Optimizing Nitrogen Application Enhances Sugar Beet (Beta vulgaris L.) Productivity by Modulating Carbon and Nitrogen Metabolism


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
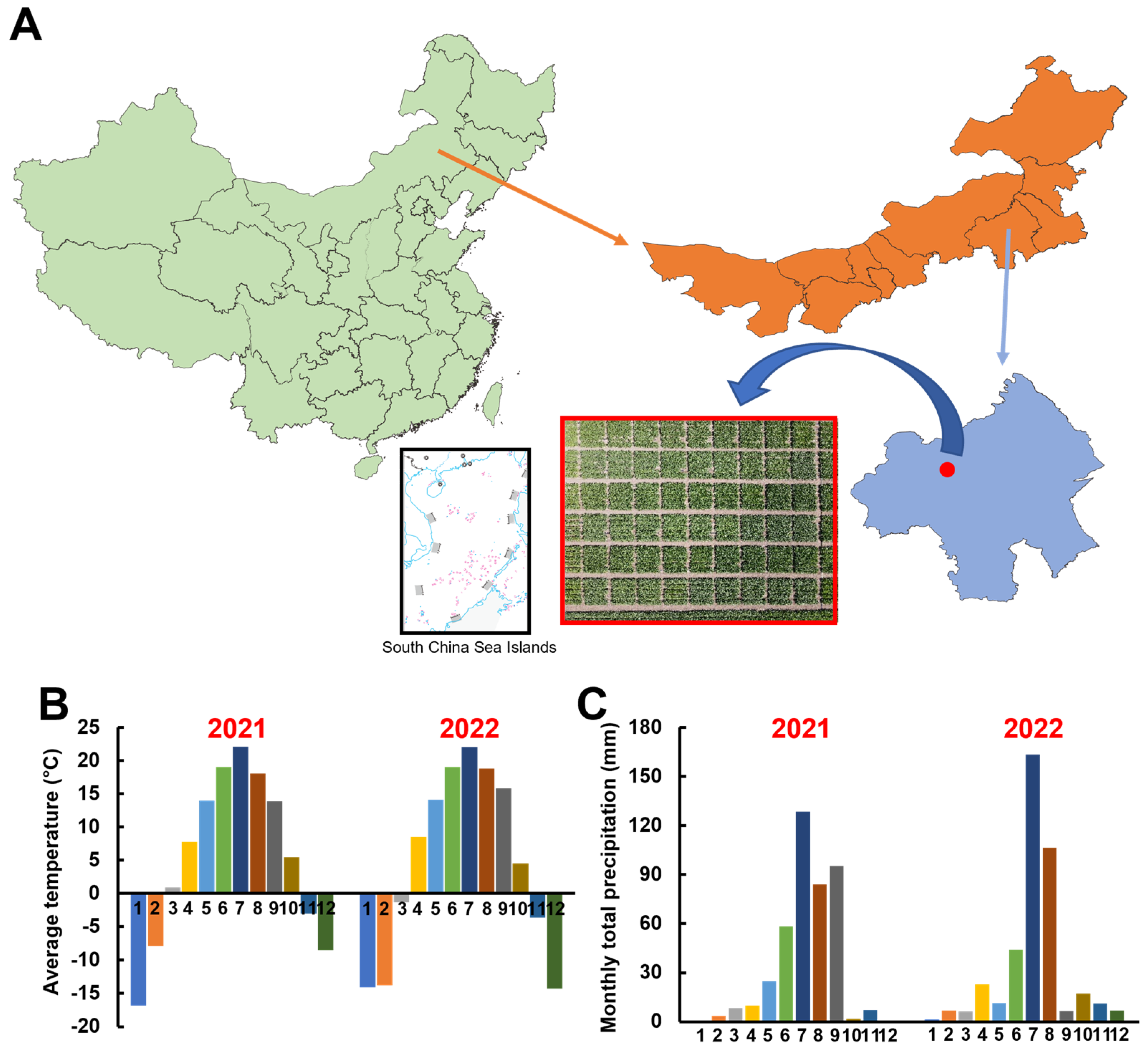
2.1. Experimental Site
2.2. Experimental Design
2.3. Sampling Period
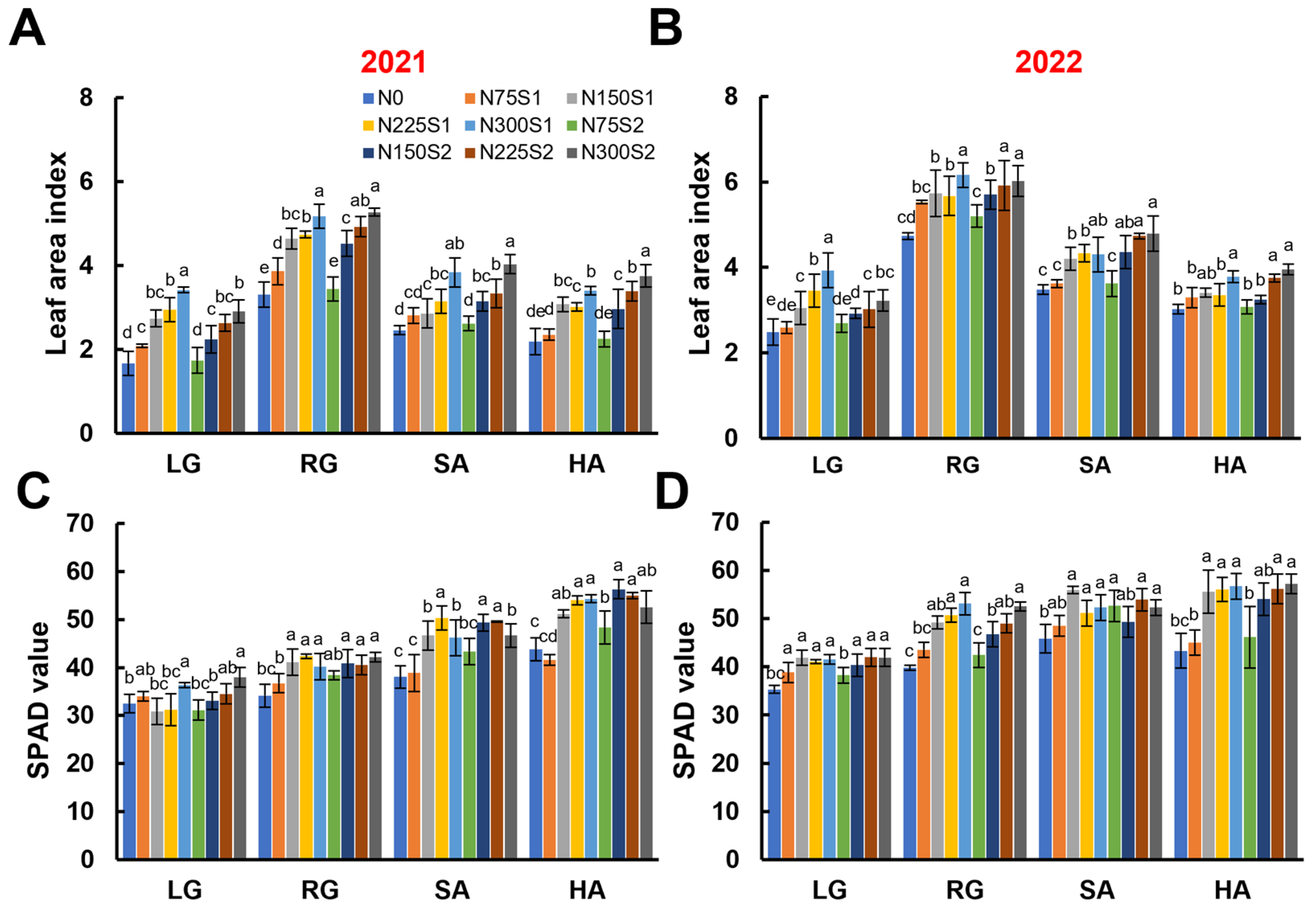
2.4. Leaf Area Index and SPAD Value
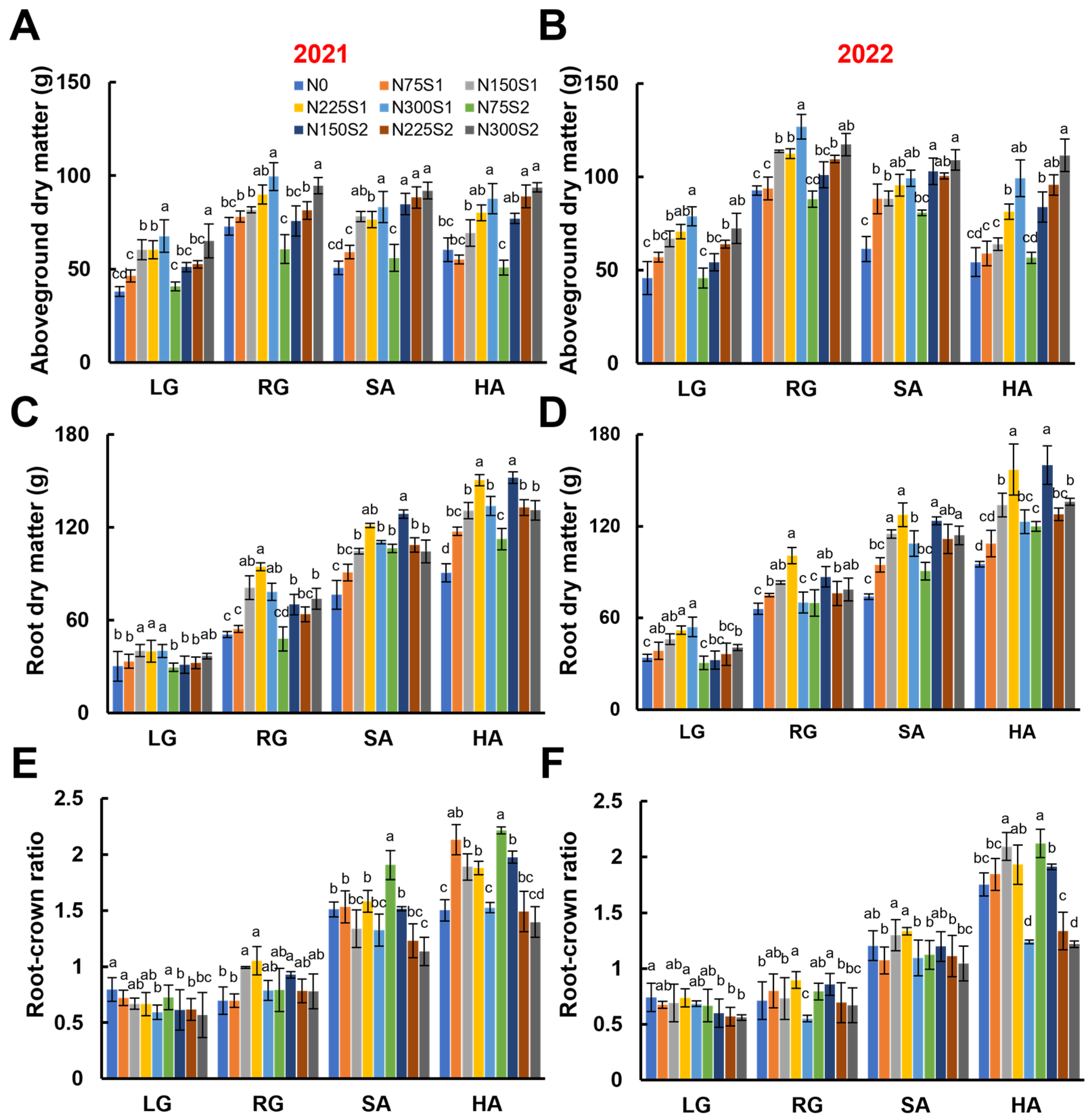
2.5. Dry Matter Accumulation
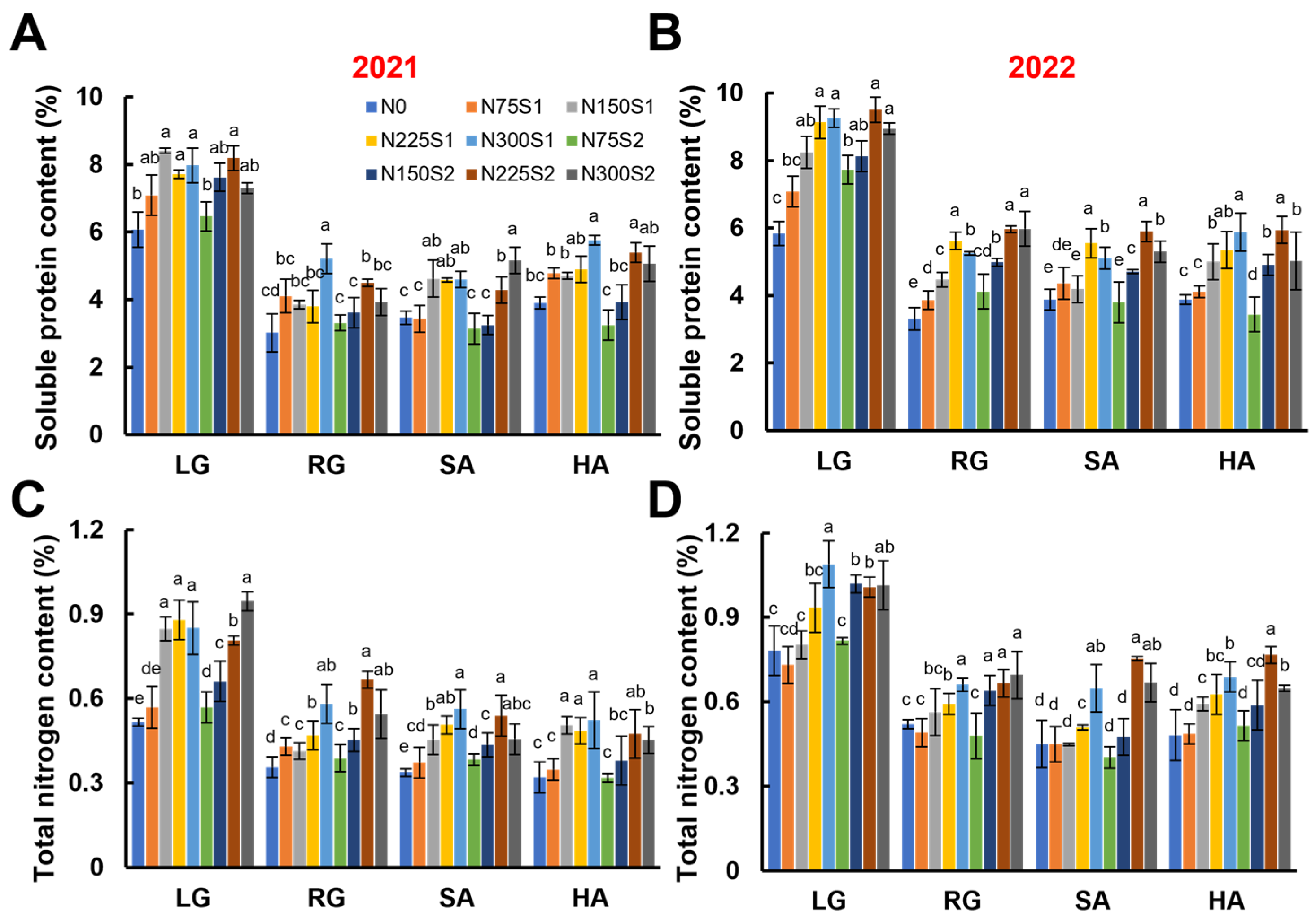
2.6. Carbon and Nitrogen Metabolism Analysis
2.7. Determination of Agronomic Nitrogen Use Efficiency, Sugar Content, Root Yield, and Sugar Yield
2.8. Statistical Analysis
3. Results
3.1. Leaf-Related Parameters During the Growth Stages of Sugar Beet
3.2. Dry Matter Accumulation for the Aboveground Parts and Roots During the Growth Stages of Sugar Beet
3.3. Nitrogen Metabolism During the Growth Stages of Sugar Beet
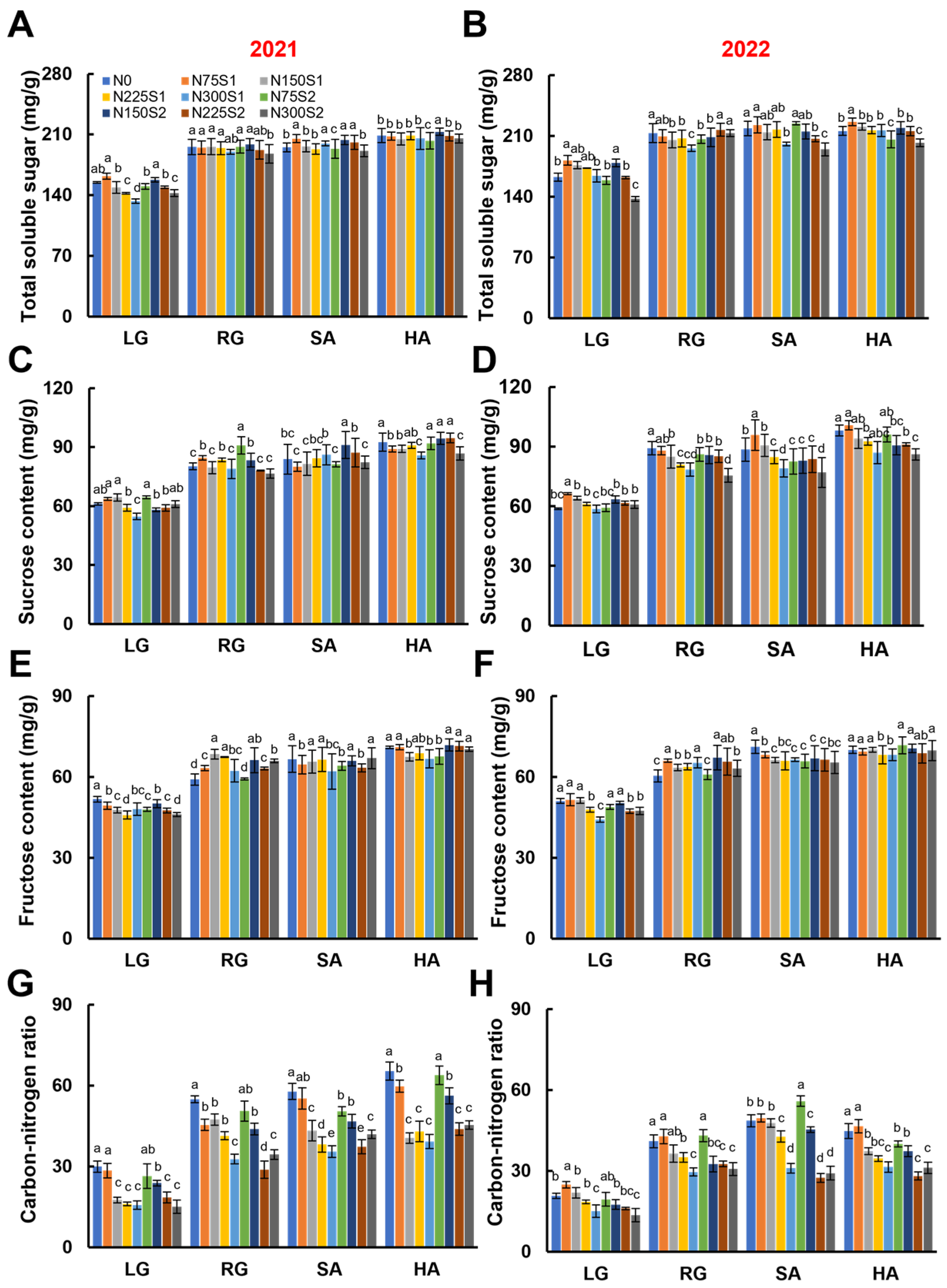
3.4. Carbon Metabolism During the Growth Stages of Sugar Beet
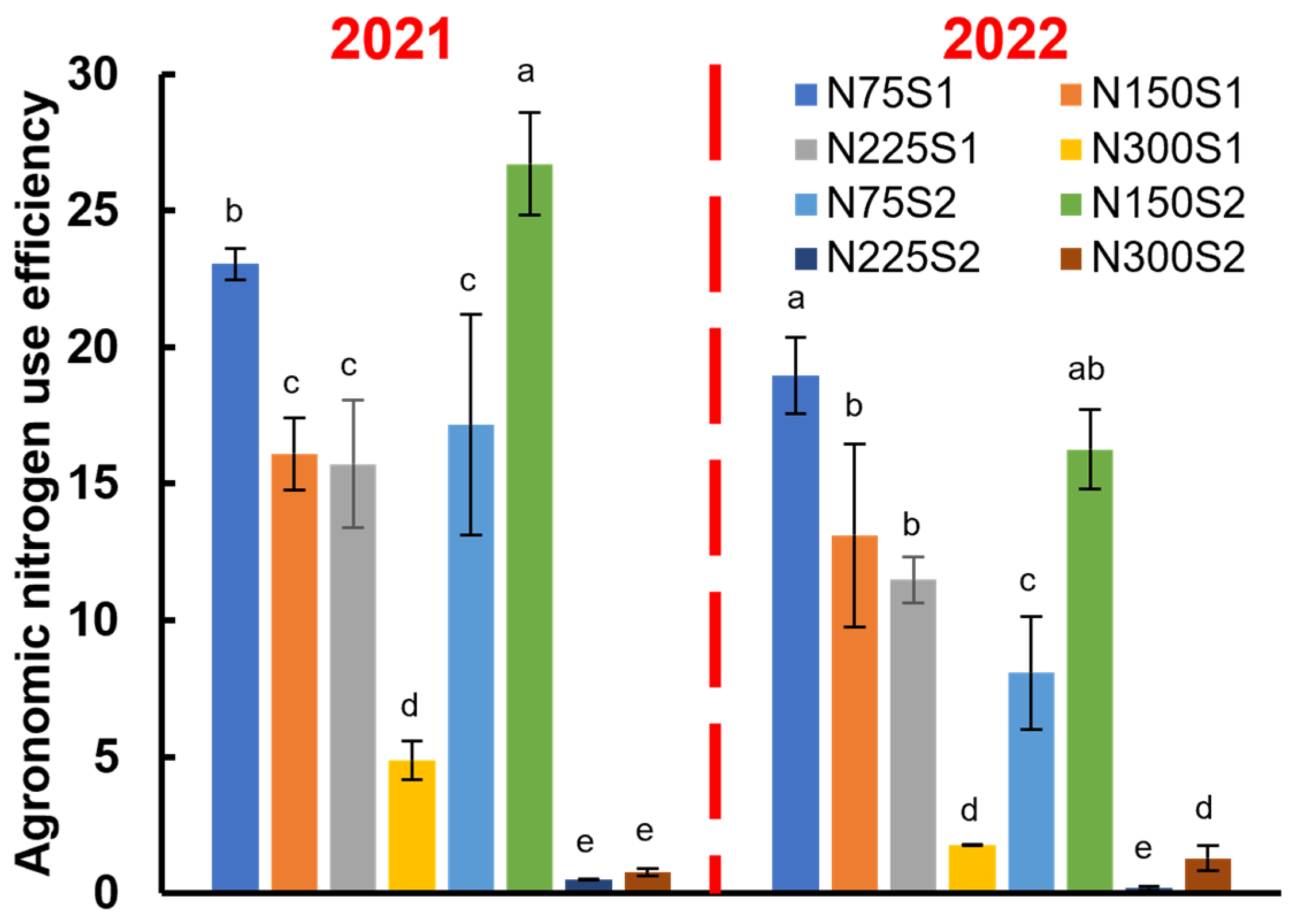
3.5. Agronomic Nitrogen Use Efficiency
3.6. Root Yield and Quality
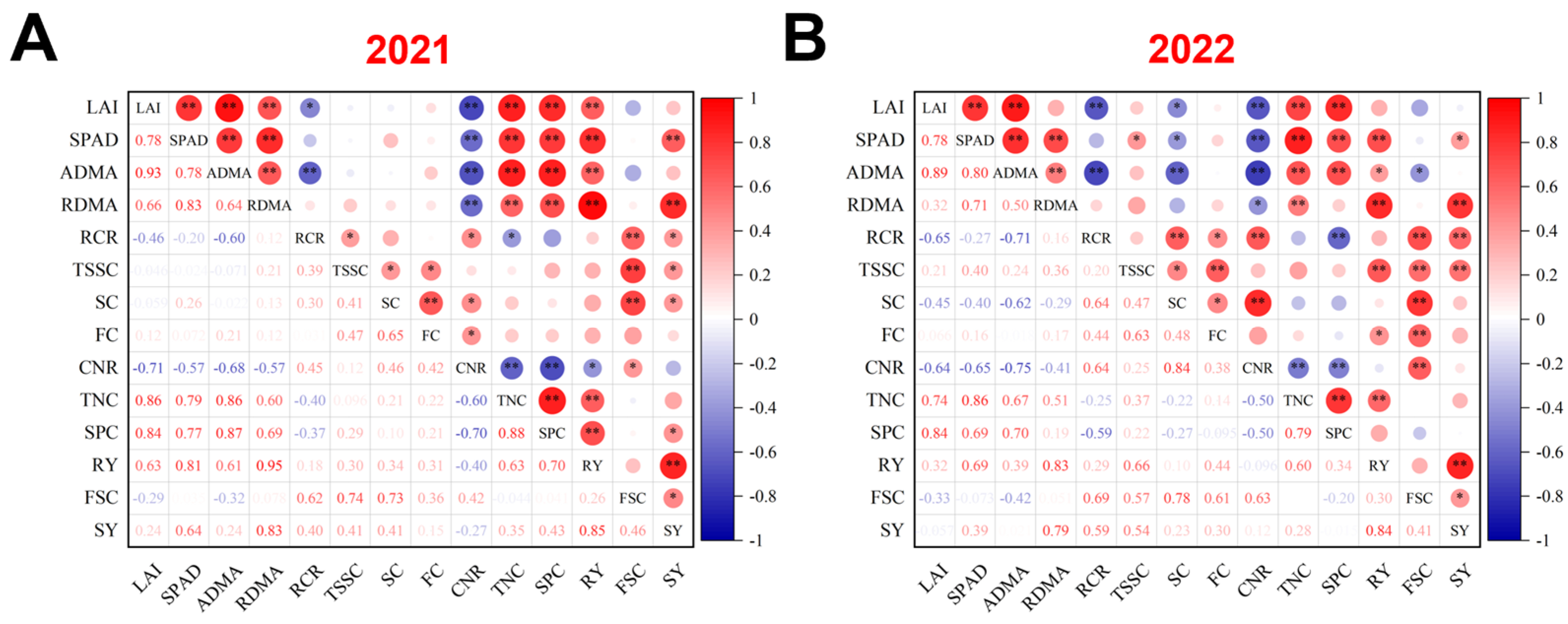
3.7. Correlation Analysis Between Physiological Parameters, Yield, and Quality of Sugar Beet
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LAI | Leaf area index |
| CCOs | Continuous cropping obstacles |
| GOT | Glutamic oxaloacetate transaminase |
| GPT | Glutamate pyruvate transaminase |
| GBSS | Granule-bound starch synthase |
| SSS | Soluble starch synthase |
| LG | Leaf growth stage |
| RG | Root growth stage |
| SA | Sugar accumulation stage |
| HA | Harvest stage |
| C/N | Carbon–nitrogen |
| ANUE | Agronomic nitrogen use efficiency |
| ADMA | Aboveground dry matter accumulation |
| RDMA | Root dry matter accumulation |
| RCR | Root–crown ratio |
| TSSC | Total soluble sugar content |
| SC | Sucrose content |
| FC | Fructose content |
| CNR | Carbon–nitrogen ratio |
| TNC | Total nitrogen content |
| SPC | Soluble protein content |
| RY | Root yield |
| FSC | Final sugar content |
| SY | Sugar yield |
References
- Ebrahimi, P.; Khamirikar, F.; Lante, A. Unlocking the Biorefinery Approaches to Valorize Sugar Beet Leaves (B. vulgaris L.) for Food Industry Applications: A Critical Review. Food Res. Int. 2024, 197, 115145. [Google Scholar] [CrossRef]
- Gruska, R.M.; Baryga, A.; Kunicka-Styczyńska, A.; Brzeziński, S.; Rosicka-Kaczmarek, J.; Miśkiewicz, K.; Sumińska, T. Fresh and Stored Sugar Beet Roots as a Source of Various Types of Mono- and Oligosaccharides. Molecules 2022, 27, 5125. [Google Scholar] [CrossRef]
- Tomaszewska, J.; Bieliński, D.; Binczarski, M.; Berlowska, J.; Dziugan, P.; Piotrowski, J.; Stanishevsky, A.; Witońska, I.A. Products of Sugar Beet Processing as Raw Materials for Chemicals and Biodegradable Polymers. RSC Adv. 2018, 8, 3161–3177. [Google Scholar] [CrossRef]
- Xiao, S.; Chai, H.; Shao, K.; Shen, M.; Wang, Q.; Wang, R.; Sui, Y.; Ma, Y. Image-Based Dynamic Quantification of Aboveground Structure of Sugar Beet in Field. Remote Sens. 2020, 12, 269. [Google Scholar] [CrossRef]
