
Hydro-Electro Hybrid Priming Synchronizes Cell Wall Remodeling to Accelerate Carrot (Daucus carota L.) Seed Germination

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Material and Priming Treatments
2.2. Germination Assay and Determination of Mechanical Properties of Endosperm
2.3. Transcriptome Sequencing and Analysis
2.3.1. RNA Extraction and Library Construction
2.3.2. Sequencing and Data Preprocessing
2.3.3. Sequence Alignment and Differential Expression Analysis
2.3.4. Enrichment Analysis of DEGs
2.4. Activity Determination of Key Enzymes in Cell Wall Remodeling Pathways
2.5. qRT-PCR Analysis of Key DEGs in Cell Wall Remodeling Pathways
2.6. Statistical Analysis
3. Results
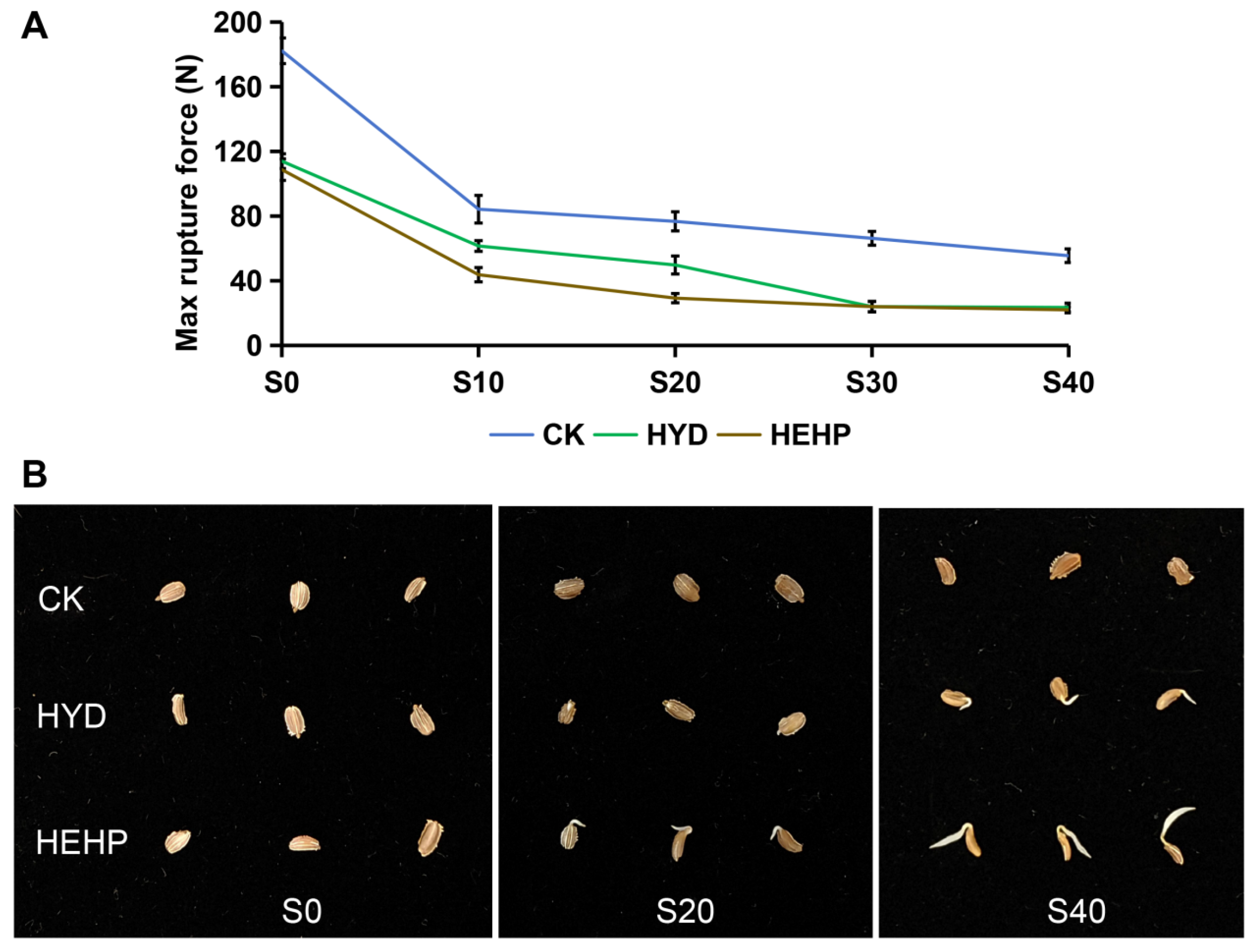
3.1. Differences in Endosperm Mechanical Characteristics of Carrot Seeds
3.2. Statistics of DEGs and Comparison with the Number of DEGs Sampled Before Imbibition
3.3. GO and KEGG Enrichment Analysis of DEGs
3.4. DEGs Involved in Cell Wall Remodeling Pathways
3.5. Analysis of Key Enzyme Activities in Cell Wall Remodeling Pathways
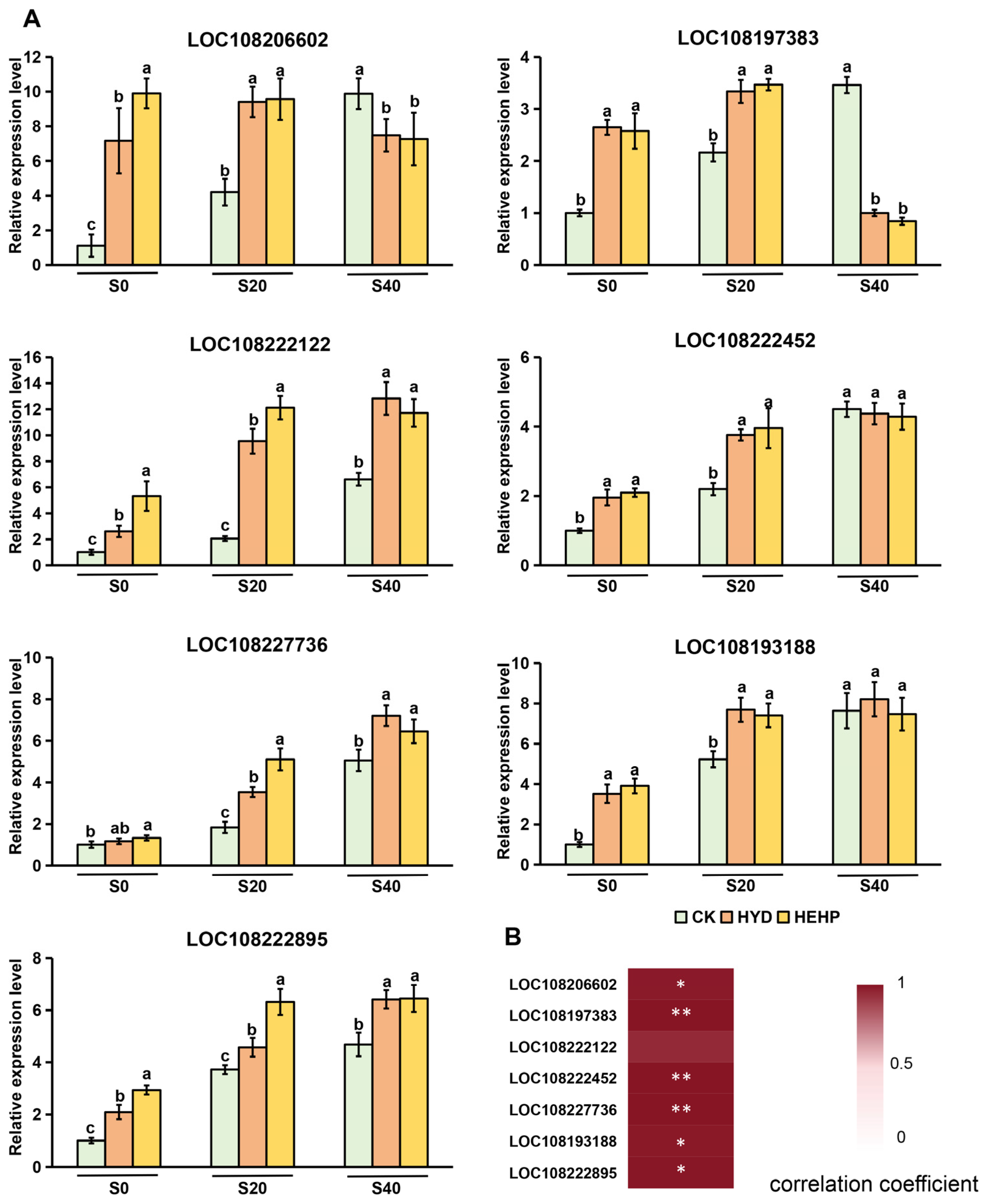
3.6. Analysis of Expression Patterns of Key Genes in Cell Wall Remodeling Pathways
4. Discussion
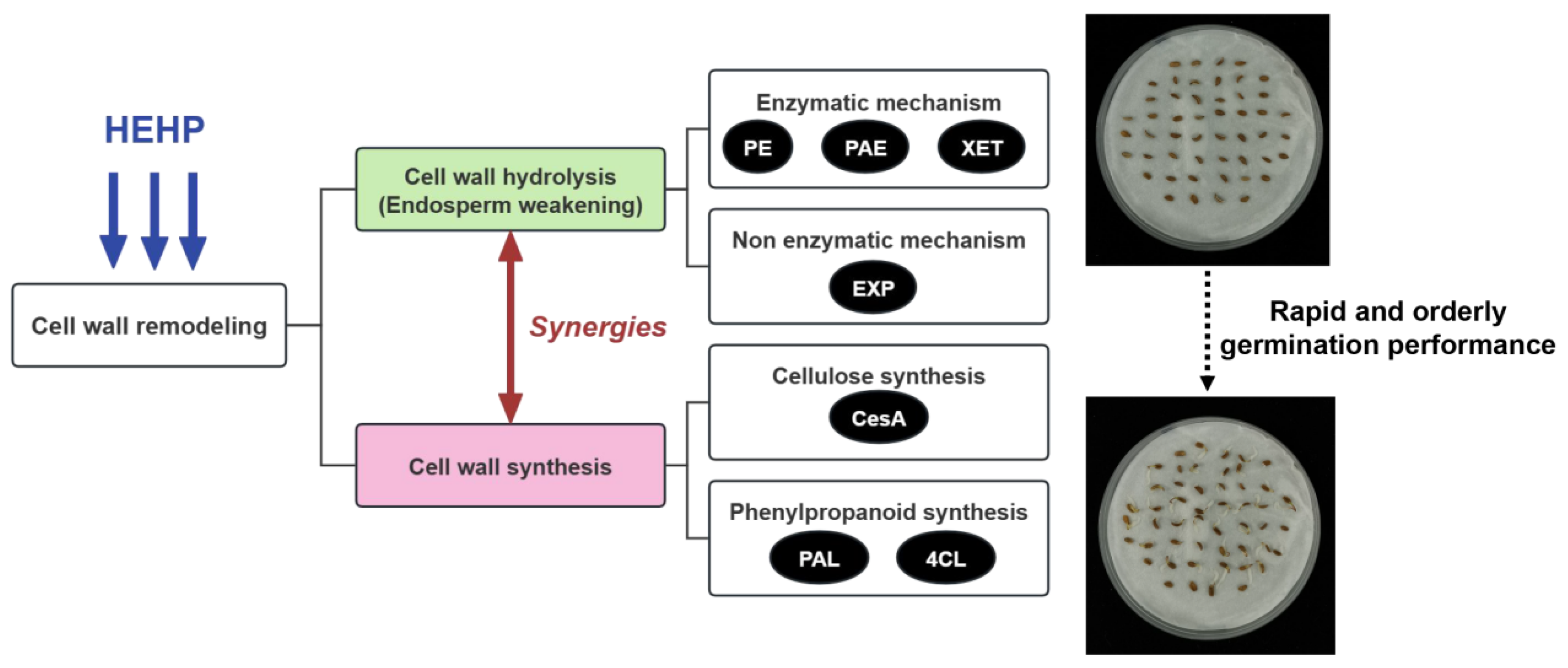
4.1. HEHP Synergistically Coordinates Cell Wall Decomposition and Synthesis for Enhanced Endosperm Weakening
4.2. Phenylpropanoid Biosynthesis as a Pivotal Link Between Priming and Cell Wall Reinforcement
4.3. Temporal Dynamics of Cell Wall Remodeling Underpin Germination Synchronization
4.4. Redox Signals May Serve as a Bridge Between Electrostatic Fields and Metabolic Reprogramming
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jhanji, S.; Goyal, E.; Chumber, M.; Kaur, G. Exploring fine tuning between phytohormones and ROS signaling cascade in regulation of seed dormancy, germination and seedling development. Plant Physiol. Biochem. 2024, 207, 108352. [Google Scholar] [CrossRef] [PubMed]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [PubMed]
- Luby, C.H.; Maeda, H.A.; Goldman, I.L. Genetic and phenological variation of tocochromanol (vitamin E) content in wild (Daucus carota L. var. carota) and domesticated carrot (D. carota L. var. sativa). Hortic. Res. 2014, 1, 14015. [Google Scholar] [CrossRef]
- Que, F.; Hou, X.L.; Wang, G.L.; Xu, Z.S.; Xiong, A.S. Advances in research on the carrot, an important root vegetable in the Apiaceae family. Hortic. Res. 2019, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I. Seed priming: What’s next? Seed Sci. Technol. 2023, 51, 379–405. [Google Scholar] [CrossRef]
