
Sodium Chloride Enhances Nitrogen Use Efficiency but Reduces Yield Benefits Under Elevated CO2 in Upland Rice

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions
2.2. Treatments and Experimental Design
2.3. Evaluated Traits and Measurements
- Vcmax: Maximum carboxylation rate of RuBisCO, reflecting enzyme activity capacity;
- J: Rate of photosynthetic electron transport, indicating the efficiency of light-dependent reactions;
- LCP (light compensation point): The light intensity at which photosynthesis equals respiration;
- Amax: Maximum photosynthetic rate under light saturation, indicating photosynthetic potential;
2.4. Statistical Analysis
3. Results
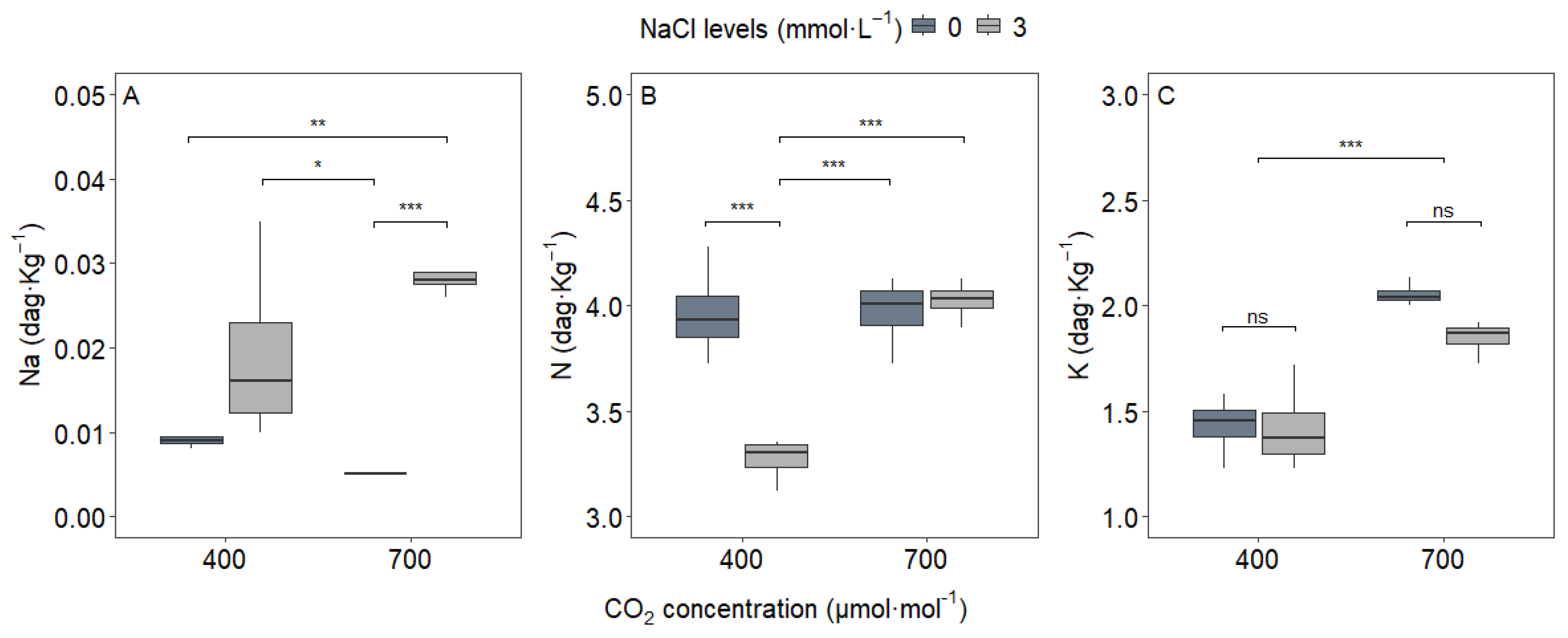
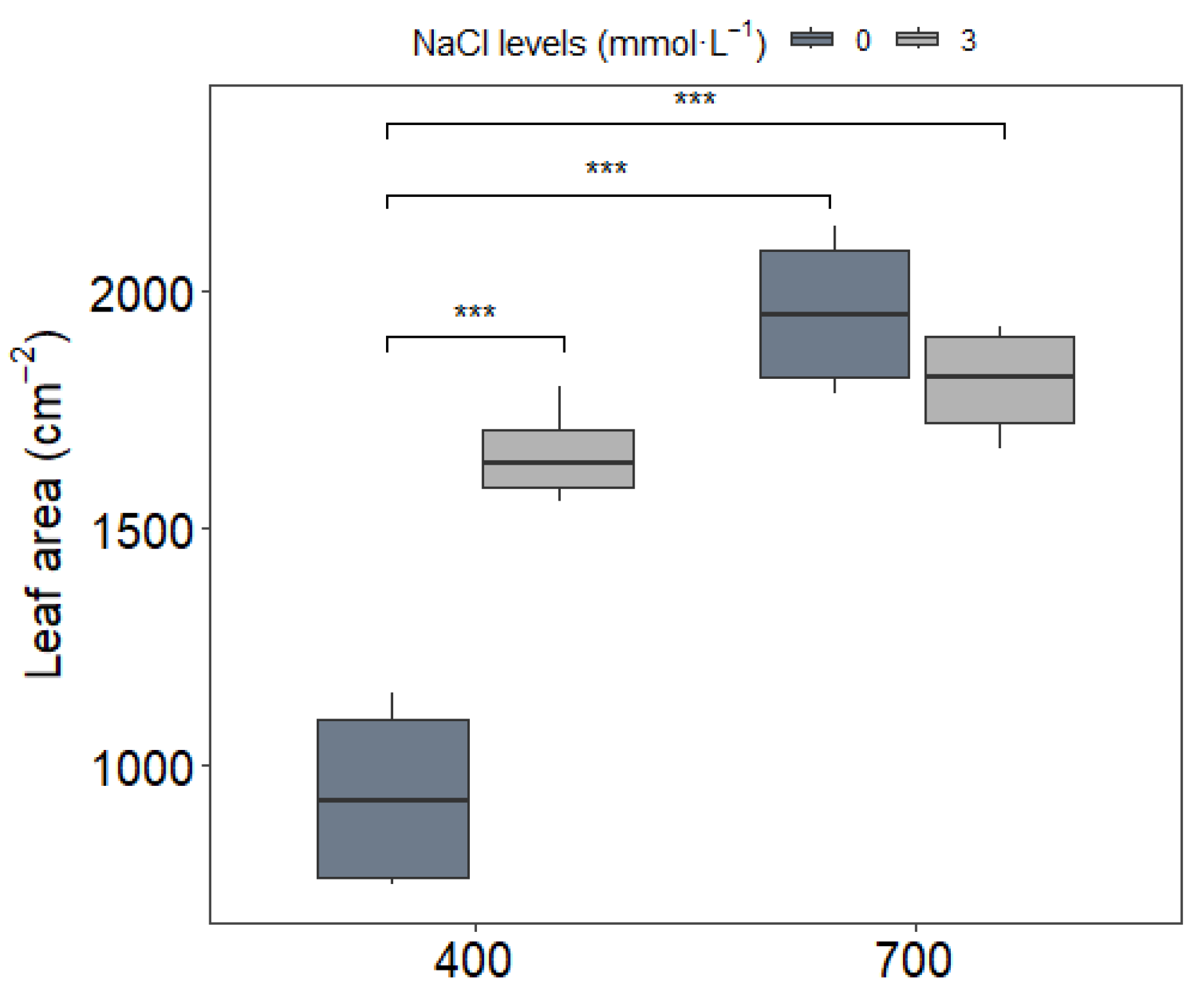
3.1. Leaf Nutrient Content and Leaf Area
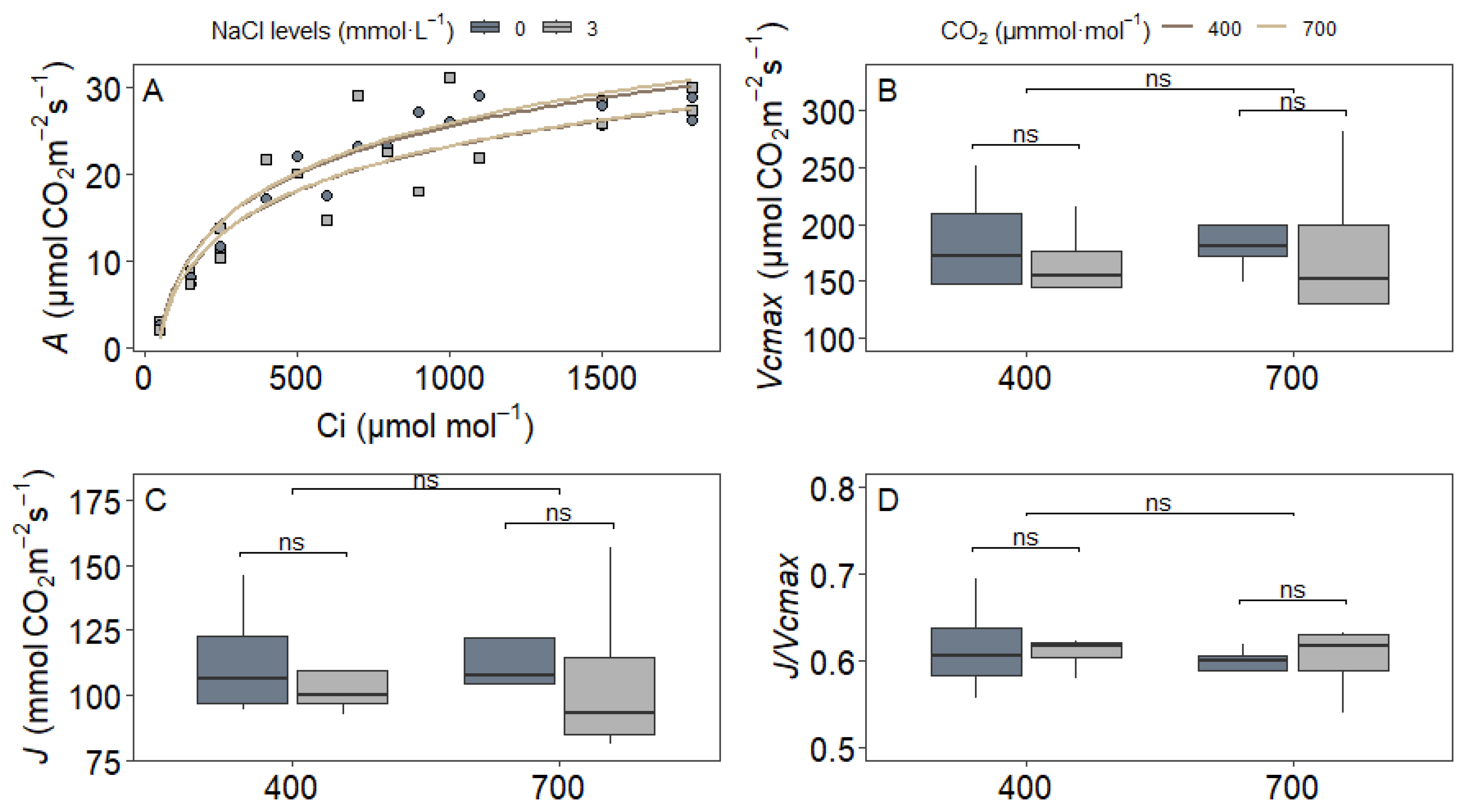
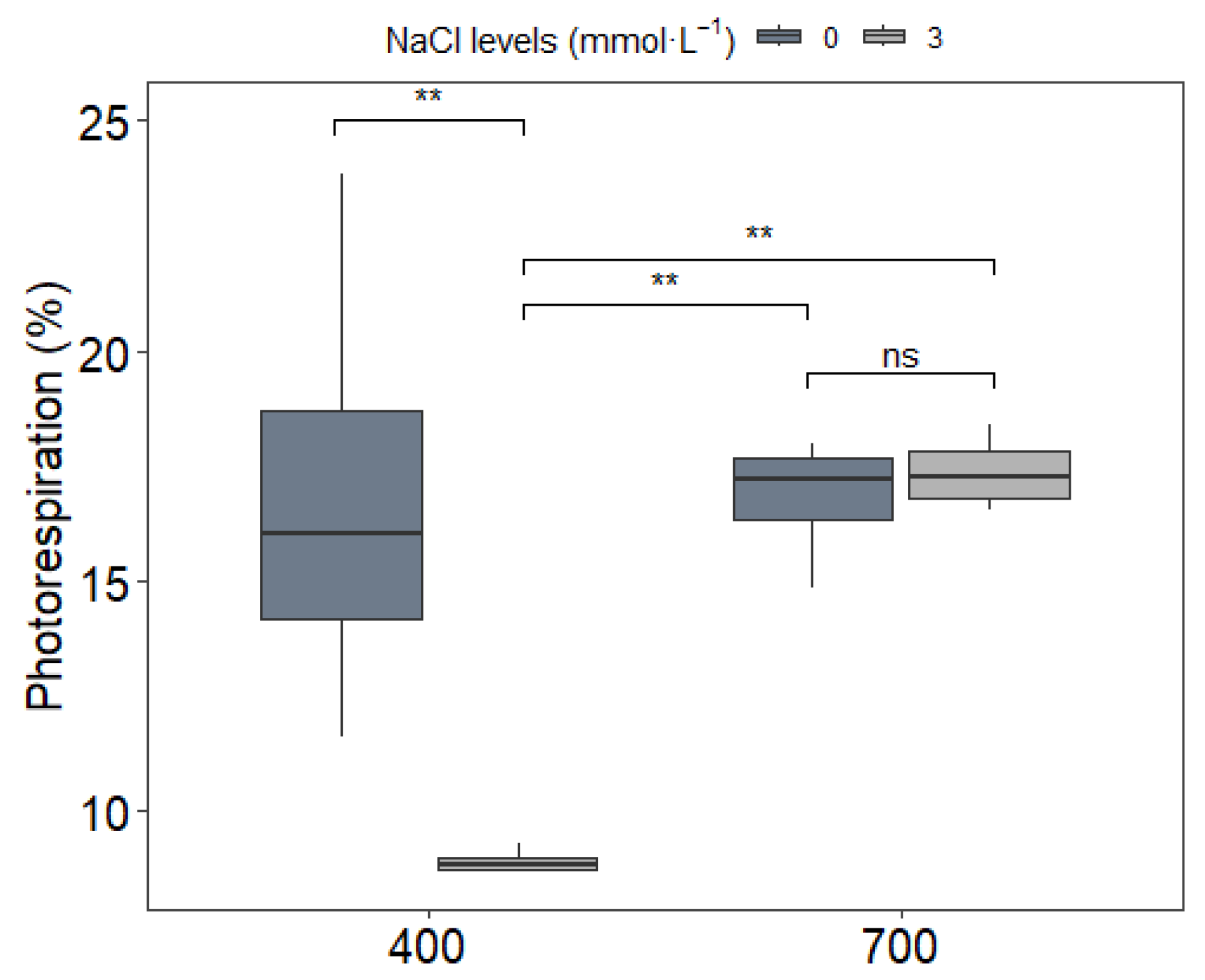
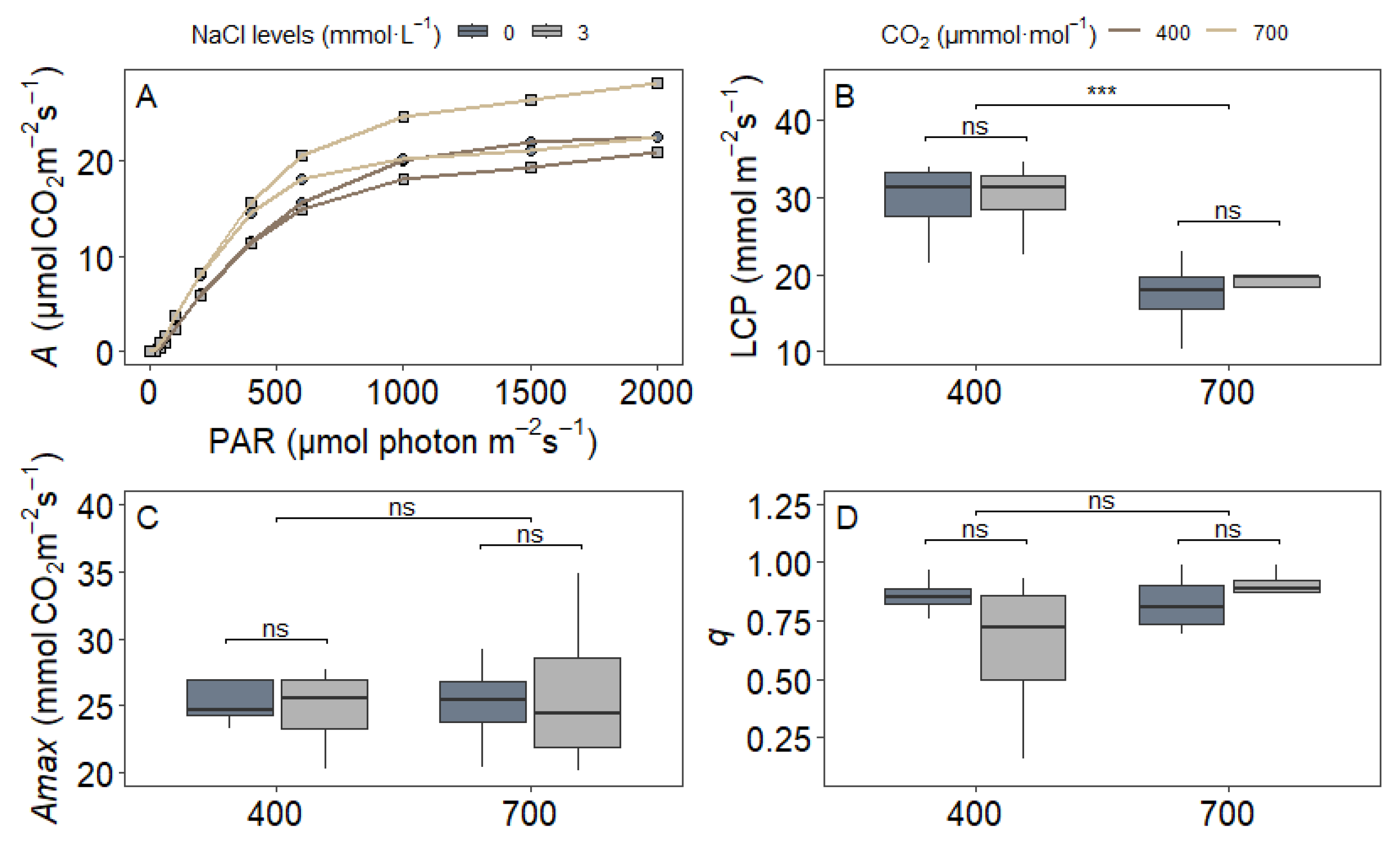
3.2. Gas Exchange Parameters
3.3. Rice Yield and Photosystem Efficiency