- Usmani, Z.; Sharma, M.; Diwan, D.; Tripathi, M.; Whale, E.; Jayakody, L.N.; Moreau, B.; Thakur, V.K.; Tuohy, M.; Gupta, V.K. Valorization of Sugar Beet Pulp to Value-Added Products: A Review. Bioresour. Technol. 2022, 346, 126580. [Google Scholar] [CrossRef]
- Yousefabadi, V.A.; Mehdikhani, P.; Nadali, F.; Sharifi, M.; Azizi, H.; Ahmadi, M.; Fasahat, P. Evaluation of Yield and Stability of Sugar Beet (Beta vulgaris L.) Genotypes Using GGE Biplot and AMMI Analysis. Sci. Rep. 2024, 14, 27384. [Google Scholar] [CrossRef]
- McGinnis, E.E.; Meyer, M.H.; Smith, A.G. Sweet and Sour: A Scientific and Legal Look at Herbicide-Tolerant Sugar Beet. Plant Cell 2010, 22, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Khusnitdinova, M.; Abdrakhmanova, A.; Pozharskiy, A.; Kapytina, A.; Kerimbek, N.; Nizamdinova, G.; Taskuzhina, A.; Adilbayeva, K.; Kolchenko, M.; Gritsenko, D. Problems and Prospects of Sugar Beet Cultivation in Kazakhstan. Agron. Res. 2023, 21, 1174–1185. [Google Scholar] [CrossRef]
- Tayyab, M.; Wakeel, A.; Mubarak, M.U.; Artyszak, A.; Ali, S.; Hakki, E.E.; Mahmood, K.; Song, B.; Ishfaq, M. Sugar Beet Cultivation in the Tropics and Subtropics: Challenges and Opportunities. Agronomy 2023, 13, 1213. [Google Scholar] [CrossRef]
- Wang, N.; Fu, F.; Wang, H.; Wang, P.; He, S.; Shao, H.; Ni, Z.; Zhang, X. Effects of Irrigation and Nitrogen on Chlorophyll Content, Dry Matter and Nitrogen Accumulation in Sugar Beet (Beta vulgaris L.). Sci. Rep. 2021, 11, 16651. [Google Scholar] [CrossRef]
- Wang, Q.; Shao, K.; Cai, Z.; Che, Y.; Chen, H.; Xiao, S.; Wang, R.; Liu, Y.; Li, B.; Ma, Y. Prediction of Sugar Beet Yield and Quality Parameters Using Stacked-LSTM Model with Pre-Harvest UAV Time Series Data and Meteorological Factors. Artif. Intell. Agric. 2025, 15, 252–265. [Google Scholar] [CrossRef]
- Chang, Y.; Li, G.; Jian, C.; Zhang, B.; Sun, Y.; Li, N.; Zhang, S. Influence of Water and Fertilizer Reduction on Respiratory Metabolism in Sugar Beet Taproot (Beta vulgaris L.). Plants 2024, 13, 2282. [Google Scholar] [CrossRef]
- Guo, X.-X.; Tian, L.; Li, Y.-H.; Huang, C.-Y.; Li, Z.; Zhang, P.; Jian, C.-Y.; Han, K.; Zhou, J.-C. Effects of Continuous Cropping and Application of Bio-Organic Fertilizer on Photosynthetic Performance, Dry Matter Accumulation and Distribution of Sugar Beet. Sci. Rep. 2025, 15, 1512. [Google Scholar] [CrossRef] [PubMed]
- Amirahmadi, E.; Ghorbani, M.; Krexner, T.; Hörtenhuber, S.J.; Bernas, J.; Neugschwandtner, R.W.; Konvalina, P.; Moudrý, J. Life Cycle Assessment of Biochar and Cattle Manure Application in Sugar Beet Cultivation—Insights into Root Yields, White Sugar Quality, Environmental Aspects in Field and Factory Phases. J. Clean. Prod. 2024, 476, 143772. [Google Scholar] [CrossRef]
