
Patterns of Change in Plant Leaf Functional Traits Along an Altitudinal Gradient in a Karst Climax Community

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
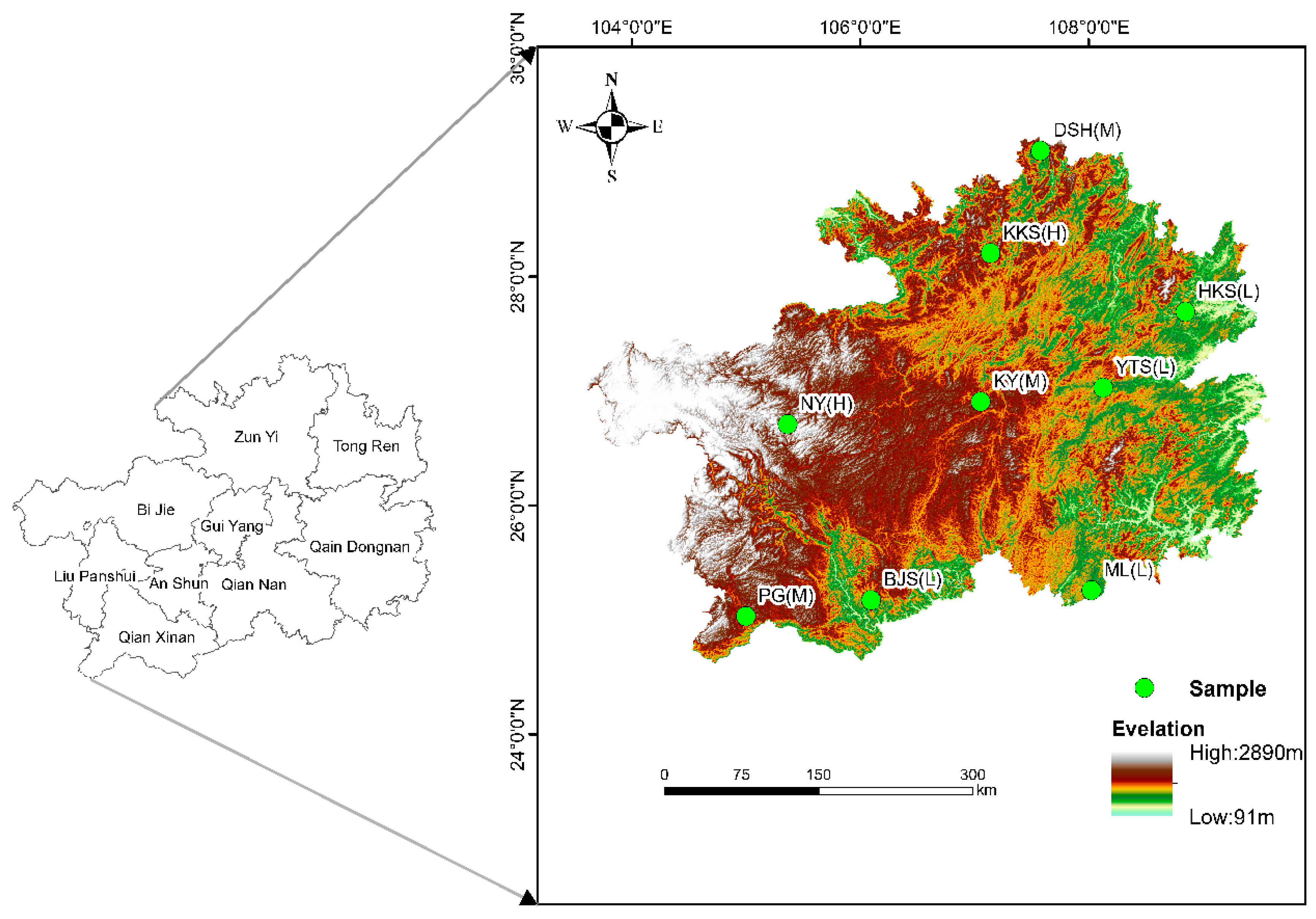
2.1. Study Area
2.2. Sample Setting and Sample Collection
2.3. Measurement Methods
3. Results
3.1. Changes in Plant Functional Traits Along the Altitudinal Gradient in the Karst Climax Community
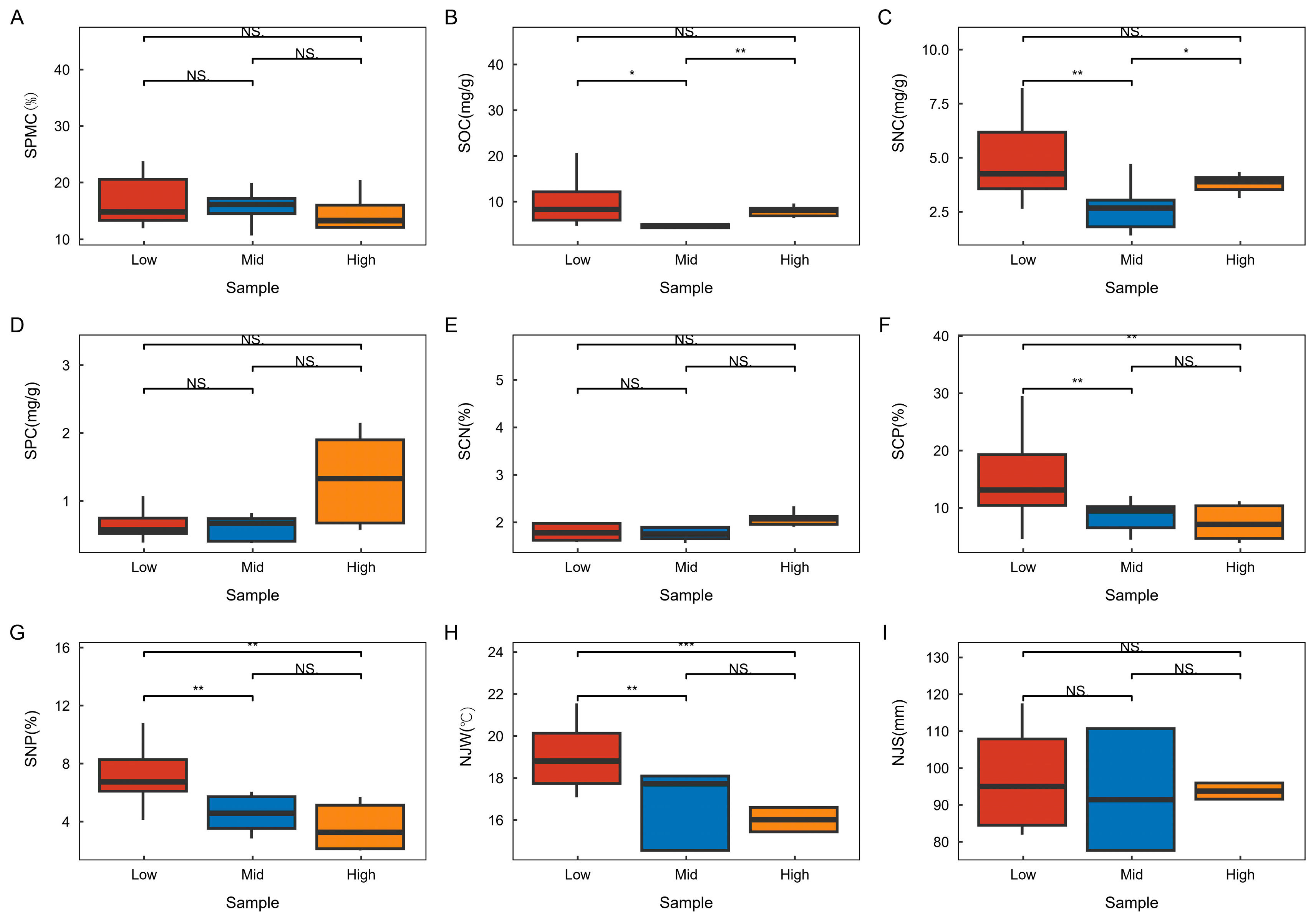
3.2. Changes in Environmental Factors Across Different Altitudinal Gradients in the Karst Summit Community
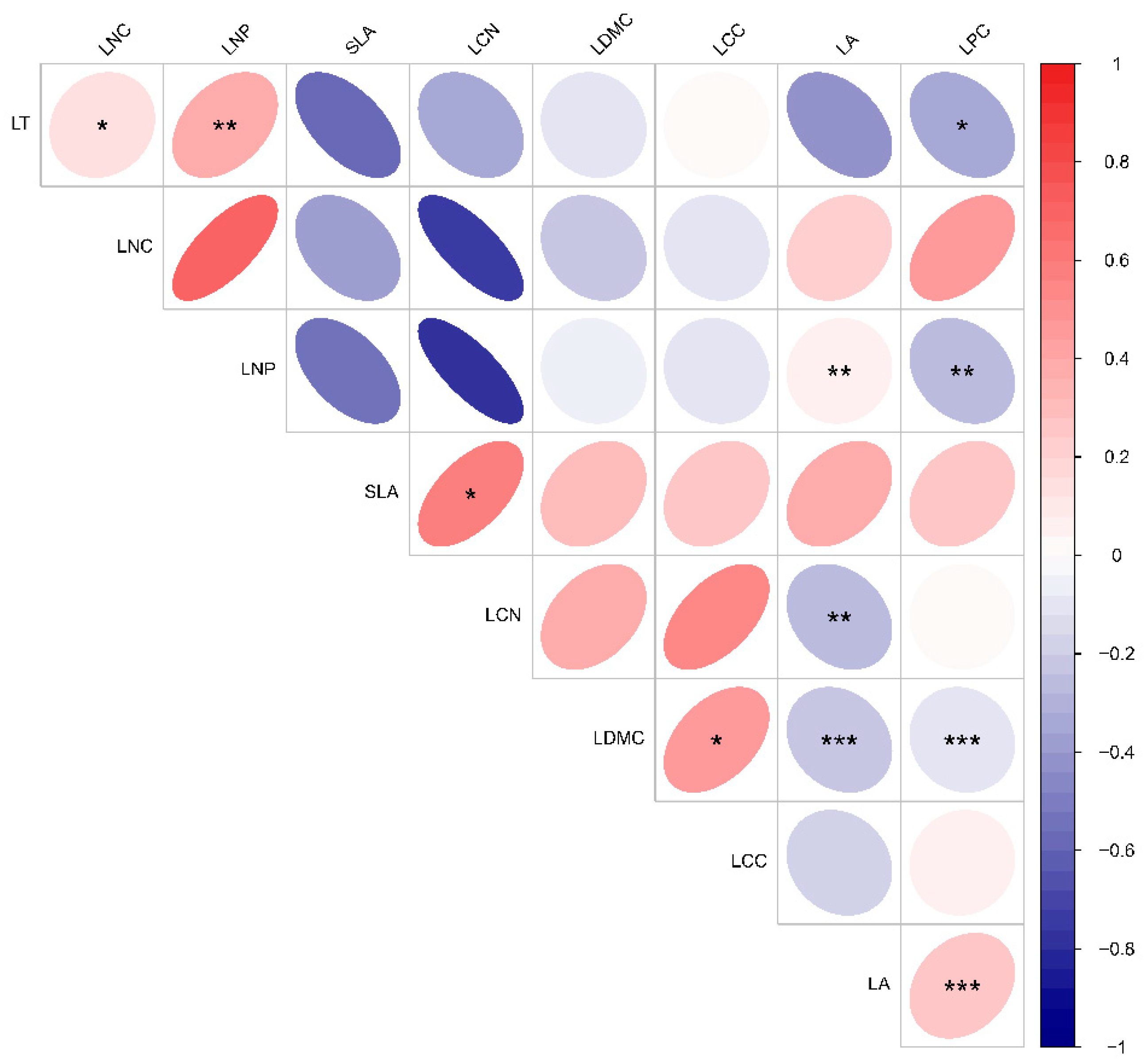
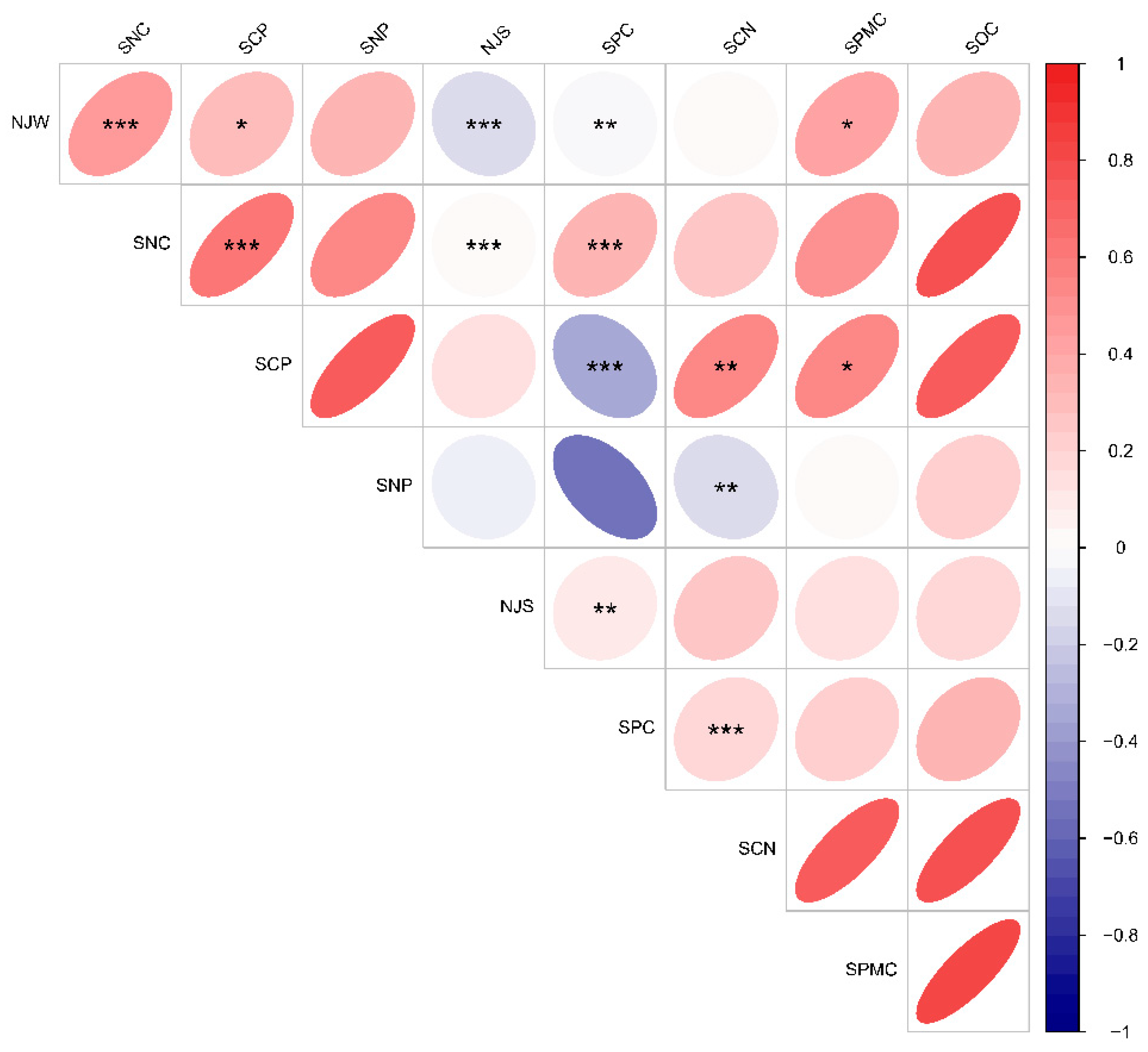
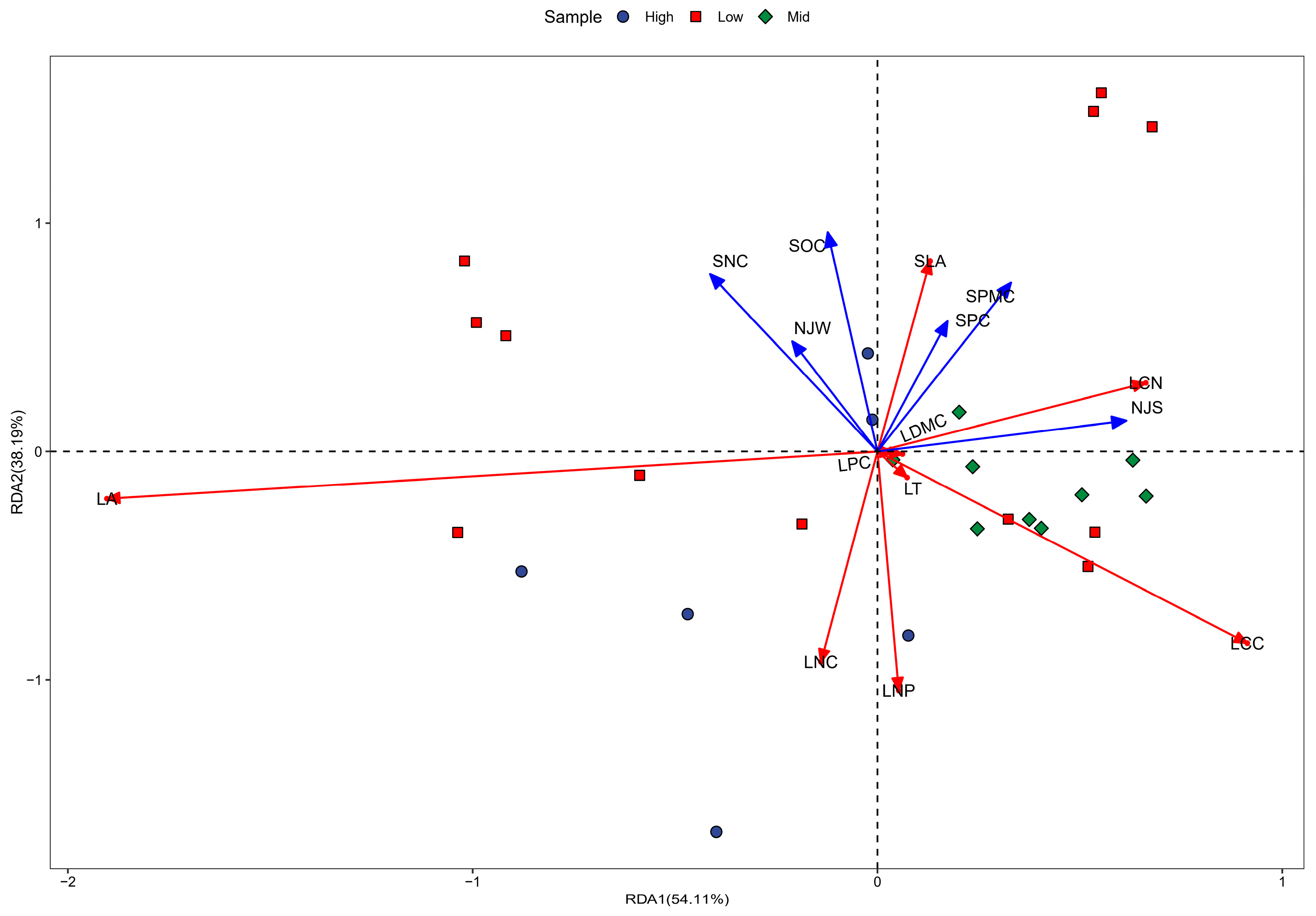
3.3. Variability of Leaf Functional Traits and Environmental Factors Across Altitudinal Gradients in Karst Climax Communities
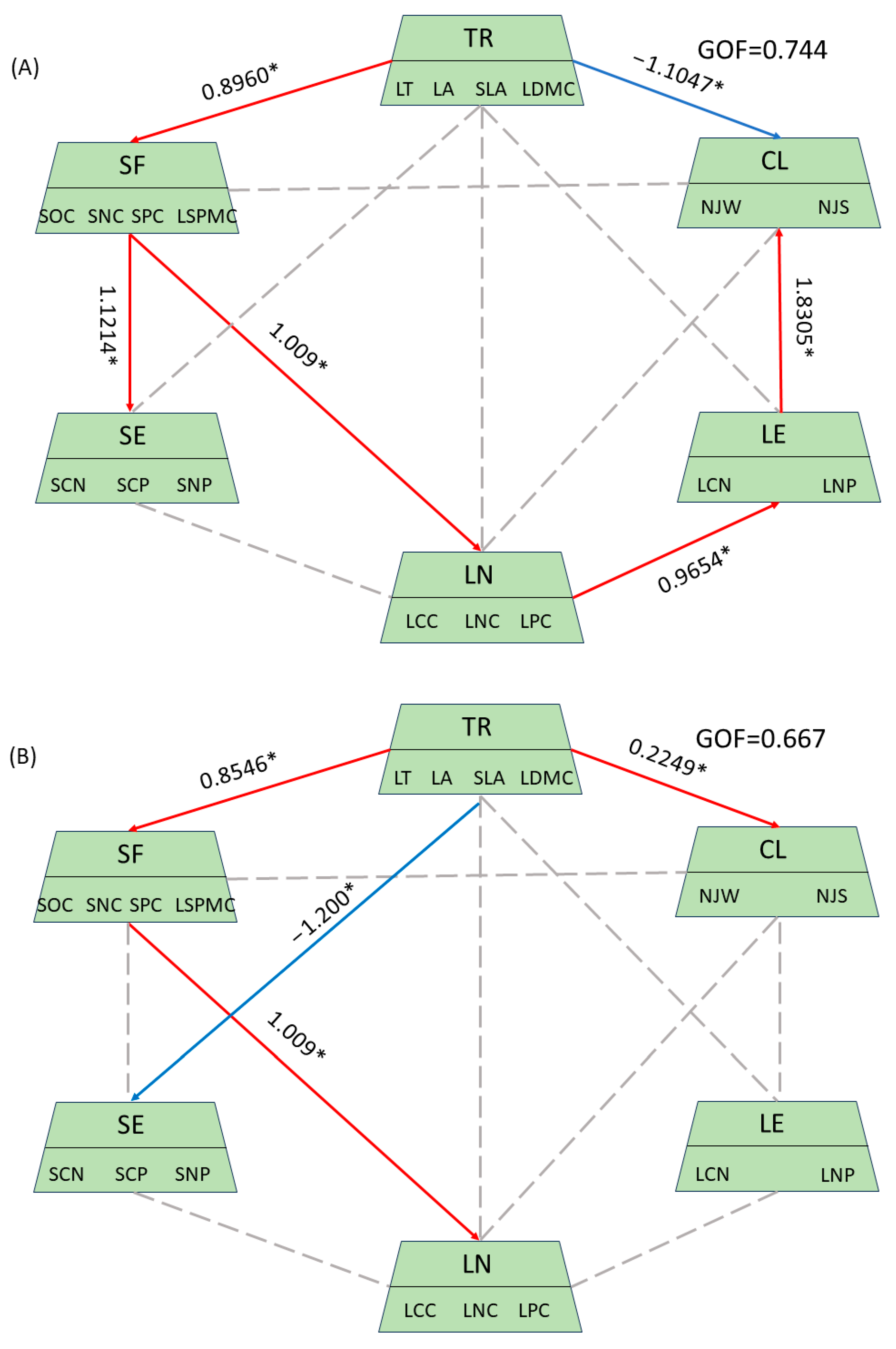
3.4. Response of Plant Functional Traits to Environmental Changes Along the Altitude Gradient
4. Discussion
4.1. Changes in Plant Functional Traits Along the Altitude Gradient
4.2. Variation in Plant Functional Traits and Environmental Factors in the Karst Climax Community
4.3. Response of Plant Functional Traits to Elevation Gradient