3.4. Nitrogen Use Efficiency—NUE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Strategy on Climate Change 2022–2031; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- Eisenhut, M.; Weber, A.P.M. Improving Crop Yield. Science 2019, 363, 32–33. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.P.R.A.D.; Cruz, A.B.; dos Santos Botelho, A.; Pennacchi, J.P.; Santana, G.F. Crop Physiology, the Technology and the Production Gap. Theor. Exp. Plant Physiol. 2024, 36, 567–582. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.S.; Xia, J.Q.; Alfatih, A.; Song, Y.; Huang, Y.J.; Sun, L.Q.; Wan, G.Y.; Wang, S.M.; Wang, Y.P.; Hu, B.H.; et al. Rice NIN-LIKE PROTEIN 3 modulates nitrogen use efficiency and grain yield under nitrate-sufficient conditions. Plant Cell Environ. 2022, 45, 1520–1536. [Google Scholar] [CrossRef] [PubMed]
- Bauwe, H. Humboldt Review: Photorespiration—Rubisco’s repair crew. J. Plant Physiol. 2023, 280, 153899. [Google Scholar] [CrossRef]
- Yeo, M.E.; Yeo, A.R.; Flowers, T.J. Photosynthesis and Photorespiration in the Genus Oryza. J. Exp. Bot. 1994, 45, 553–560. [Google Scholar] [CrossRef]
