


Increased Light Intensity Mitigates CO2 and CH4 Emissions from Paddy Soil by Mediating Iron Redox Cycling Coupled with Organic Carbon Transformation


Abstract
1. Introduction
2. Materials and Methods
2.1. Description of Experimental Soil
2.2. Soil Incubation Experiment
2.3. Sample Analysis
2.4. Data Calculation and Analysis
3. Results
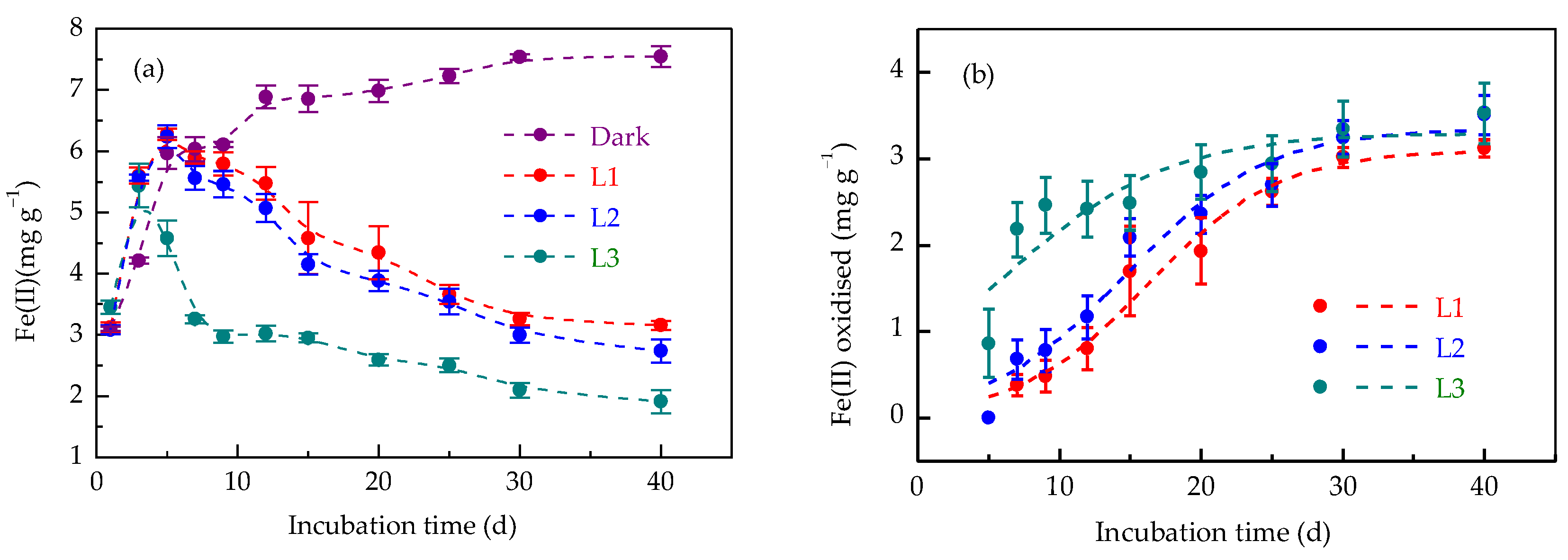
3.1. Responses of Iron Redox Cycling to Light Intensity
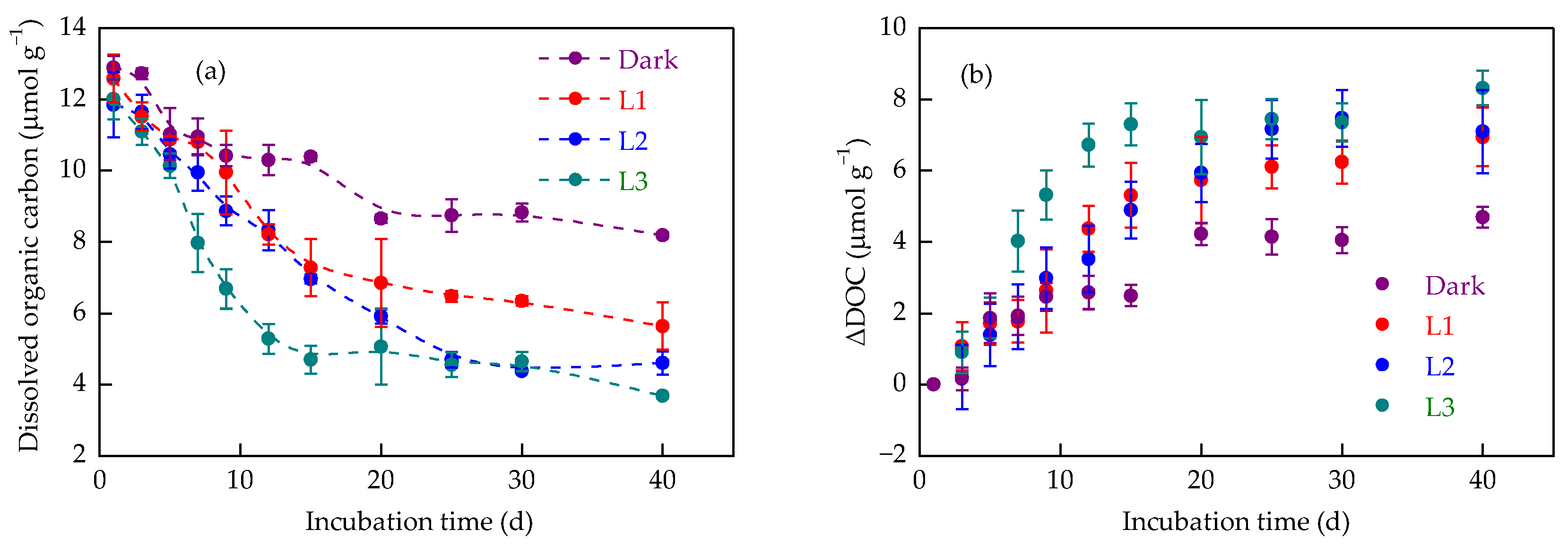
3.2. Changes in Organic Carbon Mineralisation Under Varying Light Intensities
3.3. Dynamics of Dissolved Organic Carbon in Response to Light Intensity
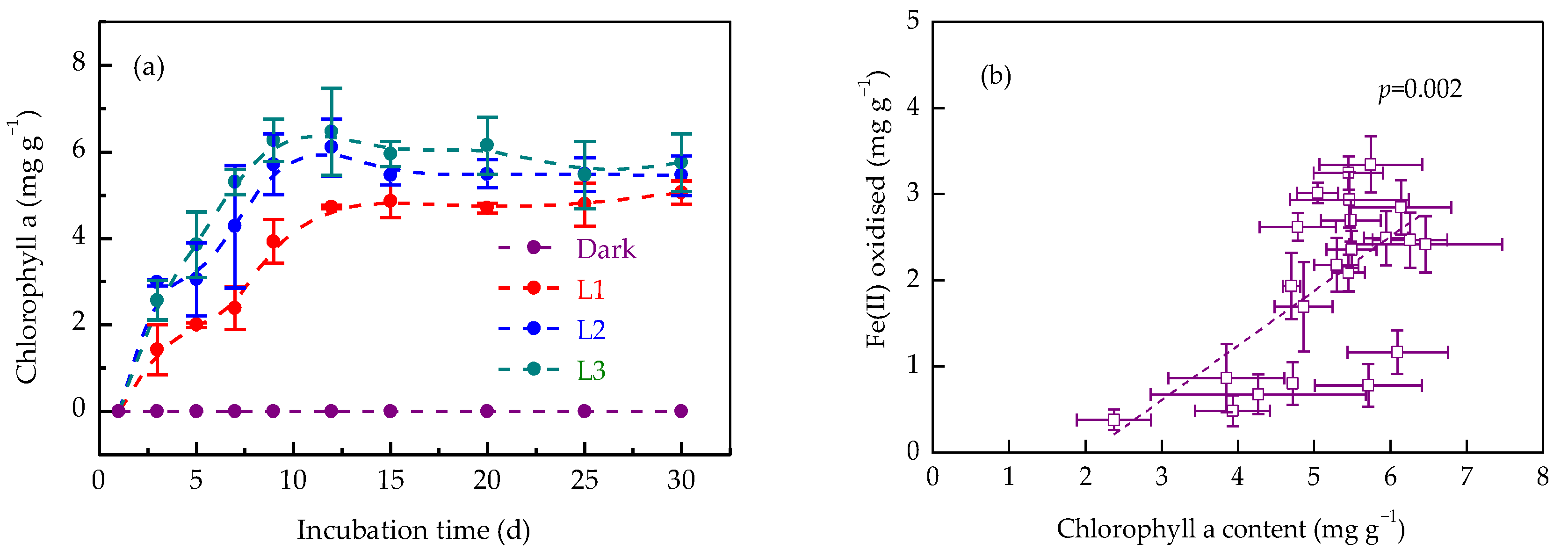
3.4. Effects of Light Intensity on Chlorophyll a Production
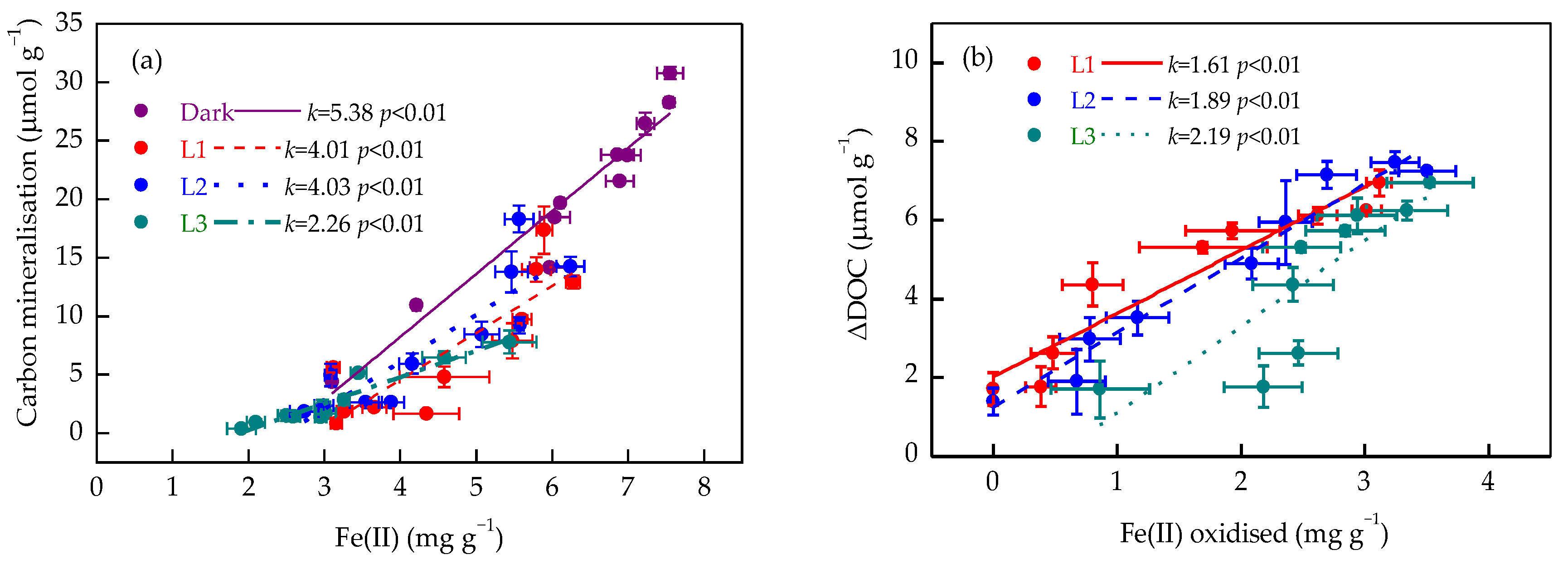
3.5. Relationships Between Iron Redox Processes and Organic Carbon Transformation
4. Discussion