- Choi, J.Y.; Ju, Y.H.; Nakamichi, A.; Cho, S.W.; Woo, S.H.; Sakagami, J.I. Effect of seed hydropriming on the elongation of plumule and radicle during the germination process and changes in enzyme activity under water-deficient conditions. Plants 2024, 13, 3537. [Google Scholar] [CrossRef]
- Jiang, B.W.; Wang, L.Y.; Xu, C.T.; Yan, M. Hydropriming enhances the germination of aged ultra-dry wheat seeds. Seed Sci. Technol. 2020, 48, 57–63. [Google Scholar] [CrossRef]
- Thakur, M.; Tiwari, S.; Kataria, S.; Anand, A. Recent advances in seed priming strategies for enhancing planting value of vegetable seeds. Sci. Hortic. 2022, 305, 111355. [Google Scholar] [CrossRef]
- Aasim, M.; Akin, F.; Ali, S.A. Synergizing LED Technology and Hydropriming for Intelligent Modeling and Mathematical Expressions to Optimize Chickpea Germination and Growth Indices. J. Plant Growth Regul. 2024, 43, 2340–2359. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Hu, M.H.; Gao, Z.; Chen, X.X.; Huang, D.F. Biological mechanisms of a novel hydro-electro hybrid priming recovers potential vigor of onion seeds. Environ. Exp. Bot. 2018, 150, 260–271. [Google Scholar] [CrossRef]
- Adhikari, B.; Adhikari, M.; Ghimire, B.; Adhikari, B.C.; Park, G.; Choi, E.H. Cold plasma seed priming modulates growth, redox homeostasis and stress response by inducing reactive species in tomato (Solanum lycopersicum). Free Radic. Biol. Med. 2020, 156, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Samarah, N.H.; Hani, M.; Makhadmeh, I.M. Effect of magnetic treatment of water or seeds on germination and productivity of tomato plants under salinity stress. Horticulturae 2021, 7, 220. [Google Scholar] [CrossRef]
- Ries, A.; Benitez, J.V.; Samudio, A.; Armoa, R.; Nakayama, H.D. Germination of bean seeds (Vigna unguiculata L. Walp.) in strong electric fields. MethodsX 2023, 11, 102490. [Google Scholar] [CrossRef]
- Mamlic, Z.; Maksimovic, I.; Canak, P.; Mamlic, G.; Djukic, V.; Vasiljevic, S.; Dozet, G. The use of electrostatic field to improve soybean seed germination in organic production. Agronomy 2021, 11, 1473. [Google Scholar] [CrossRef]
- Wang, G.; Huang, J.; Gao, W.; Lu, J.; Li, J.; Liao, R.; Jaleel, C.A. The effect of high-voltage electrostatic field (HVEF) on aged rice (Oryza sativa L.) seeds vigor and lipid peroxidation of seedlings. J. Electrost. 2009, 67, 759–764. [Google Scholar] [CrossRef]