- Huang, W.; Han, S.; Wang, L.; Li, W. Carbon and Nitrogen Metabolic Regulation in Freshwater Plant Ottelia Alismoides in Response to Carbon Limitation: A Metabolite Perspective. Front. Plant Sci. 2022, 13, 962622. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and Signaling Aspects Underpinning the Regulation of Plant Carbon Nitrogen Interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Zayed, O.; Hewedy, O.A.; Abdelmoteleb, A.; Ali, M.; Youssef, M.S.; Roumia, A.F.; Seymour, D.; Yuan, Z.C. Nitrogen Journey in Plants: From Uptake to Metabolism, Stress Response, and Microbe Interaction. Biomolecules 2023, 13, 1443. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Q.; Wang, Z.; Wang, L.; Li, X.; Fan, Z.; Zhang, Y.; Li, J.; Gao, X.; Shi, J.; et al. Effects of Nitrogen Fertilizer on Photosynthetic Characteristics, Biomass, and Yield of Wheat under Different Shading Conditions. Agronomy 2021, 11, 1989. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Li, Q.; Zeng, X.P.; Liu, Y.; Li, Y.R. Fate of Nitrogen in Agriculture and Environment: Agronomic, Eco-Physiological and Molecular Approaches to Improve Nitrogen Use Efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Baslam, M.; Mitsui, T.; Sueyoshi, K.; Ohyama, T. Recent Advances in Carbon and Nitrogen Metabolism in C3 Plants. Int. J. Mol. Sci. 2021, 22, 318. [Google Scholar] [CrossRef]
- Hu, W.; Coomer, T.D.; Loka, D.A.; Oosterhuis, D.M.; Zhou, Z. Potassium Deficiency Affects the Carbon-Nitrogen Balance in Cotton Leaves. Plant Physiol. Biochem. 2017, 115, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, X.; Liu, Y.; Sun, G.; Kong, D.; Guo, W.; Sun, H. Regulatory Effect of Graphene on Growth and Carbon/Nitrogen Metabolism of Maize (Zea mays L.). J. Sci. Food Agric. 2024, 104, 1572–1582. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant Carbon Metabolism and Climate Change: Elevated CO2 and Temperature Impacts on Photosynthesis, Photorespiration and Respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Yang, S.; Ma, A.; Lunzhu, C.; Wang, M.; Wang, G.; Guo, S. Grain Chalkiness Is Reduced by Coordinating the Biosynthesis of Protein and Starch in Fragrant Rice (Oryza sativa L.) Grain under Nitrogen Fertilization. Field Crop. Res. 2023, 302, 109098. [Google Scholar] [CrossRef]
- Sun, J.; Jin, L.; Li, R.; Meng, X.; Jin, N.; Wang, S.; Xu, Z.; Liu, Z.; Lyu, J.; Yu, J. Effects of Different Forms and Proportions of Nitrogen on the Growth, Photosynthetic Characteristics, and Carbon and Nitrogen Metabolism in Tomato. Plants 2023, 12, 4175. [Google Scholar] [CrossRef]
- Iqbal, A.; Jing, N.; Qiang, D.; Kayoumu, M.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. Genotypic Variation in Carbon and Nitrogen Metabolism in the Cotton Subtending Leaves and Seed Cotton Yield under Various Nitrogen Levels. J. Sci. Food Agric. 2023, 103, 2602–2617. [Google Scholar] [CrossRef]
- Feng, W.; Xue, W.; Zhao, Z.; Shi, Z.; Wang, W.; Bai, Y.; Wang, H.; Qiu, P.; Xue, J.; Chen, B. Nitrogen Fertilizer Application Rate Affects the Dynamic Metabolism of Nitrogen and Carbohydrates in Kernels of Waxy Maize. Front. Plant Sci. 2024, 15, 1416397. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Y.; Mu, B.; Wang, F.; Feng, N.; Zheng, D. Nitrogen Limitation Affects Carbon and Nitrogen Metabolism in Mung Bean (Vigna radiata L.). J. Plant Physiol. 2023, 290, 154105. [Google Scholar] [CrossRef]
