
How the Functional Constitution of Plant Communities in Field Margins Affects Wild Bee Community Composition and Functional Structure



Abstract
1. Introduction
- To determine which general characteristics of the plant communities of cereal margins are important for bees (objective 1a) and how these characteristics influence the functional structure of bee communities (objective 1b);
- To understand the role of functional diversity and resource accessibility in the plant communities of cereal margins and how these factors influence wild bee abundance and community composition (objective 2a), as well as the functional structure of the wild bee community (objective 2b).
2. Materials and Methods
2.1. Study Sites
- They were well established margins, more than one-metre wide.
- They were located between two cereal crops, between a cereal crop and fallow land, or between a cereal crop and a low traffic country road.
- The distance between the sampled margins was greater than two kilometres. This distance was chosen because the typical foraging range of most wild bee species is less than one kilometre [24]. This approach allowed us to reasonably assume that the bee community sampled at each site each year was likely distinct [25].
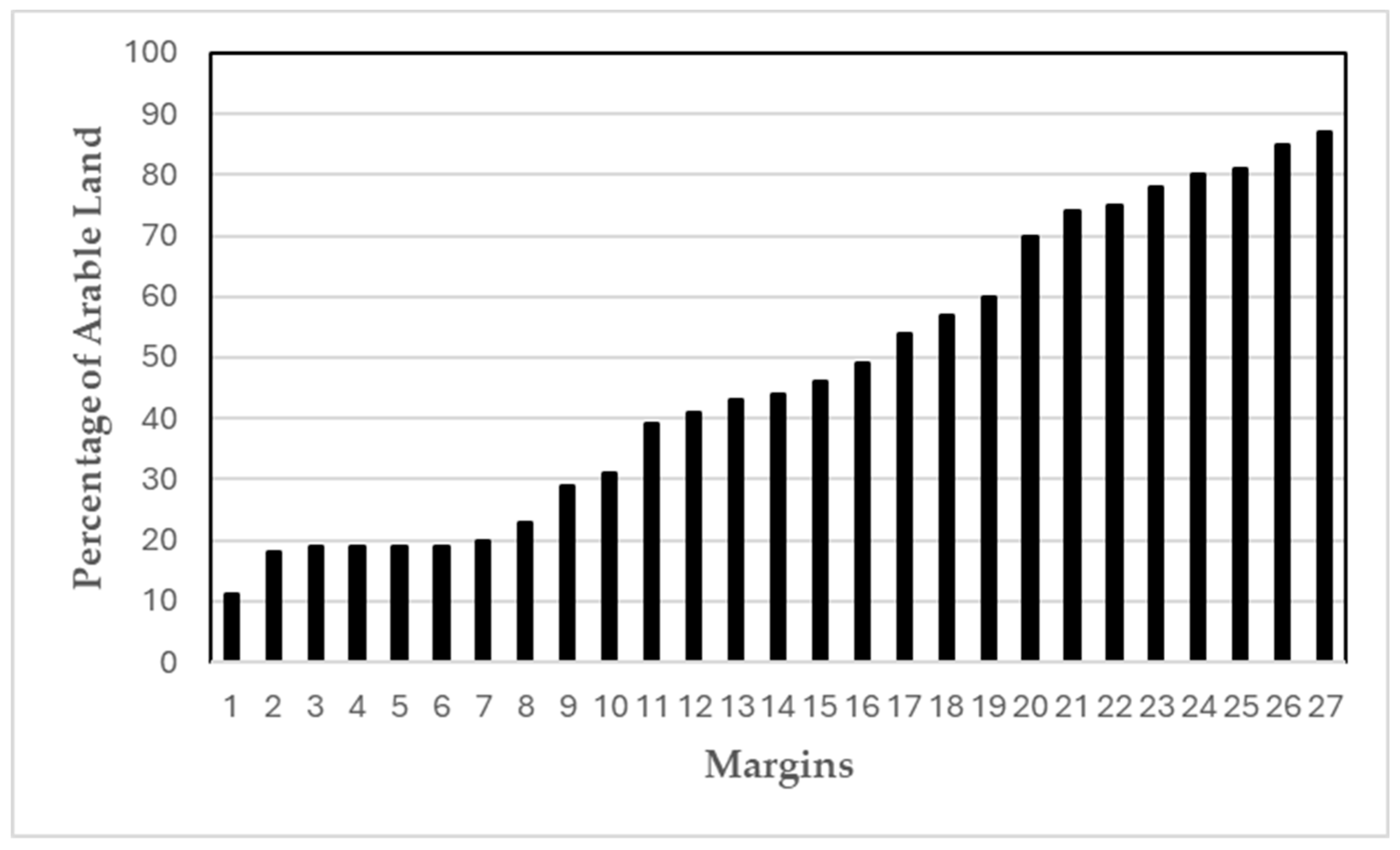
- The landscapes surrounding the margins show a gradient in percentage of surrounding arable land in order to have a good representation of margins from landscapes with different agricultural intensities.