- Zhao, S.; Garcia, D.; Zhao, Y.; Huang, D. Hydro-electro hybrid priming promotes carrot (Daucus carota L.) seed germination by activating lipid utilization and respiratory metabolism. Int. J. Mol. Sci. 2021, 22, 11090. [Google Scholar] [CrossRef]
- Zhao, S.; Zou, H.; Jia, Y.; Pan, X.; Huang, D. Carrot (Daucus carota L.) seed germination was promoted by hydro-electro hybrid priming through regulating the accumulation of proteins involved in carbohydrate and protein metabolism. Front. Plant Sci. 2022, 13, 824439. [Google Scholar] [CrossRef]
- Shigeyama, T.; Watanabe, A.; Tokuchi, K.; Toh, S.; Sakurai, N.; Shibuya, N.; Kawakami, N. α-Xylosidase plays essential roles in xyloglucan remodelling, maintenance of cell wall integrity, and seed germination in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 5615–5629. [Google Scholar] [CrossRef]
- Zhang, J.L.; Liu, L.; Dong, D.H.; Xu, J.Y.; Li, H.X.; Deng, Q.L.; Zhang, Y.; Huang, W.; Zhang, H.J.; Guo, Y.D. The transcription factor SlLBD40 regulates seed germination by inhibiting cell wall remodeling enzymes during endosperm weakening. Plant Physiol. 2025, 197, kiaf022. [Google Scholar] [CrossRef]
- Jemmat, A.M.; Ranocha, P.; Le Ru, A.; Neel, M.; Jauneau, A.; Raggi, S.; Ferrari, S.; Burlat, V.; Dunand, C. Coordination of five class III peroxidase-encoding genes for early germination events of Arabidopsis thaliana. Plant Sci. 2020, 298, 110565. [Google Scholar] [CrossRef]
- Chandrasekaran, U.; Zhao, X.; Luo, X.; Wei, S.; Shu, K. Endosperm weakening: The gateway to a seed’s new life. Plant Physiol. Biochem. 2022, 178, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Han, X.; Yin, Y.L.; Wang, G.B.; Huang, D.F.; Lan, Y.B. Enzymatic loosening mechanism of endosperm weakening plays a key role in promoting carrot (Daucus carota L.) seed germination by hydro-electro hybrid priming. Sci. Hortic. 2024, 333, 113255. [Google Scholar] [CrossRef]
- Cao, X.L.; Li, M.L.; Li, J.; Song, Y.X.; Zhang, X.N.; Yang, D.L.; Li, M.F.; Wei, J.H. Co-expression of hydrolase genes improves seed germination of Sinopodophyllum hexandrum. Ind. Crops Prod. 2021, 164, 113414. [Google Scholar] [CrossRef]
- Muthusamy, M.; Kim, J.Y.; Yoon, E.K.; Kim, J.A.; Lee, S.I. BrEXLB1, a Brassica rapa Expansin-Like B1 Gene Is Associated with Root Development, Drought Stress Response, and Seed Germination. Genes 2020, 11, 404. [Google Scholar] [CrossRef]
- Chen, S.K.; Luo, Y.X.; Wang, G.J.; Feng, C.Z.; Li, H.F. Genome-wide identification of expansin genes in Brachypodium distachyon and functional characterization of BdEXPA27. Plant Sci. 2020, 296, 110490. [Google Scholar] [CrossRef]
