants 2017, 3, 825–832. [Google Scholar] [CrossRef]
- Liu, M.N.; He, J.F.; He, G.H.; Zhang, Y.; Zhang, M.P.; Wang, Y.; Wang, K.Y.; Zhao, M.Z. Genome-wide identification analysis of the ATP-binding cassette transporter family and expression analysis under methyl jasmonate treatment in Panax ginseng. BMC Plant Biol. 2025, 25, 565. [Google Scholar] [CrossRef]
- Yao, X.H.; Sui, X.L.; Zhang, Y.Y. Amino acid metabolism and transporters in plant-pathogen interactions: Mechanisms and implications. Plant Cell Environ. 2025, 5, 15594. [Google Scholar] [CrossRef]
- Zhang, Q.; Shen, L.K.; Lin, F.; Liao, Q.; Xiao, S.; Zhang, W.H. Anionic phospholipid-mediated transmembrane transport and intracellular membrane trafficking in plant cells. New Phytol. 2025, 245, 1386–1402. [Google Scholar] [CrossRef]
- Yoshino, K.; Yamamoto, K.; Hara, K.; Sonoda, M.; Yamamoto, Y.; Sakamoto, K. The conservation of polyol transporter proteins and their involvement in lichenized Ascomycota. Fungal Biol. 2019, 123, 318–329. [Google Scholar] [CrossRef]
- Liu, H.; Jiao, Q.; Fan, L.; Jiang, Y.; Alyemeni, M.N.; Ahmad, P.; Chen, Y.; Zhu, M.; Liu, H.; Zhao, Y.; et al. Integrated physio-biochemical and transcriptomic analysis revealed mechanism underlying of Si-mediated alleviation to cadmium toxicity in wheat. J. Hazard. Mater. 2023, 452, 131366. [Google Scholar] [CrossRef]
- Ozturk, B.; Yildiz, K.; Ozkan, Y. Effects of pre-harvest methyl jasmonate treatments on bioactive compounds and peel color development of “fuji” apples. Int. J. Food Prop. 2015, 18, 954–962. [Google Scholar] [CrossRef]
- Zamani, H.; Arvin, M.J.; Jahromi, A.A.; Abdossi, V.; Torkashvand, A.M. The effect of sodium silicate and methyl jasmonate on pigments and antioxidant activity of tomato (Solanum lycopersicum L.) under salinity stress. J. Agric. Sci. Tarim Bilim. Derg. 2020, 26, 479–487. [Google Scholar] [CrossRef]
- He, W.; Luo, H.; Xu, H.; Zhou, Z.; Li, D.; Bao, Y.; Fu, Q.; Song, J.; Jiao, Y.; Zhang, Z. Effect of exogenous methyl jasmonate on physiological and carotenoid composition of yellow maize sprouts under NaCl stress. Food Chem. 2021, 361, 130177. [Google Scholar] [CrossRef]
- Vukmirovic, A.; Skvorc, Z.; Bogdan, S.; Krstonosic, D.; Bogdan, I.K.; Karazija, T.; Bacurin, M.; Brener, M.; Sever, K. The role of phosphorus fertilization in antioxidant responses of drought-stressed common beech and sessile oak provenances. Int. J. Mol. Sci. 2025, 26, 3053. [Google Scholar] [CrossRef]
- Tavallali, V.; Karimi, S. Methyl jasmonate enhances salt tolerance of almond rootstocks by regulating endogenous phytohormones, antioxidant activity and gas-exchange. J. Plant Physiol. 2019, 234, 98–105. [Google Scholar] [CrossRef]
- Muller, M.; Munne-Bosch, S. Hormonal impact on photosynthesis and photoprotection in plants. Plant Physiol. 2021, 185, 1500–1522. [Google Scholar] [CrossRef]
- Munoz, P.; Munne-Bosch, S. Photo-oxidative stress during leaf, flower and fruit development. Plant Physiol. 2018, 176, 1004–1014. [Google Scholar] [CrossRef]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef]
- Zhao, L.S.; Wang, N.; Li, K.; Li, C.Y.; Guo, J.P.; He, F.Y.; Liu, G.M.; Chen, X.L.; Gao, J.; Liu, L.N.; et al. Architecture of symbiotic dinoflagellate photosystem I-light-harvesting supercomplex in Symbiodinium. Nat. Commun. 2024, 15, 2392. [Google Scholar] [CrossRef]