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Wang, Y.; He, N.; Ye, Z.; Chen, C.; Zang, R.; Feng, Y.; Lu, Q.; Li, J. Plant functional traits regulate soil bacterial diversity across temperate deserts. Sci. Total Environ. 2020, 715, 136976. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Li, Y.; Liu, C.; Xu, L.; Li, M.; Zhang, J.; He, J.; Tang, Z.; Han, X.; Ye, Q.; et al. Plant Trait Networks: Improved Resolution of the Dimensionality of Adaptation. Trends Ecol. Evol. 2020, 35, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Cheng, R.; Shi, Z.; Liu, S.; Wang, W.; Liu, X.; He, F. Response of leaf functional traits and the relationships among them to altitude of Salix dissa in Balang Mountai. Acta Ecol. Sin. 2013, 33, 2712–2718. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Yang, S.; Hou, J. Leaf functional traits of main tree species at different environmental gradients in Dongling Mountain, Beijing. For. Stud. China 2024, 36, 72–77. [Google Scholar]
- Dong, S.; Liu, E. Comparison of leaf functional traits of dominant woody plants on shady slope and sunny slope in the loessial hilly region. Resea. Soil Water Conser. 2015, 22, 326–331. [Google Scholar]
- Huang, Q.; Xie, L.; Cao, H.; Yang, F.; Ni, H. Variation characteristics of leaf functional traits of Populus davidiana in Wudalianchi Volcano, northeastern China. Forestry Stud. Chin. 2021, 43, 81–89. [Google Scholar]
- Qie, Y.; Jiang, L.; Lv, G.; Yang, X.; Wang, H.; Teng, D. Response of plant leaf functional traits to soil aridity and salinity in temperate desert ecosystem. J. Ecol. Environ. Sci. 2018, 27, 2000–2010. [Google Scholar]
- Brunet, J.; Holmquist, K.G. The influence of distinct pollinators on female and male reproductive success in the Rocky Mountain columbine. Mol. Ecol. 2009, 18, 3745–3758. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, J.; Ai, X.; Yao, L.; Huang, X.; Wu, M.; Zhu, Q.; Hong, J. Effects of topography on leaf functional traits across plant life forms in Xingdou Mountain, Hubei, China. Chin. J. Plant Ecol. 2019, 43, 447–457. [Google Scholar] [CrossRef]
- He, G.; Tian, Q.; Li, Z.; Song, L.; Zhang, S.; Cao, X.; Yang, W. Change in leaf functional traits of woody plants along altitudinal gradients at species and community levels on the motianling northern slope. Acta Bot. Boreali-Occident. Sin. 2018, 38, 553–563. [Google Scholar]