2.2. Sampling of the Wild Bee Community
2.3. Observations of Bee Foraging Activity in Margins
2.4. Sampling of the Vegetation Community
2.5. Landscape Structure
2.6. Functional Trait Analysis
2.6.1. Vegetation
2.6.2. Bee Genera
2.7. Statistical Analysis
3. Results
4. Discussion
4.1. General Observations
4.2. Relationships Between General Characteristics of Plant Communities and Wild Bee Abundance, Community Composition and Functional Structure (Objective 1)
4.3. Relationships Between Functional Characteristics of Plant Communities and Wild Bee Abundance, Community Composition and Functional Structure (Objective 2)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FDis | Functional dispersion |
| CWM | Community-weighted mean |
| EST | Estimate |
| SE | Standard error |
Appendix A

{kind=link}
| General Characteristics (Predictor Variables) | Wild Bee Community (Response Variables) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CWM Sociality | CWM Lecty | CWM Body Size | CWM Tongue Length | CWM Parasitism | FDis Body Size | FDis Tongue Length | FDis Parasitism | FDis Pollen Organ | ||||||||||
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | F | p | F | p | |
| FDis Mean Height | 0.0 | 0.94 | 0.0 | 1.00 | 0.9 | 0.35 | 1.3 | 0.27 | 2.9 | 0.11 | 0.1 | 0.73 | 0.5 | 0.50 | 0.7 | 0.40 | 0.3 | 0.60 |
| FDis Corolla Size | 3.7 | 0.07 | 2.7 | 0.11 | 2.2 | 0.16 | 3.3 | 0.08 | 1.8 | 0.20 | 0.5 | 0.48 | 0.5 | 0.48 | 2.9 | 0.11 | 3.7 | 0.07 |
| FDis Flower Colour | 0.2 | 0.66 | 0.2 | 0.67 | 0.8 | 0.39 | 2.2 | 0.16 | 1.0 | 0.32 | 0.5 | 0.47 | 0.5 | 0.47 | 0.3 | 0.60 | 2.9 | 0.10 |
| CWM Nectar Accessibility | 0.3 | 0.60 | 3.7 | 0.07 | 1.4 | 0.25 | 0.0 | 0.84 | 0.0 | 0.97 | 1.0 | 0.32 | 0.4 | 0.54 | 0.5 | 0.48 | 0.7 | 0.41 |
| FDis Morphology | 1.0 | 0.33 | 1.3 | 0.27 | 2.6 | 0.12 | 2.7 | 0.12 | 0.0 | 0.92 | 2.8 | 0.11 | 3.0 | 0.10 | 1.7 | 0.21 | 2.5 | 0.13 |
| Adjusted R2 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.0 | 0.0 | 0.1 | 0.2 | |||||||||
References
- Feledyn-Szewczyk, B.; Kuś, J.; Stalenga, J.; Berbeć, A.K.; Radzikowski, P. The Role of Biological Diversity in Agroecosystems and Organic Farming. In Organic Farming: A Promising Way of Food Production; Konvalina, P., Ed.; IntechOpen: Rijeka, Croatia, 2016. [Google Scholar]
- Balzan, M.V.; Bocci, G.; Moonen, A.-C. Augmenting Flower Trait Diversity in Wildflower Strips to Optimise the Conservation of Arthropod Functional Groups for Multiple Agroecosystem Services. J. Insect Conserv. 2014, 18, 713–728. [Google Scholar] [CrossRef]
- Senapathi, D.; Fründ, J.; Albrecht, M.; Garratt, M.P.D.; Kleijn, D.; Pickles, B.J.; Potts, S.G.; An, J.; Andersson, G.K.S.; Bänsch, S.; et al. Wild Insect Diversity Increases Inter-Annual Stability in Global Crop Pollinator Communities. Proc. R. Soc. B Biol. Sci. 2021, 288, 20210212. [Google Scholar] [CrossRef] [PubMed]
- Shivanna, K.R.; Tandon, R.; Koul, M. Diversity. In Reproductive Ecology of Flowering Plants: Patterns and Processes; Tandon, R., Shivanna, K., Koul, M., Eds.; Springer Singapore: Singapore, 2020; pp. 395–413. [Google Scholar]