- Li, J.; Feng, Y.; Wang, X.; Xu, G.; Luo, Z.; Peng, J.; Luo, Q.; Lu, W.; Han, Z. High Nitrogen Input Increases the Total Spikelets but Decreases the High-Density Grain Content in Hybrid Indica Rice. Field Crop. Res. 2022, 288, 108679. [Google Scholar] [CrossRef]
- Tanaka, M.; Keira, M.; Yoon, D.K.; Mae, T.; Ishida, H.; Makino, A.; Ishiyama, K. Photosynthetic Enhancement, Lifespan Extension, and Leaf Area Enlargement in Flag Leaves Increased the Yield of Transgenic Rice Plants Overproducing Rubisco Under Sufficient N Fertilization. Rice 2022, 15, 10. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, H.; Yan, H.; Jiang, X.; Ma, Y.; Qin, Y. Carbon and Nitrogen Metabolism under Nitrogen Variation Affects Flavonoid Accumulation in the Leaves of Coreopsis tinctoria. PeerJ 2021, 9, e12152. [Google Scholar] [CrossRef]
- Shi, Z.; Li, D.; Jing, Q.; Cai, J.; Jiang, D.; Cao, W.; Dai, T. Effects of Nitrogen Applications on Soil Nitrogen Balance and Nitrogen Utilization of Winter Wheat in a Rice-Wheat Rotation. Field Crop. Res. 2012, 127, 241–247. [Google Scholar] [CrossRef]
- Hou, W.; Xue, X.; Li, X.; Khan, M.R.; Yan, J.; Ren, T.; Cong, R.; Lu, J. Interactive Effects of Nitrogen and Potassium on: Grain Yield, Nitrogen Uptake and Nitrogen Use Efficiency of Rice in Low Potassium Fertility Soil in China. Field Crop. Res. 2019, 236, 14–23. [Google Scholar] [CrossRef]
- Li, J.; Liu, X.; Yao, Q.; Xu, L.; Li, W.; Tan, W.; Wang, Q.; Xing, W.; Liu, D. Tolerance and Adaptation Characteristics of Sugar Beet (Beta vulgaris L.) to Low Nitrogen Supply. Plant Signal. Behav. 2023, 18, e2159155. [Google Scholar] [CrossRef]
- Malnou, C.S.; Jaggard, K.W.; Sparkes, D.L. Nitrogen Fertilizer and the Efficiency of the Sugar Beet Crop in Late Summer. Eur. J. Agron. 2008, 28, 47–56. [Google Scholar] [CrossRef]
- Sinta, Z.; Garo, G. Influence of Plant Density and Nitrogen Fertilizer Rates on Yield and Yield Components of Beetroot (Beta vulgaris L.). Int. J. Agron. 2021, 2021, 670243. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, L.; Xu, P.; Liu, D.; Zhang, L.; Hao, Y.; Wang, K.; Fan, H. Nitrogen Use Efficiency of Drip Irrigated Sugar Beet as Affected by Sub-Optimal Levels of Nitrogen and Irrigation. Agric. Water Manag. 2024, 298, 108849. [Google Scholar] [CrossRef]
- Mekdad, A.A.A.; Shaaban, A. Integrative Applications of Nitrogen, Zinc, and Boron to Nutrients-Deficient Soil Improves Sugar Beet Productivity and Technological Sugar Contents under Semi-Arid Conditions. J. Plant Nutr. 2020, 43, 1935–1950. [Google Scholar] [CrossRef]
- Varga, I.; Jović, J.; Rastija, M.; Markulj Kulundžić, A.; Zebec, V.; Lončarić, Z.; Iljkić, D.; Antunović, M. Efficiency and Management of Nitrogen Fertilization in Sugar Beet as Spring Crop: A Review. Nitrogen 2022, 3, 170–185. [Google Scholar] [CrossRef]
- Tarkalson, D.D.; Bjorneberg, D.L.; Lentz, R.D. Effects of Manure History and Nitrogen Fertilizer Rate on Sugar Beet Production in the Northwest US. Crop. Forage Turfgrass Manag. 2018, 4, 170083. [Google Scholar] [CrossRef]