- Lin, F.; Manisseri, C.; Fagerström, A.; Peck, M.L.; Vega-Sánchez, M.E.; Williams, B.; Chiniquy, D.M.; Saha, P.; Pattathil, S.; Conlin, B.; et al. Cell wall composition and candidate biosynthesis gene expression during rice development. Plant Cell Physiol. 2016, 57, 2058–2075. [Google Scholar] [CrossRef]
- Zhang, Q.; Qin, B.; Wang, G.D.; Zhang, W.J.; Li, M.; Yin, Z.G.; Yuan, X.K.; Sun, H.Y.; Du, J.D.; Du, Y.L.; et al. Exogenous melatonin enhances cell wall response to salt stress in common bean (Phaseolus vulgaris) and the development of the associated predictive molecular markers. Front. Plant Sci. 2022, 13, 1012186. [Google Scholar] [CrossRef]
- Tong, Y.; Yi, S.C.; Liu, S.Y.; Xu, L.; Qiu, Z.X.; Zeng, D.Q.; Tang, W.W. Bruceine D may affect the phenylpropanoid biosynthesis by acting on ADTs thus inhibiting Bidens pilosa L. seed germination. Ecotoxicol. Environ. Saf. 2022, 242, 113943. [Google Scholar] [CrossRef]
- Yu, A.L.; Zhao, J.F.; Wang, Z.H.; Cheng, K.; Zhang, P.; Tian, G.; Liu, X.; Guo, E.H.; Du, Y.W.; Wang, Y.W. Transcriptome and metabolite analysis reveal the drought tolerance of foxtail millet significantly correlated with phenylpropanoids-related pathways during germination process under PEG stress. BMC Plant Biol. 2020, 20, 274. [Google Scholar] [CrossRef]
- Alibas, I.; Koksal, N. The effect of moisture content on physical, mechanical and rheological properties of soybean (Glycine max cv. ATAEM-II) seed. Legume Res. 2015, 38, 324–333. [Google Scholar] [CrossRef]
- Grsic-Rausch, S.; Rausch, T. A coupled spectrophotometric enzyme assay for the determination of pectin methylesterase activity and its inhibition by proteinaceous inhibitors. Anal. Biochem. 2004, 333, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.Y.; Huang, Y.H.; Lin, Z.Y.; Hsieh, L.S. Insights into the substrate selectivity of Bambusa oldhamii phenylalanine ammonia-lyase 1 and 2 through mutational analysis. Phytochem. Lett. 2020, 38, 140–143. [Google Scholar] [CrossRef]
- Wu, Z.C.; Cui, C.L.; Xing, A.Q.; Xu, X.H.; Sun, Y.; Tian, Z.Q.; Li, X.Y.; Zhu, J.Y.; Wang, G.M.; Wang, Y.H. Identification and response analysis of xyloglucan endotransglycosylase/hydrolases (XTH) family to fluoride and aluminum treatment in Camellia sinensis. BMC Genom. 2021, 22, 761. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Sun, S.-J.; Horikawa, Y.; Wada, M.; Sugiyama, J. Functional reconstitution of cellulose synthase in Escherichia coli. Biomacromolecules 2014, 15, 4206–4213. [Google Scholar] [CrossRef]