- Ricketts, T.H.; Lonsdorf, E. Mapping the Margin: Comparing Marginal Values of Tropical Forest Remnants for Pollination Services. Ecol. Appl. 2013, 23, 1113–1123. [Google Scholar] [CrossRef]
- Koh, I.; Lonsdorf, E.V.; Williams, N.M.; Brittain, C.; Isaacs, R.; Gibbs, J.; Ricketts, T.H. Modeling the Status, Trends, and Impacts of Wild Bee Abundance in the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, C.I.; Altieri, M.A. Plant Biodiversity Enhances Bees and Other Insect Pollinators in Agroecosystems. A Review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow Restoration Promotes Pollinator Populations and Exports Native Bees to Adjacent Fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef]
- Cirujeda, A.; Pardo, G. The Role of Field Margins in Biodiversity Conservation in Agroecosystems. In Reconciling Agricultural Production with Biodiversity Conservation; Bàrberi, P., Moonen, A.-C., Eds.; Burleigh Dodds Science Publishing: London, UK, 2020; pp. 146–176. [Google Scholar]
- Lagerlöf, J.; Stark, J.; Svensson, B. Margins of Agricultural Fields as Habitats for Pollinating Insects. Agric. Ecosyst. Environ. 1992, 40, 117–124. [Google Scholar] [CrossRef]
- McKechnie, I.M.; Thomsen, C.J.M.; Sargent, R.D. Forested Field Edges Support a Greater Diversity of Wild Pollinators in Lowbush Blueberry (Vaccinium Angustifolium). Agric. Ecosyst. Environ. 2017, 237, 154–161. [Google Scholar] [CrossRef]
- Sanchez, J.; Carrasco, A.; La Spina, M.; Pérez-Marcos, M.; Ortiz-Sánchez, F. How Bees Respond Differently to Field Margins of Shrubby and Herbaceous Plants in Intensive Agricultural Crops of the Mediterranean Area. Insects 2019, 11, 26. [Google Scholar] [CrossRef]
- Campbell, A.J.; Biesmeijer, J.C.; Varma, V.; Wäckers, F.L. Realising Multiple Ecosystem Services Based on the Response of Three Beneficial Insect Groups to Floral Traits and Trait Diversity. Basic. Appl. Ecol. 2012, 13, 363–370. [Google Scholar] [CrossRef]
- Bosch, J.; Retana, J.; Cerdá, X. Flowering Phenology, Floral Traits and Pollinator Composition in a Herbaceous Mediterranean Plant Community. Oecologia 1997, 109, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.; Izquierdo, J.; Hernández Plaza, E.; González-Andújar, J.L. The Attractiveness of Five Common Mediterranean Weeds to Pollinators. Agronomy 2021, 11, 1314. [Google Scholar] [CrossRef]
- Sammataro, D.; Yoder, J. Honey Bee Colony Health: Challenges and Sustainable Solutions; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Real, L. Pollination Biology; Elsevier: Raleigh, NA, USA, 2012. [Google Scholar]
- Dafni, A.; Potts, S.G. The Role of Flower Inclination, Depth, and Height in the Preferences of a Pollinating Beetle (Coleoptera: Glaphyridae). J. Insect Behav. 2004, 17, 823–834. [Google Scholar] [CrossRef]
- José-María, L.; Armengot, L.; Blanco-Moreno, J.M.; Bassa, M.; Sans, F.X. Effects of Agricultural Intensification on Plant Diversity in Mediterranean Dryland Cereal Fields. J. Appl. Ecol. 2010, 47, 832–840. [Google Scholar] [CrossRef]