- Last, P.J.; Draycott, A.P.; Messem, A.B.; Webb, D.J. Effects of Nitrogen Fertilizer and Irriǵation on Suǵar Beet at Broom’s Barn 1973-8. J. Agric. Sci. 1983, 101, 185–205. [Google Scholar] [CrossRef]
- Reid, J.B.; Hunt, A.G.; Johnstone, P.R.; Searle, B.P. Beetroot (Beta vulgaris L.) Growth and Response to N Supply—A Case Study. N. Z. J. Crop. Hortic. Sci. 2020, 48, 191–212. [Google Scholar] [CrossRef]
- Idris, B.E.M.; Marajan, W.A.; Adam, A.H.M. Effect of Nitrogen Fertilizer and Plant Spacing on Vegetative Growth of Sugar Beet (Beta vulgaris L.). J. Agron. Res. 2021, 4, 6–13. [Google Scholar] [CrossRef]
- Su, J.; Zhou, H.; Wang, K.; Fan, H.; Hou, Z. Effects of Nitrogen Fertilizer Management on Dry Matter Accumulation and Yield of Drip-Irrigated Sugar Beet in Arid Areas. Agronomy 2024, 14, 1010. [Google Scholar] [CrossRef]
- Da, B.; Wu, Y.; Bao, W. Analysis of Spatial Distribution and Spillover Effects of Fertilizer Application Intensity in Inner Mongolia, China. Sustainability 2024, 16, 4697. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Zhang, J.; Li, X.; Jiang, Z.; Dong, S. Effects of DA-6 and MC on the Growth, Physiology, and Yield Characteristics of Soybean. BMC Plant Biol. 2025, 25, 304. [Google Scholar] [CrossRef]
- Su, B.Y.; Song, Y.X.; Song, C.; Cui, L.; Yong, T.W.; Yang, W.Y. Growth and Photosynthetic Responses of Soybean Seedlings to Maize Shading in Relay Intercropping System in Southwest China. Photosynthetica 2014, 52, 332–340. [Google Scholar] [CrossRef]
- Sun, J.; Li, W.; Li, C.; Chang, W.; Zhang, S.; Zeng, Y.; Zeng, C.; Peng, M. Effect of Different Rates of Nitrogen Fertilization on Crop Yield, Soil Properties and Leaf Physiological Attributes in Banana Under Subtropical Regions of China. Front. Plant Sci. 2020, 11, 613760. [Google Scholar] [CrossRef]
- Zhong, C.; Cao, X.; Hu, J.; Zhu, L.; Zhang, J.; Huang, J.; Jin, Q. Nitrogen Metabolism in Adaptation of Photosynthesis to Water Stress in Rice Grown under Different Nitrogen Levels. Front. Plant Sci. 2017, 8, 1079. [Google Scholar] [CrossRef]
- Lei, S.; Zeng, B.; Yuan, Z.; Su, X. Changes in Carbohydrate Content and Membrane Stability of Two Ecotypes of Calamagrostis arundinacea Growing at Different Elevations in the Drawdown Zone of the Three Gorges Reservoir. PLoS ONE 2014, 9, e91394. [Google Scholar] [CrossRef]
- Shi, H.; Wang, B.; Yang, P.; Li, Y.; Miao, F. Differences in Sugar Accumulation and Mobilization between Sequential and Non-Sequential Senescence Wheat Cultivars under Natural and Drought Conditions. PLoS ONE 2016, 11, e0166155. [Google Scholar] [CrossRef]
- Cai, Y.; Shao, L.; Li, X.; Liu, G.; Chen, S. Gibberellin Stimulates Regrowth after Defoliation of Sheepgrass (Leymus chinensis) by Regulating Expression of Fructan-Related Genes. J. Plant Res. 2016, 129, 935–944. [Google Scholar] [CrossRef]
- Wan, X.; Wu, W.; Shah, F. Nitrogen Fertilizer Management for Mitigating Ammonia Emission and Increasing Nitrogen Use Efficiencies by 15N Stable Isotopes in Winter Wheat. Sci. Total Environ. 2021, 790, 147587. [Google Scholar] [CrossRef]