- Duan, Y.H.; Ma, Y.Y.; Zhao, X.D.; Huang, R.L.; Su, R.X.; Qi, W.; He, Z.M. Real-time adsorption and action of expansin on cellulose. Biotechnol. Biofuels 2018, 11, 317. [Google Scholar] [CrossRef]
- Montechiarini, N.H.; Delgado, L.; Morandi, E.N.; Carrillo, N.J.; Gosparini, C.O. The expansin EXP1 gene in the elongation zone is induced during soybean embryonic axis germination and differentially expressed in response to ABA and PEG treatments. Seed Sci. Res. 2021, 31, 60–68. [Google Scholar] [CrossRef]
- Zhu, J.Q.; Tang, G.Y.; Xu, P.L.; Li, G.W.; Ma, C.L.; Li, P.X.; Jiang, C.Y.; Shan, L.; Wan, S.B. Genome-wide identification of xyloglucan endotransglucosylase/hydrolase gene family members in peanut and their expression profiles during seed germination. PeerJ 2022, 10, e13428. [Google Scholar] [CrossRef]
- Ducatti, K.R.; Batista, T.B.; Hirai, W.Y.; Luccas, D.A.; Moreno, L.D.; Guimaraes, C.C.; Bassel, G.W.; da Silva, E.A.A. Transcripts expressed during germination sensu stricto are associated with vigor in soybean seeds. Plants 2022, 11, 1310. [Google Scholar] [CrossRef]
- Liu, H.N.; Pei, M.S.; Ampomah-Dwamena, C.; Shang, Y.X.; Yu, Y.H.; Wei, T.L.; Shi, Q.F.; Guo, D.L. Alternative splicing of the PECTINESTERASE gene encoding a cell wall-degrading enzyme affects postharvest softening in grape. J. Integr. Agric. 2024, 23, 863–875. [Google Scholar] [CrossRef]
- Ishida, K.; Yokoyama, R. Reconsidering the function of the xyloglucan endotransglucosylase/hydrolase family. J. Plant Res. 2022, 135, 145–156. [Google Scholar] [CrossRef]
- Zhao, J.; He, Y.Q.; Li, X.Y.; Weng, X.A.; Feng, D.F.; Ying, J.F.; Wang, Z.F. An integrated RNA-Seq and physiological study reveals gene responses involving in the initial imbibition of seed germination in rice. Plant Growth Regul. 2020, 90, 249–263. [Google Scholar] [CrossRef]
- Pourbagher, M.; Pourbagher, R.; Abbaspour-Fard, M.H. Reduction of Adverse Effects of Drought Stress on Germination Indices and Antioxidant Enzymes of Licorice Seeds (Glycyrrhiza) Using Cold Plasma. J. Plant Growth Regul. 2025, 44, 1032–1042. [Google Scholar] [CrossRef]
- Barros, J.; Serrani-Yarce, J.C.; Chen, F.; Baxter, D.; Venables, B.J.; Dixon, R.A. Role of bifunctional ammonia-lyase in grass cell wall biosynthesis. Nat. Plants 2016, 2, 16050. [Google Scholar] [CrossRef]