- Li, H.; Yu, Y. Leaf functional traits of Zanthoxylum planispinum var. dintanensis at different altitudes in dry-hot valley rocky desertification region. Guihaia 2020, 40, 782–791. [Google Scholar]
- Xu, L.; He, P.; Zhang, T.; Liu, H.; Ye, Q. Comparative studies on leaf hydraulic traits of six palm (Arecaceae) species originally distributed in different habitats. J. Trop. Subtrop. Bot. 2020, 28, 472–478. [Google Scholar]
- Wei, Y.; Wang, Z.; Liang, W.; Ma, F.; Han, L. Response and adaptation of twig-leaf functional traits of Populus euphratica to groundwater gradients. Acta Bot. Boreal. Occident. Sin. 2020, 40, 1043–1051. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westo, Y.M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Wang, R.; Yu, G.; He, N.; Wang, Q.; Zhao, N.; Zhao, N.; Xu, Z. Altitudinal variation in the covariation of stomatal traits with leaf functional traits in Changbai Mountain. Acta Ecol. Sin. 2016, 36, 2175–2184. [Google Scholar]
- Yang, R.; Zhang, B.; Wang, L.; Yang, J.; Su, W.; Zhang, G.; Yang, B.; Zhou, R. The response of plant functional traits’ group to gradients of altitude in dry-hot valley of Yuan-Mou. J. Ecol. Environ. Sci. 2015, 24, 49–56. [Google Scholar]