- Pallavicini, Y.; Bastida, F.; Hernández-Plaza, E.; Petit, S.; Izquierdo, J.; Gonzalez-Andujar, J.L. Local Factors Rather than the Landscape Context Explain Species Richness and Functional Trait Diversity and Responses of Plant Assemblages of Mediterranean Cereal Field Margins. Plants 2020, 9, 778. [Google Scholar] [CrossRef]
- Cirujeda, A.; Pardo, G.; Marí, A.I.; Aibar, J.; Pallavicini, Y.; González-Andújar, J.L.; Recasens, J.; Solé-Senan, X.O. The Structural Classification of Field Boundaries in Mediterranean Arable Cropping Systems Allows the Prediction of Weed Abundances in the Boundary and in the Adjacent Crop. Weed Res. 2019, 59, 300–311. [Google Scholar] [CrossRef]
- Morrison, J. Flowering Native Weeds for the Conservation of Wild Pollinators in Agroecosystems; Universitat Politècnica de Catalunya: Castelldefels, Spain, 2018. [Google Scholar]
- Ruralcat. Available online: http://ruralcat.gencat.cat (accessed on 1 March 2023).
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee Foraging Ranges and Their Relationship to Body Size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Riedinger, V.; Mitesser, O.; Hovestadt, T.; Steffan-Dewenter, I.; Holzschuh, A. Annual Dynamics of Wild Bee Densities: Attractiveness and Productivity Effects of Oilseed Rape. Ecology 2015, 96, 1351–1360. [Google Scholar] [CrossRef]
- Pywell, R.F.; Warman, E.A.; Carvell, C.; Sparks, T.H.; Dicks, L.V.; Bennett, D.; Wright, A.; Critchley, C.N.R.; Sherwood, A. Providing Foraging Resources for Bumblebees in Intensively Farmed Landscapes. Biol. Conserv. 2005, 121, 479–494. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring Bee Diversity in Different European Habitats and Biogeographical Regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef]
- Gezon, Z.J.; Wyman, E.S.; Ascher, J.S.; Inouye, D.W.; Irwin, R.E. The Effect of Repeated, Lethal Sampling on Wild Bee Abundance and Diversity. Methods Ecol. Evol. 2015, 6, 1044–1054. [Google Scholar] [CrossRef]
- Ascher, J.S.; Pickering, J. Discover Life Bee Species and World Checklist (Hymenoptera: Apoidea: Anthophila). Available online: https://www.discoverlife.org (accessed on 1 September 2014).
- Cane, J.H. Estimation of Bee Size Using Intertegular Span (Apoidea). J. Kans. Entomol. Soc. 1987, 60, 145–147. [Google Scholar]
- van Rijn, I.; Neeson, T.M.; Mandelik, Y. Reliability and Refinement of the Higher Taxa Approach for Bee Richness and Composition Assessments. Ecol. Appl. 2015, 25, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Chiu, C.H.; Jost, L. Unifying Species Diversity, Phylogenetic Diversity, Functional Diversity, and Related Similarity and Differentiation Measures through Hill Numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef]
- Tutin, T. Flora Europaea; Cambridge University Press: Cambridge, UK, 1964. [Google Scholar]
- Sistema de Información Geográfica de Parcelas Agrícolas. 1 March. Available online: http://sigpac.mapa.es/fega/visor/ (accessed on 1 March 2023).