- Fang, L.; Xu, X.; Li, J.; Zheng, F.; Li, M.Z.; Yan, J.W.; Li, Y.; Zhang, X.H.; Li, L.; Ma, G.H.; et al. Transcriptome analysis provides insights into the non-methylated lignin synthesis in Paphiopedilum armeniacum seed. BMC Genom. 2020, 21, 524. [Google Scholar] [CrossRef]
- Tong, Y.; Liu, S.Y.; Yi, S.C.; Qiu, Z.X.; Wang, Y.H.; Zeng, D.Q.; Tang, W.W. Bruceine D, the main active ingredient of Brucea javanica (L.) residue inhibits the germination of Bidens pilosa L. seeds by suppressing phenylpropanoid biosynthesis. Ind. Crops Prod. 2021, 172, 114079. [Google Scholar] [CrossRef]
- Seyfferth, C.; Wessels, B.A.; Vahala, J.; Kangasjärvi, J.; Delhomme, N.; Hvidsten, T.R.; Tuominen, H.; Lundberg-Felten, J. Populus PtERF85 Balances Xylem Cell Expansion and Secondary Cell Wall Formation in Hybrid Aspen. Cells 2021, 10, 1971. [Google Scholar] [CrossRef]
- Peng, C.Y.; Wu, Y.; Hua, Q.L.; Shen, Y.B. Hydrological transport and endosperm weakening mechanisms during dormancy release in Tilia henryana seeds. J. Plant Physiol. 2025, 304, 154405. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Niu, Y.N.; Bai, X.D.; Mao, T.T. Transcriptomic and metabolic profiling reveals a lignin metabolism network involved in mesocotyl elongation during maize seed germination. Plants 2022, 11, 1034. [Google Scholar] [CrossRef]
- Zhang, W.H.; Wang, G.H.; Zhang, B.; Sui, W.J.; Si, C.L.; Zhou, L.P.; Jia, H.Y. Green potassium fertilizer from enzymatic hydrolysis lignin: Effects of lignin fractionation on wheat seed germination and seedling growth. Int. J. Biol. Macromol. 2024, 262, 130017. [Google Scholar] [CrossRef]
- Qi, M.Y.; Liu, Y.; Shi, S.S.; Xian, Y.H.; Liu, Q.Y.; Yan, H.Y.; Zhang, Y.; Yuan, Y. Inhibition mechanism of high voltage prick electrostatic field (HVPEF) on Staphylococcus aureus through ROS-mediated oxidative stress. Lwt-Food Sci. Technol. 2022, 155, 112990. [Google Scholar] [CrossRef]
- Liu, X.; Quan, W.L.; Bartels, D. Stress memory responses and seed priming correlate with drought tolerance in plants: An overview. Planta 2022, 255, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Zheng, X.Y.; Tao, Y.; Xie, C.; Li, D.D.; Han, Y.B. Moderate electric field-stimulated brown rice germination: Insights into membrane permeability modulation and antioxidant system activation. Food Chem. 2025, 479, 143737. [Google Scholar] [CrossRef] [PubMed]