- Wang, C.; Lu, J.; Zhou, C.; Yao, H.; Jiayang, L.; La, B. Altitude distribution of leaf functional traits of Quercus aquifolioides in southeastern Tibet. J. Agrofor. Environ. 2021, 41, 366–372. [Google Scholar]
- Khatri, K.; Negi, B.; Bargali, K.; Bargali, S.S. Phenotypic variation in morphology and associated functional traits in Ageratina adenophora along an altitudinal gradient in Kumaun Himalaya, India. Biologia 2023, 78, 1333–1347. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Descombes, P.; Glauser, G.; Pellissier, L.; Rasmann, S. Plant physical and chemical defence variation along elevation gradients: A functional trait-based approach. Oecologia 2018, 187, 561–571. [Google Scholar] [CrossRef]
- Zhang, L.; Tan, X.; Dong, Z.; Zheng, J.; Yuan, Z.; Li, C. The relationship between plant functional traits and soil physicochemical properties in the riparian zones of downtown Chongqing. Acta Ecol. Sin. 2023, 43, 1892–1901. [Google Scholar]
- Liu, X.; Li, L.; Che, Y.; Jiao, J. Functional traits of plant leaves at different succession stages in alpine meadow. Bull. Bota. Res. 2019, 39, 760–769. [Google Scholar]
- Zhou, X.; Xin, J.; Huang, X.; Li, H.; Li, F.; Song, W. Linking Leaf Functional Traits with Soil and Climate Factors in Forest Ecosystems in China. Plants 2022, 11, 3545. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Wang, X.; Shen, Z.; Tang, Z.; He, J.; Yu, D.; Jiang, Y.; Wang, Z.; Zheng, C.; Zhu, J.; et al. Methods and protocols for plant community inventory. Biodiv. Sci. 2009, 17, 533–548. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; Agricultural Press: Beijing, China, 2005; pp. 45–52. [Google Scholar]
- NY/T 2017-2011; Determination of Nitrogen, Phosphorus and Potassium in Plants. Ministry of Agriculture: Beijing, China, 2011.
- LY/T1228-2015; Determination of Nitrogen in Forest Soil. Ministry of Agriculture: Beijing, China, 2015.
- LY/T 1232-2015; Phosphorus Determination Methods of Forest Soils. Ministry of Agriculture: Beijing, China, 2015.
- Pérezharguindeguy, N.; Díaz, S.; Garnier, E. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field-methodology matters. Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 12 February 2025).