- Dahal, K.; Knowles, V.L.; Plaxton, W.C.; Hüner, N.P.A. Enhancement of Photosynthetic Performance, Water Use Efficiency and Grain Yield During Long-Term Growth Under Elevated CO2 in Wheat and Rye Is Growth Temperature and Cultivar Dependent. Environ. Exp. Bot. 2014, 106, 207–220. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Milo, R. The Global Mass and Average Rate of Rubisco. Proc. Natl. Acad. Sci. USA 2019, 116, 4738–4743. [Google Scholar] [CrossRef]
- Hunt, S. Measurements of Photosynthesis and Respiration in Plants. Physiol. Plant. 2003, 117, 314–325. [Google Scholar] [CrossRef]
- Wang, W.; Cai, C.; He, J.; Gu, J.; Zhu, G.; Zhang, W.; Liu, G. Yield, Dry Matter Distribution and Photosynthetic Characteristics of Rice Under Elevated CO2 and Increased Temperature Conditions. Field Crops Res. 2020, 248, 107605. [Google Scholar] [CrossRef]
- Kazemi, S.; Eshghizadeh, H.R.; Zahedi, M. Responses of Four Rice Varieties to Elevated CO2 and Different Salinity Levels. Rice Sci. 2018, 25, 142–151. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Wheeler, R.M.; Levine, L.H.; Stutte, G.W. Low Concentrations of Sodium Stimulate Specific Physiological Responses in Leaf Lettuce (Lactuca sativa L.). Plant Cell Environ. 2003, 26, 967–975. [Google Scholar] [CrossRef]
- Marschner, P. (Ed.) Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar] [CrossRef]
- Hongqiao, L.; Suyama, A.; Mitani-Ueno, N.; Hell, R.; Maruyama-Nakashita, A. A Low Level of NaCl Stimulates Plant Growth by Improving Carbon and Sulfur Assimilation in Arabidopsis thaliana. Plants 2021, 10, 2138. [Google Scholar] [CrossRef] [PubMed]
- Piccolo, E.L.; Ceccanti, C.; Guidi, L.; Landi, M. Role of beneficial elements in plants: Implications for the photosynthetic process. Photosynthetica 2021, 59, 349–360. [Google Scholar] [CrossRef]
- Vieira, D.A.; Toro-Herrera, M.A.; Pennacchi, J.P.; Mendonça, A.M.C.; Marchiori, P.E.R.; Botelho, F.B.S.; Barbosa, J.P.R.A.D. Mapping Phenotypic Parameters Linked to Salt Stress Tolerance Strategies in Rice Lines. Bragantia 2022, 81, e20210172. [Google Scholar] [CrossRef]