- Yang, S.; Ge, Y.; Wang, J.; Liu, R.; Tang, D.; Li, A.; Zhu, Z. A Dataset for Estimating Alfalfa Leaf Area and Predicting Leaf Area Index. Front. Plant Sci. 2024, 15, 1290920. [Google Scholar] [CrossRef]
- Kubar, M.S.; Wang, C.; Noor, R.S.; Feng, M.; Yang, W.; Kubar, K.A.; Soomro, K.; Yang, C.; Sun, H.; Mohamed, H.; et al. Nitrogen Fertilizer Application Rates and Ratios Promote the Biochemical and Physiological Attributes of Winter Wheat. Front. Plant Sci. 2022, 13, 1011515. [Google Scholar] [CrossRef]
- Lan, T.; Du, L.; Wang, X.; Zhan, X.; Liu, Q.; Wei, G.; Lyu, C.; Liu, F.; Gao, J.; Feng, D.; et al. Synergistic Effects of Planting Density and Nitrogen Fertilization on Chlorophyll Degradation and Leaf Senescence after Silking in Maize. Crop. J. 2024, 12, 605–613. [Google Scholar] [CrossRef]
- Linders, K.M.; Santra, D.; Schnable, J.C.; Sigmon, B. Variation in Leaf Chlorophyll Concentration in Response to Nitrogen Application across Maize Hybrids in Contrasting Environments. Micropubl. Biol. 2024, 2024, 001115. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, S.M. Effects of SPAD Value Variations According to Nitrogen Application Levels on Rice Yield and Its Components. Front. Plant Sci. 2024, 15, 1437371. [Google Scholar] [CrossRef]
- Xu, X.; Yang, G.; Yang, X.; Li, Z.; Feng, H.; Xu, B.; Zhao, X. Monitoring Ratio of Carbon to Nitrogen (C/N) in Wheat and Barley Leaves by Using Spectral Slope Features with Branch-and-Bound Algorithm. Sci. Rep. 2018, 8, 10034. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Lončarić, Z.; Kristek, S.; Kulundžić, A.M.; Rebekić, A.; Antunović, M. Sugar Beet Root Yield and Quality with Leaf Seasonal Dynamics in Relation to Planting Densities and Nitrogen Fertilization. Agriculture 2021, 11, 407. [Google Scholar] [CrossRef]
- Duan, J.; Shao, Y.; He, L.; Li, X.; Hou, G.; Li, S.; Feng, W.; Zhu, Y.; Wang, Y.; Xie, Y. Optimizing Nitrogen Management to Achieve High Yield, High Nitrogen Efficiency and Low Nitrogen Emission in Winter Wheat. Sci. Total Environ. 2019, 697, 134088. [Google Scholar] [CrossRef] [PubMed]
- Noor, H.; Ding, P.; Ren, A.; Sun, M.; Gao, Z. Effects of Nitrogen Fertilizer on Photosynthetic Characteristics and Yield. Agronomy 2023, 13, 1550. [Google Scholar] [CrossRef]
- Guo, C.; Yuan, X.; Yan, F.; Xiang, K.; Wu, Y.; Zhang, Q.; Wang, Z.; He, L.; Fan, P.; Yang, Z.; et al. Nitrogen Application Rate Affects the Accumulation of Carbohydrates in Functional Leaves and Grains to Improve Grain Filling and Reduce the Occurrence of Chalkiness. Front. Plant Sci. 2022, 13, 921130. [Google Scholar] [CrossRef]
- Debele, M.; Taressa, B. Urea Split Application to Maize (Zea mays L.) Growth Stages of Medium Maturities, Influenced on Grain Yield and Parameter for Yield at Bako, East Wollega, Ethiopia. Int. J. Agron. 2023, 2023, 6673773. [Google Scholar] [CrossRef]
- Priyanka; Srivastava, P.K.; Rawat, R. Retrieval of Leaf Chlorophyll Content Using Drone Imagery and Fusion with Sentinel-2 Data. Smart Agric. Technol. 2023, 6, 100353. [Google Scholar] [CrossRef]