- Gaston, S. PLS Path Modeling with R; Trowchez: Berkeley, CA, USA, 2013. [Google Scholar]
- Holscher, D.; Schmitt, S.; Kupfer, K. Growth and leaf traits of four broad-leaved tree species along a hillside gradient. Forstwisseschaftliches Cent. 2002, 121, 229–239. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, J.; Wang, G.; Zhou, J.; Liu, Y.; Ha, W. A study of functional traits of natural secondary forests and their influencing factors in different succession stages in karst areas: A case study of Dahei Mountain, Yunnan Province. Acta Ecol. Sin. 2021, 42, 397–406. [Google Scholar]
- Wang, Y.; He, M.; Jiang, G.; Yin, P.; Ying, W.; Yang, Q. Characteristics of leaf functional traits of Quercus spinosa and their responses to environmental factors at different altitude gradients in Wumeng township. Acta Ecol. Sin. 2024, 44, 7238–7248. [Google Scholar]
- Reich, P.B.; Wang, Z.H. Photosynthesis-nitrogen relation in Aamazonian tree species: Ⅱ. Variation vis-a-vis specific leaf influenes mass and area-based expessions. Oecologia 1994, 97, 73–81. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- Liu, N.; Yu, L.F.; Zhao, Q.; Wu, Y.N.; Yan, L.B. C:N:P stoichiometry of leaf-litter-soil continuum in secondary forests of the rocky desertification regions of the karst plateau. Chin. J. Appl. Environ. Biol. 2020, 26, 681–688. [Google Scholar]
- Zeng, Z.; Wang, K.; Liu, X.; Zeng, F.; Song, T.; Peng, W.; Zhang, H.; Du, H. Stoichiometric characteristics of live fresh leaves and leaf litter from typical plant communities in a karst region of northwestern Guangxi, China. Acta Ecol. Sin. 2016, 36, 1907–1914. [Google Scholar]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef]
- Dong, N.; Prentice, I.C.; Wright, I.J.; Evans, B.J.; Togashi, H.F.; Caddy-Retalic, S.; McInerney, F.A.; Sparrow, B.; Leitch, E.; Lowe, A.J. Components of leaf-trait variation along environmental gradients. New Phytol. 2020, 228, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Siefert, A.; Violle, C.; Chalmandrier, L.; Albert, C.H.; Taudiere, A.; Fajardo, A.; Aarssen, L.W.; Baraloto, C.; Carlucci, M.B.; Cianciaruso, M.V.; et al. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol. Lett. 2015, 18, 1406–1419. [Google Scholar] [CrossRef]
- Ordoñez, J.C.; Van Bodegom, P.M.; Witte, J.M.; Wright, I.J.; Reich, P.B.; Aerts, R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob. Ecol. Biogeogr. 2009, 18, 137–149. [Google Scholar] [CrossRef]
- Jiang, A.; Jiang, J.; Liu, M. Responses of leaf traits of Sassafras tsumu (Hemsl.) Hemsl. along an altitudinal gradient. Chin. J. Ecol. 2016, 35, 1467–1474. [Google Scholar]
- Pottier, J.; Dubuis, A.; Pellissie, L. The accuracy of plant assemblage prediction from species distribution models varies along environmental gradients. Glob. Ecol. Biogeogr. 2013, 22, 52–63. [Google Scholar] [CrossRef]
- Xia, M.; Wang, X.; Hao, M.; Zhao, X.; Zhang, C. Distribution pattern of functional traits and its response to topographic factors in a conifer and broad-leaved mixed forest in Jiaohe, Jilin province. Acta Ecol. Sin. 2021, 41, 2794–2802. [Google Scholar]
- Zhao, G.; Liu, M.; Shi, P.; Zong, N.; Zhang, X.; Zhang, X. Variation of leaf and root traits and ecological adaptive strategies along a precipitation gradient on Changtang Plateau. Acta Ecol. Sin. 2020, 40, 295–309. [Google Scholar]
- Hulshof, C.M.; Violle, C.; Spasojevic, M.J. Intra-specific and inter-specific variation in specific leaf aea reveal the importance of abiotic divers of species diversity across elevation and latitude. J. Veg. Sci. 2013, 24, 921–931. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Altitude | Sample | Longitude (°) | Latitude (°) | Elevation (m) | Dominant Species |
|---|---|---|---|---|---|
| Low altitude (600–1000 m) | ML | 108.022222 | 25.258333 | 646.32 | Cornus wilsoniana, Lindera communis, Nandina domestica, Selaginella tamariscina |
| HKS | 108.820979 | 27.566865 | 685.11 | Cunninghamia lanceolata, Liquidambar formosana Hance, Symplocos sumuntia, Camellia oleifera | |