- Rumbaugh, T.D.; Gorka, M.J.; Baker, C.S.; Golbeck, J.H.; Silakov, A. Light-induced H2 generation in a photosystem I-O2-tolerant FeFe hydrogenase nanoconstruct. Proc. Natl. Acad. Sci. USA 2024, 121, e2400267121. [Google Scholar] [CrossRef]
- Dai, G.Z.; Song, W.Y.; Xu, H.F.; Tu, M.; Yu, C.; Li, Z.K.; Shang, J.L.; Jin, C.L.; Ding, C.S.; Zuo, L.Z.; et al. Hypothetical chloroplast reading frame 51 encodes a photosystem I assembly factor in cyanobacteria. Plant Cell 2024, 36, 1844–1867. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Lv, G. Nitraria sibirica adapts to long-term soil water deficit by reducing photosynthesis, stimulating antioxidant systems, and accumulating osmoregulators. Plant Physiol. Biochem. 2024, 206, 108265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, Y.; Jing, T.; Liu, X.; Ai, X.; Bi, H. Melatonin promotes the chilling tolerance of cucumber seedlings by regulating antioxidant system and relieving photoinhibition. Front. Plant Sci. 2021, 12, 789617. [Google Scholar] [CrossRef] [PubMed]
- Trotta, A.; Redondo-Gomez, S.; Pagliano, C.; Figueroa Clemente, M.E.; Rascio, N.; La Rocca, N.; Antonacci, A.; Andreucci, F.; Barbato, R. Chloroplast ultrastructure and thylakoid polypeptide composition are affected by different salt concentrations in the halophytic plant Arthrocnemum macrostachyum. J. Plant Physiol. 2012, 169, 111–116. [Google Scholar] [CrossRef]
- Chen, J.H.; Chen, S.T.; He, N.Y.; Wang, Q.L.; Zhao, Y.; Gao, W.; Guo, F.Q. Nuclear-encoded synthesis of the D1 subunit of photosystem II increases photosynthetic efficiency and crop yield. Nat. Plants 2020, 6, 570–580. [Google Scholar] [CrossRef]
- Semin, B.K.; Davletshina, L.N. High-efficiency oxygen evolution by photosystem II oxygen-evolving complex containing 3Mn per reaction center. J. Biol. Inorg. Chem. 2023, 28, 393–401. [Google Scholar] [CrossRef]
- Arshad, F.; Eaton-Rye, J.J. Indirect interactions involving the PsbM or PsbT subunits and the PsbO, PsbU and PsbV proteins stabilize assembly and activity of Photosystem II in Synechocystis sp. PCC 6803. Photosynth. Res. 2024, 160, 61–75. [Google Scholar] [CrossRef]
- Zhao, M.; Jin, J.; Wang, J.; Gao, T.; Luo, Y.; Jing, T.; Hu, Y.; Pan, Y.; Lu, M.; Schwab, W.; et al. Eugenol functions as a signal mediating cold and drought tolerance via UGT71A59-mediated glucosylation in tea plants. Plant J. 2022, 109, 1489–1506. [Google Scholar] [CrossRef]
- Ali, S.; Ali, H.; Hussain, S. Influence of osmoprotectants and their different application methods on wheat (Triticumaestivum L.) cultivars under water deficiency in south punjab region of pakistan. Appl. Ecol. Environ. Res. 2023, 21, 3895–3910. [Google Scholar] [CrossRef]
- Dube, S.P.; Sibiya, J.; Kutu, F. Genetic diversity and population structure of maize inbred lines using phenotypic traits and single nucleotide polymorphism (SNP) markers. Sci. Rep. 2023, 13, 17851. [Google Scholar] [CrossRef]