- Vieira, D.A. Agronomic and Physiological Responses of Rice to Sodium Treatments and High CO2. Ph.D. Thesis, Federal University of Lavras (UFLA), Lavras, Brazil, 2021. [Google Scholar]
- Alvarez, R.C.F.; Crusciol, C.A.C.; Nascente, A.S. Análise de crescimento e produtividade de cultivares de arroz de terras altas dos tipos tradicional, intermediário e moderno. Pesq. Agropec. Trop. 2012, 42, 397–406. [Google Scholar] [CrossRef]
- Mohapatra, P.K.; Sahu, B.B. Botany of Rice Plant. In Panicle Architecture of Rice and Its Relationship with Grain Filling; Springer: Cham, Switzerland, 2022; pp. 27–48. [Google Scholar] [CrossRef]
- Counce, P.A.; Keisling, T.C.; Mitchell, A.J. A Uniform, Adaptative System for Expressing Rice Development. Crop Sci. 2000, 40, 436–443. [Google Scholar] [CrossRef]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated Digital Image Analysis for Rapid and Accurate Measurement of Leaf Area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef]
- Arantes, M.K.; da Silva Filho, M.P.; Pennacchi, J.P.; das Chagas Mendonca, A.M.; Barbosa, J.P. Phenotypic Plasticity of Leaf Anatomical Traits Helps to Explain Gas-Exchange Response to Water Shortage in Grasses of Different Photosynthetic Types. Theor. Exp. Plant Physiol. 2020, 32, 341–356. [Google Scholar] [CrossRef]
- Marshall, B.; Biscoe, P.V. A Model for C3 Leaves Describing the Dependence of Net Photosynthesis on Irradiance: II. Application to the Analysis of Flag Leaf Photosynthesis. J. Exp. Bot. 1980, 31, 41–48. [Google Scholar] [CrossRef]
- Thornley, J.H.M.; Johnson, I.R. Plant and Crop Modelling: A Mathematical Approach to Plant and Crop Physiology; Oxford University Press: Oxford, UK, 1990. [Google Scholar]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research, R package version 1.3-6; 2020. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 12 May 2025).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 12 May 2025).