- Billah, M.; Karmakar, S.; Mina, F.B.; Haque, M.N.; Rashid, M.M.; Hasan, M.F.; Acharjee, U.K.; Talukder, M.R. Investigation of mechanisms involved in seed germination enhancement, enzymatic activity and seedling growth of rice (Oryza sativa L.) using LPDBD (Ar plus Air) plasma. Arch. Biochem. Biophys. 2021, 698, 108726. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Hu, P.G.; Li, F.J.; Wu, H.H.; Shen, Y.; White, J.C.; Tian, X.L.; Li, Z.H.; Giraldo, J.P. Emerging investigator series: Molecular mechanisms of plant salinity stress tolerance improvement by seed priming with cerium oxide nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Kato, M.; Ma, G.; Zhang, L.C.; Uthairatanakij, A.; Srilaong, V.; Laohakunjit, N.; Jitareerat, P. Electron beam radiation delayed the disassembly of cell wall polysaccharides in harvested mangoes. Postharvest Biol. Technol. 2021, 178, 111544. [Google Scholar] [CrossRef]
- Moles, T.M.; Guglielminetti, L.; Reyes, T.H. Differential effects of sodium chloride on germination and post-germination stages of two tomato genotypes. Sci. Hortic. 2019, 257, 108730. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Total DEGs | Upregulated | Downregulated |
|---|---|---|---|
| CK vs. HYD | 3282 | 2341 | 941 |
| CK vs. HEHP | 4591 | 3168 | 1423 |
| HYD vs. HEHP | 90 | 58 | 32 |
| CK0 vs. CK | 11,095 | 6520 | 4575 |
| HYD0 vs. HYD | 8175 | 5371 | 2804 |
| HEHP0 vs. HEHP | 4069 | 2596 | 1473 |
| Coding Protein | Gene ID | Gene Expression Difference Multiple (log2FC) | ||
|---|---|---|---|---|
| HEHP/CK | HYD/CK | HEHP/HYD | ||
| EXP | LOC108214289 | −1.49 | −0.37 | −1.13 |
| LOC108206602 | 6.19 | 6.46 | −0.27 | |
| LOC108225470 | 2.79 | 2.36 | 0.43 | |
| LOC108195513 | 1.22 | 0.95 | 0.27 | |
| LOC108218688 | 0.98 | 0.28 | 0.71 | |
| LOC108213182 | 1.36 | 0.81 | 0.55 | |
| LOC108196118 | −1.20 | −0.05 | −1.14 | |
| LOC108205620 | 3.63 | 2.69 | 0.93 | |
| LOC108209261 | 0.83 | 1.14 | −0.31 | |
| XET | LOC108217609 | 1.01 | 1.12 | −0.11 |
| LOC108217607 | 1.73 | 1.54 | 0.19 | |
| LOC108200367 | 1.88 | 1.76 | 0.12 | |
| LOC108197383 | 4.64 | 4.63 | 0.00 | |
| LOC108228029 | 1.17 | 0.97 | 0.21 | |
| LOC108200853 | 3.32 | 2.87 | 0.44 | |
| LOC108223975 | −1.67 | −1.15 | −0.52 | |
| LOC108215374 | 1.04 | 0.89 | 0.15 | |
| LOC108220223 | 1.34 | 1.11 | 0.23 | |
| LOC108223887 | 1.16 | 0.91 | 0.25 | |
| LOC108198401 | −2.28 | −2.00 | −0.28 | |
| LOC108211917 | 4.53 | 4.12 | 0.41 | |
| LOC108217615 | 0.59 | 1.22 | −0.63 | |
| LOC108217616 | 1.54 | 1.21 | 0.33 | |
| PE | LOC108207846 | 2.16 | 1.86 | 0.30 |
| LOC108199564 | −2.82 | −1.91 | −0.90 | |
| LOC108214632 | 1.74 | 1.17 | 0.58 | |
| LOC108226789 | 2.36 | 1.89 | 0.47 | |