- Gous, A.; Eardley, C.D.; Johnson, S.D.; Swanevelder, D.Z.H.; Willows-Munro, S. Floral Hosts of Leaf-Cutter Bees (Megachilidae) in a Biodiversity Hotspot Revealed by Pollen DNA Metabarcoding of Historic Specimens. PLoS ONE 2021, 16, e0244973. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A Distance-based Framework for Measuring Functional Diversity from Multiple Traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing Functional Diversity in the Field—Methodology Matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- R Development Core Team, R. A Language and Environment for Statistical Computing; R Development Core Team R: Auckland, New Zealand, 2021. [Google Scholar]
- Williams, N.M.; Crone, E.E.; Roulston, T.H.; Minckley, R.L.; Packer, L.; Potts, S.G. Ecological and Life-History Traits Predict Bee Species Responses to Environmental Disturbances. Biol. Conserv. 2010, 143, 2280–2291. [Google Scholar] [CrossRef]
- Russell, K.N.; Ikerd, H.; Droege, S. The Potential Conservation Value of Unmowed Powerline Strips for Native Bees. Biol. Conserv. 2005, 124, 133–148. [Google Scholar] [CrossRef]
- Forrest, J.R.K.; Thorp, R.W.; Kremen, C.; Williams, N.M. Contrasting Patterns in Species and Functional-trait Diversity of Bees in an Agricultural Landscape. J. Appl. Ecol. 2015, 52, 706–715. [Google Scholar] [CrossRef]
- Kuppler, J.; Neumüller, U.; Mayr, A.V.; Hopfenmüller, S.; Weiss, K.; Prosi, R.; Schanowski, A.; Schwenninger, H.-R.; Ayasse, M.; Burger, H. Favourite Plants of Wild Bees. Agric. Ecosyst. Environ. 2023, 342, 108266. [Google Scholar] [CrossRef]
- Morrison, J.; Izquierdo, J.; Plaza, E.H.; González-Andújar, J.L. The Role of Field Margins in Supporting Wild Bees in Mediterranean Cereal Agroecosystems: Which Biotic and Abiotic Factors Are Important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models; The Comprehensive R Archive Network: Berlin, Germany, 2022. [Google Scholar]
- Droege, S.; Tepedino, V.J.; Lebuhn, G.; Link, W.; Minckley, R.L.; Chen, Q.; Conrad, C. Spatial Patterns of Bee Captures in North American Bowl Trapping Surveys. Insect Conserv. Divers. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Le Féon, V.; Poggio, S.L.; Torretta, J.P.; Bertrand, C.; Molina, G.A.R.; Burel, F.; Baudry, J.; Ghersa, C.M. Diversity and Life-History Traits of Wild Bees (Insecta: Hymenoptera) in Intensive Agricultural Landscapes in the Rolling Pampa, Argentina. J. Nat. Hist. 2016, 50, 1175–1196. [Google Scholar] [CrossRef]
- Nielsen, A.; Steffan-Dewenter, I.; Westphal, C.; Messinger, O.; Potts, S.G.; Roberts, S.P.M.; Settele, J.; Szentgyörgyi, H.; Vaissière, B.E.; Vaitis, M.; et al. Assessing Bee Species Richness in Two Mediterranean Communities: Importance of Habitat Type and Sampling Techniques. Ecol. Res. 2011, 26, 969–983. [Google Scholar] [CrossRef]
- Kleijn, D.; Winfree, R.; Bartomeus, I.; Carvalheiro, L.G.; Henry, M.; Isaacs, R.; Klein, A.-M.; Kremen, C.; M’Gonigle, L.K.; Rader, R.; et al. Delivery of Crop Pollination Services Is an Insufficient Argument for Wild Pollinator Conservation. Nat. Commun. 2015, 6, 7414. [Google Scholar] [CrossRef]