- Hope, R.M. Rmisc: R Miscellaneous Functions, R package version 1.5; 2022. Available online: https://CRAN.R-project.org/package=Rmisc (accessed on 12 May 2025).
- Kassambara, A. Ggpubr: ‘ggplot2’ Based Publication Ready Plots, R package version 0.6.0; 2023. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 12 May 2025).
- Reddy, I.N.B.L.; Kim, B.K.; Yoon, I.S.; Kim, K.H.; Kwon, T.R. Salt Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Al Shiblawi, F.R.; Sentenac, H. Roles and Transport of Sodium and Potassium in Plants. In The Alkali Metal Ions: Their Role for Life; Sigel, A., Sigel, H., Sigel, R., Eds.; Springer: Cham, Switzerland, 2016; Volume 16. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Ou, Y.; Wang, L.; Gao, Y.; Lucas, G.R.; Resco de Dios, V.; Yao, Y. Transcriptome and low-affinity sodium transport analysis reveals salt tolerance variations between two poplars. Int. J. Mol. Sci. 2023, 24, 5732. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Liu, M.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Nie, C.; Yu, M.; Shabala, S. Non-stomatal limitation of photosynthesis by soil salinity. Crit. Rev. Environ. Sci. Technol. 2020, 51, 791–825. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, D.A.; Toro-Herrera, M.A.; Pennacchi, J.P.; Carvalho, M.M.P.; Botelho, F.B.S.; Marchiori, P.E.R.; Barbosa, J.P.R.A.D. Sodium Chloride Enhances Nitrogen Use Efficiency but Reduces Yield Benefits Under Elevated CO2 in Upland Rice. Agronomy 2025, 15, 1212. https://doi.org/10.3390/agronomy15051212
Vieira DA, Toro-Herrera MA, Pennacchi JP, Carvalho MMP, Botelho FBS, Marchiori PER, Barbosa JPRAD. Sodium Chloride Enhances Nitrogen Use Efficiency but Reduces Yield Benefits Under Elevated CO2 in Upland Rice. Agronomy. 2025; 15(5):1212. https://doi.org/10.3390/agronomy15051212
Chicago/Turabian StyleVieira, Daniel Amorim, Mayra Alejandra Toro-Herrera, João Paulo Pennacchi, Marília Mickaele Pinheiro Carvalho, Flavia Barbosa Silva Botelho, Paulo Eduardo Ribeiro Marchiori, and João Paulo Rodrigues Alves Delfino Barbosa. 2025. "Sodium Chloride Enhances Nitrogen Use Efficiency but Reduces Yield Benefits Under Elevated CO2 in Upland Rice" Agronomy 15, no. 5: 1212. https://doi.org/10.3390/agronomy15051212
APA StyleVieira, D. A., Toro-Herrera, M. A., Pennacchi, J. P., Carvalho, M. M. P., Botelho, F. B. S., Marchiori, P. E. R., & Barbosa, J. P. R. A. D. (2025). Sodium Chloride Enhances Nitrogen Use Efficiency but Reduces Yield Benefits Under Elevated CO2 in Upland Rice. Agronomy, 15(5), 1212. https://doi.org/10.3390/agronomy15051212