| BJS | 106.094444 | 25.174444 | 752.06 | Quercus acutissima, Albizia julibrissin, Arytera littoralis | |
| YTS | 108.116208 | 27.110187 | 835.61 | Liquidambar formosana, Rhus chinensis, Styrax confusus, Pteridium aquilinum | |
| Middle altitude (1000–1400 m) | KY | 107.054326 | 26.908215 | 1095.21 | Cunninghamia lanceolata, Cyclobalanopsis myrsinifolia, Castanea seguinii Dode |
| PG | 104.997222 | 25.033333 | 1152.3 | Cyclobalanopsis myrsinifolia, Cinnamomum glanduliferum, Callicarpa macrophylla Vahl | |
| DSH | 107.576389 | 29.102778 | 1239.5 | Fagus longipetiolata, Cunninghamia lanceolata, Woonyoungia septentrionalis, Synedrella nodiflora | |
| High altitude (1400–1800 m) | KKS | 107.163258 | 28.221634 | 1498.61 | Cunninghamia lanceolata, Fagus longipetiolata Seem, Eurya japonica Thunb |
| NY | 105.3625 | 26.7125 | 1818.32 | Betula luminifera, Lithocarpus glaber |
| Functional Feature Type | Data Type | Attribute |
|---|---|---|
| Leaf thickness (LT) | Numerical value | Average blade thickness (mm) |
| Leaf area (LA) | Numerical value | Average plant leaf area (cm2) |
| Specific leaf area (SLA) | Numerical value | Fresh leaf area/leaf dry mass (cm2/g) |
| Leaf dry matter content (LDMC) | Numerical value | Leaf dry weight/leaf saturated fresh weight (%) |
| Leaf carbon content (LCC) | Numerical value | Plant leaf organic carbon content (g/kg) |
| Leaf nitrogen content (LNC) | Numerical value | Plant leaf total nitrogen content (g/kg) |
| Leaf phosphorus content (LPC) | Numerical value | Plant leaf total phosphorus content (g/kg) |
| Leaf carbon-to-nitrogen ratio (LCN) | Numerical value | Plant leaf carbon content/leaf nitrogen content (%) |
| Leaf nitrogen-to-phosphorus ratio (LNP) | Numerical value | Plant leaf nitrogen content/leaf phosphorus content (%) |
| Type | Factors | Maximum | Minimum | Mean Value | Fold | SD | CV (%) |
|---|---|---|---|---|---|---|---|
| Trait | LT | 0.39 | 0.16 | 0.23 | 2.44 | 0.06 | 26.09 |
| LA | 91.03 | 12.00 | 34.23 | 7.59 | 22.91 | 66.93 | |
| SLA | 359.63 | 102.56 | 204.27 | 3.51 | 73.57 | 36.02 | |
| LDMC | 0.59 | 0.10 | 0.20 | 5.90 | 0.12 | 60.00 | |
| LCC | 87.77 | 42.33 | 67.51 | 2.07 | 10.19 | 15.09 | |
| LNC | 31.16 | 4.05 | 14.71 | 7.69 | 6.70 | 45.55 | |
| LPC | 2.79 | 0.92 | 1.62 | 3.03 | 0.46 | 28.40 | |
| LCN | 19.60 | 2.21 | 6.11 | 8.87 | 4.53 | 74.14 | |
| LNP | 15.59 | 2.01 | 9.32 | 7.76 | 3.37 | 36.16 | |
| Soil | SPMC | 36.71 | 10.68 | 16.83 | 3.44 | 6.15 | 36.51 |
| SOC | 34.83 | 2.69 | 8.44 | 12.95 | 6.30 | 74.68 | |
| SNC | 8.21 | 1.42 | 3.92 | 5.78 | 1.66 | 42.39 | |
| SPC | 2.54 | 0.38 | 0.85 | 6.68 | 0.57 | 66.53 | |
| SCN | 4.66 | 1.59 | 2.07 | 2.93 | 0.73 | 34.98 | |
| SCP | 29.52 | 3.88 | 11.20 | 7.61 | 5.92 | 52.88 | |
| SNP | 12.14 | 2.01 | 5.55 | 6.04 | 2.69 | 48.46 | |
| NJW | 21.55 | 14.56 | 17.63 | 1.48 | 1.99 | 11.26 | |
| NJS | 117.53 | 81.96 | 95.22 | 1.43 | 12.63 | 13.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Deng, Y.; Zhao, H.; Li, F.; Fan, Z.; Tian, T.; Feng, T. Patterns of Change in Plant Leaf Functional Traits Along an Altitudinal Gradient in a Karst Climax Community. Agronomy 2025, 15, 1143. https://doi.org/10.3390/agronomy15051143
Wang Y, Deng Y, Zhao H, Li F, Fan Z, Tian T, Feng T. Patterns of Change in Plant Leaf Functional Traits Along an Altitudinal Gradient in a Karst Climax Community. Agronomy. 2025; 15(5):1143. https://doi.org/10.3390/agronomy15051143
Chicago/Turabian StyleWang, Yang, Ying Deng, Hong Zhao, Fangbing Li, Zuhong Fan, Tian Tian, and Tu Feng. 2025. "Patterns of Change in Plant Leaf Functional Traits Along an Altitudinal Gradient in a Karst Climax Community" Agronomy 15, no. 5: 1143. https://doi.org/10.3390/agronomy15051143
APA StyleWang, Y., Deng, Y., Zhao, H., Li, F., Fan, Z., Tian, T., & Feng, T. (2025). Patterns of Change in Plant Leaf Functional Traits Along an Altitudinal Gradient in a Karst Climax Community. Agronomy, 15(5), 1143. https://doi.org/10.3390/agronomy15051143