- Mekdad, A.A.A.; Rady, M.M. Response of Beta Vulgaris L. to Nitrogen and Micronutrients in Dry Environment. Plant Soil Environ. 2016, 62, 23–29. [Google Scholar] [CrossRef]
- Elsayed, S.; El-Hendawy, S.; Elsherbiny, O.; Okasha, A.M.; Elmetwalli, A.H.; Elwakeel, A.E.; Memon, M.S.; Ibrahim, M.E.M.; Ibrahim, H.H. Estimating Chlorophyll Content, Production, and Quality of Sugar Beet under Various Nitrogen Levels Using Machine Learning Models and Novel Spectral Indices. Agronomy 2023, 13, 2743. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, G.; Chen, Y.; Jiang, Y.; Dai, Q.; Huo, Z.; Shi, Y.; Zhao, L.; Liao, P.; Wang, W.; et al. Excessive Nitrogen Application Leads to Lower Rice Yield and Grain Quality by Inhibiting the Grain Filling of Inferior Grains. Agriculture 2022, 12, 962. [Google Scholar] [CrossRef]
- Liang, W.; Zhang, Z.; Wen, X.; Liao, Y.; Liu, Y. Effect of Non-Structural Carbohydrate Accumulation in the Stem Pre-Anthesis on Grain Filling of Wheat Inferior Grain. Field Crop. Res. 2017, 211, 66–76. [Google Scholar] [CrossRef]
- Zhang, Y.; Ye, C.; Su, Y.; Peng, W.; Lu, R.; Liu, Y.; Huang, H.; He, X.; Yang, M.; Zhu, S. Soil Acidification Caused by Excessive Application of Nitrogen Fertilizer Aggravates Soil-Borne Diseases: Evidence from Literature Review and Field Trials. Agric. Ecosyst. Environ. 2022, 340, 108176. [Google Scholar] [CrossRef]
- Zhu, F.; Gilliam, F.S.; Mulder, J.; Yoh, M.; Mo, J.; Lu, X. Effects of Excess Nitrogen (N) on Fine Root Growth in Tropical Forests of Contrasting N Status. Forests 2022, 13, 1328. [Google Scholar] [CrossRef]
- Huang, K.; Li, M.; Li, R.; Rasul, F.; Shahzad, S.; Wu, C.; Shao, J.; Huang, G.; Li, R.; Almari, S.; et al. Soil Acidification and Salinity: The Importance of Biochar Application to Agricultural Soils. Front. Plant Sci. 2023, 14, 1206820. [Google Scholar] [CrossRef]
- Kabir, T.; De Laporte, A.; Nasielski, J.; Weersink, A. Adjusting Nitrogen Rates with Split Applications: Modelled Effects on N Losses and Profits Across Weather Scenarios. Eur. J. Agron. 2021, 129, 126328. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, M.; Liu, J.; Wang, W.; Liu, S. Fertilizer and Soil Nitrogen Utilization of Pear Trees as Affected by the Timing of Split Fertilizer Application in Rain-Fed Orchard. Sci. Hortic. 2019, 252, 363–369. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Shi, Y.; Yu, Z. Optimized Split Nitrogen Fertilizer Increase Photosynthesis, Grain Yield, Nitrogen Use Efficiency and Water Use Efficiency under Water-Saving Irrigation. Sci. Rep. 2020, 10, 20310. [Google Scholar] [CrossRef]
- Du, X.; Xi, M.; Kong, L. Split Application of Reduced Nitrogen Rate Improves Nitrogen Uptake and Use Efficiency in Sweetpotato. Sci. Rep. 2019, 9, 14058. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Abubakar, S.A.; Si, Z.; Kama, R.; Gao, Y.; Duan, A. Suitable Split Nitrogen Application Increases Grain Yield and Photosynthetic Capacity in Drip-Irrigated Winter Wheat (Triticum aestivum L.) under Different Water Regimes in the North China Plain. Front. Plant Sci. 2023, 13, 1105006. [Google Scholar] [CrossRef]