- Ullmann, K.S.; Cane, J.H.; Thorp, R.W.; Williams, N.M. Soil Management for Ground-Nesting Bees. In Towards Sustainable Crop Pollination Services—Measures at Field, Farm and Landscape Scales; Gemmill-Herren, B., Azzu, N., Bicksler, A., Guidotti, A., Eds.; FAO: Rome, Italy, 2020; ISBN 978-92-5-132578-0. [Google Scholar]
- Carvell, C.; Osborne, J.L.; Bourke, A.F.G.; Freeman, S.N.; Pywell, R.F.; Heard, M.S. Bumble Bee Species’ Responses to a Targeted Conservation Measure Depend on Landscape Context and Habitat Quality. Ecol. Appl. 2011, 21, 1760–1771. [Google Scholar] [CrossRef]
- Hannon, L.E.; Sisk, T.D. Hedgerows in an Agri-Natural Landscape: Potential Habitat Value for Native Bees. Biol. Conserv. 2009, 142, 2140–2154. [Google Scholar] [CrossRef]
- Rosa García, R.; Miñarro, M. Role of Floral Resources in the Conservation of Pollinator Communities in Cider-Apple Orchards. Agric. Ecosyst. Environ. 2014, 183, 118–126. [Google Scholar] [CrossRef]
- Chamorro, F.J.; Faria, C.M.A.; Araújo, F.S.; Freitas, B.M. Elevated Pan Traps Optimise the Sampling of Bees, Including When the Availability of Floral Resources Is High. Insect Conserv. Divers. 2023, 16, 16–32. [Google Scholar] [CrossRef]
- Ogilvie, J.E.; Forrest, J.R. Interactions between Bee Foraging and Floral Resource Phenology Shape Bee Populations and Communities. Curr. Opin. Insect Sci. 2017, 21, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial Community Evenness Favours Functionality under Selective Stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef] [PubMed]
| Trait | Categories |
|---|---|
| Nectar Accessibility | None, Concealed, Partly Exposed, Fully Exposed |
| Flower Colour | Blue/Dark Purple, Green, Orange/Red, Pink/Light Purple, White/Cream, Yellow |
| Height (Mean) | Continuous |
| Corolla Size (Mean Radius) | Continuous |
| Flower Morphology | Bell, Disk, Flag, Funnel, Lip, Ray, Ray and Disk |
| Trait | Categories |
|---|---|
| Sociality | 1 (only social 1 species) 0.5 (both social and solitary species) 0 (only solitary species) |
| Lecty (diet specialization) 2 | 1 (only oligolectic species) 0.5 (both oligolectic and polylectic species) 0 (only polylectic species) |
| Body Size (Median ITD) | Continuous |
| Tongue Length (Median) | Continuous |
| Parasitism | 1 (cleptoparasitic) 0 (non-parasitic) |
| Pollen Organ | Corbicula, Crop (“honey stomach”), Scopa Abdomen (scopa found on abdomen), Scopa Leg (scopa found on leg), Scopa Abdomen & Leg (scopa found on both the abdomen and the leg), Scopa Leg & Propodeum (scopa found on both the leg and the propodeum), Scopa & Floccus Leg (scopa and floccus found on the leg), None |
| Across Individual Margins | Overall | |||||
|---|---|---|---|---|---|---|
| Total Bee Abundance | Morphospecies Richness | Morphospecies Diversity | Morphospecies Evenness | Body Size (ITD; mm) | Tongue Length (mm) | |
| Min. | 38 | 9 | 3.2 | 0.45 | 0.4 | 0.6 |
| Max. | 295 | 31 | 17.8 | 0.93 | 7.7 | 15.1 |
| Mean | 129 | 21 | 11.2 | 0.78 | 1.8 | 2.5 |
| General Characteristics (Predictor Variables) | Wild Bee Community (Response Variables) | |||||||
|---|---|---|---|---|---|---|---|---|
| Abundance | Morphospecies Diversity | Morphospecies Evenness | Foraging Activity 1 | |||||
| F | p | F | p | F | p | Chi-sq | p | |
| % Arable Land | 6.0 | 0.02 (+) | 0.3 | 0.60 | 1.5 | 0.23 | 1.5 | 0.23 |