- Turkoglu, A.; Haliloglu, K.; Mohammadi, S.A.; Ozturk, A.; Bolouri, P.; Ozkan, G.; Bocianowski, J.; Pour-Aboughadareh, A.; Jamshidi, B. Genetic diversity and population structure in turkiye bread wheat genotypes revealed by simple sequence repeats (SSR) markers. Genes 2023, 14, 1182. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, L.; Wu, Z.; Chen, J.; Wang, T.; Zhang, X.; Mei, G.; Wang, J.; Lv, G. Classification and expression profile of the U-Box E3 ubiquitin ligase enzyme gene family in maize (Zea mays L.). Plants 2022, 11, 2459. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.C.; Yang, Y.T.; Wu, R.; Han, B.X.; Wu, D.L.; Ou, J.M. Transcriptome analysis of Akebia quinata (Thunb.) Decne. and discovery of key enzyme genes in the phenylpropanoid biosynthesis pathway. Acta Physiol. Plant. 2023, 45, 117. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, J.; Wei, F.; Song, T.; Yu, Y.; Yu, M.; Fan, Q.; Yang, Y.; Xue, G.; Zhang, X. Nucleoredoxin gene TaNRX1 positively regulates drought tolerance in transgenic wheat (Triticum aestivum L.). Front. Plant Sci. 2021, 12, 756338. [Google Scholar] [CrossRef]
- Jin, S.; Ye, N.; Chen, X.; Wang, P.; Gu, M.; Hou, B.; Zhang, C.; Zheng, Y.; Sun, Y. Identification of PAL genes related to anthocyanin synthesis in tea plants and its correlation with anthocyanin content. Hortic. Plant J. 2022, 8, 381–394. [Google Scholar] [CrossRef]
- Tian, J.; Xu, R.; Chang, K.; Yuan, S.; Huang, C.; Wang, J.; Li, S.; Liu, F.; Zhong, F. Identification of PAL gene in purple cabbage and functional analysis related to anthocyanin synthesis. Horticulturae 2023, 9, 469. [Google Scholar] [CrossRef]
- Cui, X.; Tang, M.; Li, L.; Chang, J.; Yang, X.; Chang, H.; Zhou, J.; Liu, M.; Wang, Y.; Zhou, Y.; et al. Expression patterns and molecular mechanisms regulating drought tolerance of soybean Glycine max (L.) Merr. conferred by transcription factor gene GmNAC19. Int. J. Mol. Sci. 2024, 25, 2396. [Google Scholar] [CrossRef]
- Han, X.; Wu, Z.; Liu, F.; Wang, Y.; Wei, X.; Tian, P.; Ling, F. Transcriptomic analysis and salt-tolerance gene mining during rice germination. Genes 2023, 14, 1556. [Google Scholar] [CrossRef]
- Gu, L.; Chen, X.; Hou, Y.; Cao, Y.; Wang, H.; Zhu, B.; Du, X.; Wang, H. ZmWRKY30 modulates drought tolerance in maize by influencing myo-inositol and reactive oxygen species homeostasis. Physiol. Plant. 2024, 176, e14423. [Google Scholar] [CrossRef]
- Amist, N.; Khare, S.; Azim, Z.; Singh, N.B. Protective role of polyethylene glycol towards the damaging effects of cadmium. Appl. Biochem. Biotechnol. 2024, 197, 113–136. [Google Scholar] [CrossRef]
- Golovatskaya, I.F.; Kadyrbaev, M.K.; Boyko, E.V. Protective role of melatonin and IAA in the regulation of resistance of potato regenerants to cold stress. Potato Res. 2024, 67, 421–449. [Google Scholar] [CrossRef]
- Wang, D.; Ni, Y.; Xie, K.; Li, Y.; Wu, W.; Shan, H.; Cheng, B.; Li, X. Aquaporin ZmTIP2;3 Promotes Drought Resistance of Maize through Symbiosis with Arbuscular Mycorrhizal Fungi. Int. J. Mol. Sci. 2024, 25, 4205. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Jiang, F.; Okla, M.K.; Abbas, Z.K.; Qahtani, S.M.A.; Al-Harbi, N.A.; Abdel-Maksoud, M.A.; Olivan, L.M.G. Nanoparticles synergy: Enhancing wheat (Triticum aestivum L.) cadmium tolerance with iron oxide and selenium. Sci. Total Environ. 2024, 915, 169869. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Saleem, M.H.; Khalid, M.R.; Ali, B.; Fahad, S. Nitric oxide reduces cadmium uptake in wheat (Triticum aestivum L.) by modulating growth, mineral uptake, yield attributes, and antioxidant profile. Environ. Sci. Pollut. Res. 2024, 31, 9844–9856. [Google Scholar] [CrossRef]