| LOC108215117 | 1.16 | 0.81 | 0.34 | |
| LOC108212157 | 2.75 | 2.13 | 0.62 | |
| LOC108220333 | 3.95 | 4.06 | −0.11 | |
| LOC108222117 | 3.31 | 2.90 | 0.40 | |
| LOC108206875 | 3.34 | 3.30 | 0.04 | |
| LOC108225443 | 2.79 | 2.96 | −0.17 | |
| LOC108199289 | 3.54 | 3.53 | 0.01 | |
| LOC108193704 | 1.08 | 0.94 | 0.14 | |
| LOC108211783 | 2.64 | 2.47 | 0.16 | |
| LOC108222122 | 7.56 | 6.42 | 1.14 | |
| LOC108199981 | 1.32 | 1.17 | 0.15 | |
| LOC108195530 | 1.91 | 1.40 | 0.51 | |
| LOC108217994 | 2.09 | 1.93 | 0.16 | |
| LOC108192921 | 0.95 | 0.79 | 0.16 | |
| PAE | LOC108222452 | 4.22 | 3.94 | 0.28 |
| LOC108224941 | 1.06 | 0.74 | 0.31 | |
| LOC108194222 | 1.47 | 1.05 | 0.42 | |
| LOC108223619 | 1.14 | 1.04 | 0.10 | |
| PAL | LOC108223317 | 0.53 | 0.18 | 0.36 |
| LOC108227736 | 1.02 | 0.54 | 0.48 | |
| LOC108224761 | −2.85 | −3.36 | 0.50 | |
| LOC108217754 | 0.43 | 0.20 | 0.23 | |
| 4CL | LOC108216092 | 1.47 | 1.40 | 0.07 |
| LOC108193188 | 1.83 | 1.91 | −0.08 | |
| LOC108208836 | 0.83 | 1.37 | −0.54 | |
| CesA | LOC108209129 | 1.11 | 1.10 | 0.01 |
| LOC108208945 | 1.32 | 1.33 | −0.01 | |
| LOC108223639 | 2.22 | 2.60 | −0.39 | |
| LOC108222767 | 1.31 | 0.67 | 0.64 | |
| LOC108195360 | −1.14 | −1.03 | −0.10 | |
| LOC108213001 | −3.28 | −2.20 | −1.08 | |
| LOC108209273 | 1.90 | 1.63 | 0.27 | |
| LOC108214864 | −1.48 | −1.12 | −0.36 | |
| LOC108222895 | 3.17 | 1.43 | 1.74 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Yin, Y.; Wang, G.; Han, X.; Lan, Y.; Huang, D.; Zhao, S. Hydro-Electro Hybrid Priming Synchronizes Cell Wall Remodeling to Accelerate Carrot (Daucus carota L.) Seed Germination. Agronomy 2025, 15, 1147. https://doi.org/10.3390/agronomy15051147
Sun Y, Yin Y, Wang G, Han X, Lan Y, Huang D, Zhao S. Hydro-Electro Hybrid Priming Synchronizes Cell Wall Remodeling to Accelerate Carrot (Daucus carota L.) Seed Germination. Agronomy. 2025; 15(5):1147. https://doi.org/10.3390/agronomy15051147
Chicago/Turabian StyleSun, Yufan, Yilu Yin, Guobin Wang, Xin Han, Yubin Lan, Danfeng Huang, and Shuo Zhao. 2025. "Hydro-Electro Hybrid Priming Synchronizes Cell Wall Remodeling to Accelerate Carrot (Daucus carota L.) Seed Germination" Agronomy 15, no. 5: 1147. https://doi.org/10.3390/agronomy15051147
APA StyleSun, Y., Yin, Y., Wang, G., Han, X., Lan, Y., Huang, D., & Zhao, S. (2025). Hydro-Electro Hybrid Priming Synchronizes Cell Wall Remodeling to Accelerate Carrot (Daucus carota L.) Seed Germination. Agronomy, 15(5), 1147. https://doi.org/10.3390/agronomy15051147