| % Non-herbaceous Plants | 0.3 | 0.61 | 4.2 | 0.05 | 0.7 | 0.43 | 0.24 | 0.63 |
| Floral Richness | 0.0 | 0.87 | 0.0 | 0.91 | 0.0 | 0.94 | 5.7 | 0.02 (+) |
| % Important Families | 0.1 | 0.78 | 0.1 | 0.77 | 0.3 | 0.62 | 0.9 | 0.34 |
| Adjusted R2 | 0.1 | 0.2 | 0.1 | |||||
| General Characteristics (Predictor Variables) | Wild Bee Community (Response Variables) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CWM Sociality | CWM Lecty | CWM Body Size | CWM Tongue Length | CWM Parasitism | FDis Body Size | FDis Tongue Length | FDis Parasitism | FDis Pollen Organ | ||||||||||
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | F | p | F | p | |
| % Arable Land | 0.1 | 0.71 | 0.4 | 0.51 | 4.1 | 0.05 | 8.1 | 0.009 (−) | 7.0 | 0.02 (−) | 12.7 | 0.002 (−) | 12.0 | 0.002 (−) | 6.3 | 0.02 (−) | 3.9 | 0.06 |
| % Non-herbaceous Plants | 2.6 | 0.12 | 0.6 | 0.45 | 1.6 | 0.22 | 1.0 | 0.32 | 1.4 | 0.24 | 0.1 | 0.78 | 0.1 | 0.74 | 1.6 | 0.21 | 0.0 | 0.99 |
| Floral Richness | 0.4 | 0.52 | 0.9 | 0.35 | 0.0 | 0.88 | 0.9 | 0.35 | 3.3 | 0.08 | 0.1 | 0.76 | 0.3 | 0.57 | 2.2 | 0.15 | 1.1 | 0.31 |
| % Important Families | 0.0 | 0.96 | 0.4 | 0.56 | 0.3 | 0.59 | 0.1 | 0.80 | 7.2 | 0.01 (+) | 0.2 | 0.64 | 0.1 | 0.72 | 6.7 | 0.02 (+) | 0.6 | 0.46 |
| Adjusted R2 | 0.0 | 0.1 | 0.1 | 0.2 | 0.4 | 0.3 | 0.3 | 0.4 | 0.3 | |||||||||
| Functional Characteristics (Predictor Variables) | Wild Bee Community (Response Variables) | |||||||
|---|---|---|---|---|---|---|---|---|
| Abundance | Morphospecies Diversity | Morphospecies Evenness | Foraging Activity 1 | |||||
| F | p | F | p | F | p | Chi-sq | p | |
| FDis Mean Height | 0.8 | 0.37 | 0.0 | 0.84 | 0.2 | 0.67 | 0.1 | 0.76 |
| FDis Corolla Size | 0.2 | 0.63 | 4.5 | 0.05 | 2.2 | 0.15 | 2.2 | 0.14 |
| FDis Flower Colour | 0.9 | 0.36 | 2.1 | 0.16 | 5.2 | 0.03 (+) | 0.4 | 0.53 |
| CWM Nectar Accessibility | 2.3 | 0.14 | 0.2 | 0.66 | 0.3 | 0.59 | 11.8 | 0.001 (+) |
| FDis Morphology | 0.1 | 0.76 | 0.8 | 0.37 | 0.2 | 0.68 | 14.6 | 0.0001 (+) |
| Adjusted R2 | 0.1 | 0.1 | 0.2 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrison, J.; Izquierdo, J.; Hernández Plaza, E.; González-Andújar, J.L. How the Functional Constitution of Plant Communities in Field Margins Affects Wild Bee Community Composition and Functional Structure. Agronomy 2025, 15, 1354. https://doi.org/10.3390/agronomy15061354
Morrison J, Izquierdo J, Hernández Plaza E, González-Andújar JL. How the Functional Constitution of Plant Communities in Field Margins Affects Wild Bee Community Composition and Functional Structure. Agronomy. 2025; 15(6):1354. https://doi.org/10.3390/agronomy15061354
Chicago/Turabian StyleMorrison, Jane, Jordi Izquierdo, Eva Hernández Plaza, and José L. González-Andújar. 2025. "How the Functional Constitution of Plant Communities in Field Margins Affects Wild Bee Community Composition and Functional Structure" Agronomy 15, no. 6: 1354. https://doi.org/10.3390/agronomy15061354
APA StyleMorrison, J., Izquierdo, J., Hernández Plaza, E., & González-Andújar, J. L. (2025). How the Functional Constitution of Plant Communities in Field Margins Affects Wild Bee Community Composition and Functional Structure. Agronomy, 15(6), 1354. https://doi.org/10.3390/agronomy15061354