- Abo, A.A.; Mamo, M.A.; Handiso, Y.E. Influence of Nitrogen Fertilizer Rate and Its Time of Application on Productivity of Maize (Zea mays L.) in Hadero Tunto District, Kambata Tambaro Zone Southern Ethiopia. Int. J. Agric. Nat. Sci. 2022, 15, 149–162. Available online: https://ijans.org/index.php/ijans/article/view/577 (accessed on 15 February 2025).
- Panison, F.; Sangoi, L.; Durli, M.M.; Leolato, L.S.; Coelho, A.E.; Kuneski, H.F.; de Liz, V.O. Timing and Splitting of Nitrogen Side-Dress Fertilization of Early Corn Hybrids for High Grain Yield. Rev. Bras. Cienc. Solo 2019, 43, e0170338. [Google Scholar] [CrossRef]
- Bao, A.; Zhao, Z.; Ding, G.; Shi, L.; Xu, F.; Cai, H. Accumulated Expression Level of Cytosolic Glutamine Synthetase 1gene (OsGS1;1 or OsGS1;2) Alter Plant Development and the Carbon-Nitrogen Metabolic Status in Rice. PLoS ONE 2014, 9, e95581. [Google Scholar] [CrossRef]
- Ma, Z.; Cao, J.; Chen, X.; Yu, J. Differences in Carbon and Nitrogen Metabolism of Soft Japonica Rice in Southern China during Grain Filling Stage under Different Light and Nitrogen Fertilizer Conditions and Their Relationship with Rice Eating Quality. Front. Plant Sci. 2025, 16, 1534625. [Google Scholar] [CrossRef]
- Li, W.; Shan, Y.; Xiao, X.; Zheng, J.; Luo, Q.; Ouyang, S.; Zhang, G. Effect of Nitrogen and Sulfur Fertilization on Accumulation Characteristics and Physicochemical Properties of A- and B-Wheat Starch. J. Agric. Food Chem. 2013, 61, 2418–2425. [Google Scholar] [CrossRef] [PubMed]
- Olmedo Pico, L.B.; Vyn, T.J. Dry Matter Gains in Maize Kernels Are Dependent on Their Nitrogen Accumulation Rates and Duration during Grain Filling. Plants 2021, 10, 1222. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, D.; Maharjan, B. Optimizing Nitrogen Management to Enhance Irrigated Sugar Beet Yield and Quality. Agron. J. 2024, 16, 2564–2572. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, X.; Dong, S.; Guo, M.; Wei, L.; Shi, S. Optimizing Nitrogen Application Enhances Sugar Beet (Beta vulgaris L.) Productivity by Modulating Carbon and Nitrogen Metabolism. Agronomy 2025, 15, 1142. https://doi.org/10.3390/agronomy15051142
Xing X, Dong S, Guo M, Wei L, Shi S. Optimizing Nitrogen Application Enhances Sugar Beet (Beta vulgaris L.) Productivity by Modulating Carbon and Nitrogen Metabolism. Agronomy. 2025; 15(5):1142. https://doi.org/10.3390/agronomy15051142
Chicago/Turabian StyleXing, Xuming, Shifeng Dong, Ming Guo, Lei Wei, and Shude Shi. 2025. "Optimizing Nitrogen Application Enhances Sugar Beet (Beta vulgaris L.) Productivity by Modulating Carbon and Nitrogen Metabolism" Agronomy 15, no. 5: 1142. https://doi.org/10.3390/agronomy15051142
APA StyleXing, X., Dong, S., Guo, M., Wei, L., & Shi, S. (2025). Optimizing Nitrogen Application Enhances Sugar Beet (Beta vulgaris L.) Productivity by Modulating Carbon and Nitrogen Metabolism. Agronomy, 15(5), 1142. https://doi.org/10.3390/agronomy15051142