- Alvarez, M.E.; Savoure, A.; Szabados, L. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, P.; Su, W.; Zhang, H.; Xu, W.; Chu, X. Metabolic engineering strategy for synthetizing trans-4-hydroxy-l-proline in microorganisms. Microb. Cell Factories 2021, 20, 87. [Google Scholar] [CrossRef]
- Kim, T.L.; Lim, H.; Denison, M.I.J.; Oh, C. Transcriptomic and physiological analysis reveals genes associated with drought stress responses in populus alba x populus glandulosa. Plants 2023, 12, 3238. [Google Scholar] [CrossRef]
- Qin, S.; Liu, H.; Nie, Z.; Rengel, Z.; Gao, W.; Li, C.; Zhao, P. Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: A review. Pedosphere 2020, 30, 168–180. [Google Scholar] [CrossRef]
- Qin, S.; Liu, H.; Rengel, Z.; Gao, W.; Nie, Z.; Li, C.; Hou, M.; Cheng, J.; Zhao, P. Boron inhibits cadmium uptake in wheat (Triticum aestivum) by regulating gene expression. Plant Sci. 2020, 297, 110522. [Google Scholar] [CrossRef]
- Bari, M.A.; Prity, S.A.; Das, U.; Akther, M.S.; Sajib, S.A.; Reza, M.A.; Kabir, A.H. Silicon induces phytochelatin and ROS scavengers facilitating cadmium detoxification in rice. Plant Biol. 2020, 22, 472–479. [Google Scholar] [CrossRef]
- Gonzalez, D.A.; de la Torre, V.S.G.; Fernandez, R.R.; Barreau, L.; Merlot, S. Divergent roles of IREG/Ferroportin transporters from the nickel hyperaccumulator Leucocroton havanensis. Physiol. Plant. 2024, 176, e14261. [Google Scholar] [CrossRef] [PubMed]
- Mani, A.; Sankaranarayanan, K. In silico analysis of natural resistance-associated macrophage protein (NRAMP) family of transporters in rice. Protein J. 2018, 37, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Noor, I.; Sohail, H.; Wentao, C.; Zhu, K.; Hasanuzzaman, M.; Li, G.; Liu, J. Phosphorus-induced restructuring of the ascorbate-glutathione cycle and lignin biosynthesis alleviates manganese toxicity in peach roots. Tree Physiol. 2024, 44, tpae098. [Google Scholar] [CrossRef] [PubMed]
- Rono, J.K.; Wang, L.L.; Wu, X.C.; Cao, H.W.; Zhao, Y.N.; Khan, I.U.; Yang, Z.M. Identification of a new function of metallothionein-like gene OsMT1e for cadmium detoxification and potential phytoremediation. Chemosphere 2021, 265, 129136. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Lan, S.; Sha, J.; Zhang, Z.; Chang, Y.; Chen, Y.; Yang, S. Transcriptome Analysis Reveals the Molecular Mechanisms Through Which Exogenous Methyl Jasmonate Enhances Mercury Stress Tolerance in Maize Seedlings. Agronomy 2025, 15, 1369. https://doi.org/10.3390/agronomy15061369
Liu X, Lan S, Sha J, Zhang Z, Chang Y, Chen Y, Yang S. Transcriptome Analysis Reveals the Molecular Mechanisms Through Which Exogenous Methyl Jasmonate Enhances Mercury Stress Tolerance in Maize Seedlings. Agronomy. 2025; 15(6):1369. https://doi.org/10.3390/agronomy15061369
Chicago/Turabian StyleLiu, Xingyuan, Shanshan Lan, Jin Sha, Zuosheng Zhang, Yanmei Chang, Yunliang Chen, and Shuanglong Yang. 2025. "Transcriptome Analysis Reveals the Molecular Mechanisms Through Which Exogenous Methyl Jasmonate Enhances Mercury Stress Tolerance in Maize Seedlings" Agronomy 15, no. 6: 1369. https://doi.org/10.3390/agronomy15061369
APA StyleLiu, X., Lan, S., Sha, J., Zhang, Z., Chang, Y., Chen, Y., & Yang, S. (2025). Transcriptome Analysis Reveals the Molecular Mechanisms Through Which Exogenous Methyl Jasmonate Enhances Mercury Stress Tolerance in Maize Seedlings. Agronomy, 15(6), 1369. https://doi.org/10.3390/agronomy15061369