4.1. Light Intensity Modulates Iron Redox Cycling in Paddy Soil
4.2. Iron Redox Cycling Shapes Soil Organic Carbon Dynamics Under Increased Light Intensity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.; Lu, X.; Yu, R.; Sun, H.; Xue, H.; Qi, Z.; Cao, Z.; Zhang, Z.; Liu, T. Greenhouse gases emissions from riparian wetlands: An example from the Inner Mongolia grassland region in China. Biogeosciences 2021, 18, 4855–4872. [Google Scholar] [CrossRef]
- Yu, Z.; Loisel, J.; Brosseau, D.P.; Beilman, D.W.; Hunt, S.J. Global peatland dynamics since the Last Glacial Maximum. Geophys. Res. Lett. 2010, 37, 1–5. [Google Scholar] [CrossRef]
- Lalonde, K.; Mucci, A.; Ouellet, A.; Gélinas, Y. Preservation of organic matter in sediments promoted by iron. Nature 2012, 483, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Holmes, D.E.; Nevin, K.P. Dissimilatory Fe(III) and Mn(IV) reduction. Adv. Microb. Physiol. 2004, 49, 219–286. [Google Scholar] [CrossRef]
- Jaisi, D.P.; Hailiang, D.; Liu, C. Kinetic analysis of microbial reduction of Fe(III) in Nontronite. Environ. Sci. Technol. 2007, 41, 2437–2444. [Google Scholar] [CrossRef]
- Loughlin, E.J. Microbial reduction of geogenic and synthetic goethite and hematite. Minerals 2024, 14, 1086. [Google Scholar] [CrossRef]
- Kostka, J.E.; Wu, J.; Nealson, K.H.; Stucki, J.W. The impact of structural Fe(III) reduction by bacteria on the surface chemistry of smectite clay minerals. Geochim. Cosmochim. Acta 1999, 63, 3705–3713. [Google Scholar] [CrossRef]
- Byrne, J.M.; Klueglein, N.; Pearce, C.; Rosso, K.M.; Appel, E.; Kappler, A. Redox cycling of Fe(II) and Fe(III) in magnetite by Fe-metabolizing bacteria. Science 2015, 347, 1473–1476. [Google Scholar] [CrossRef]
- Kamura, T.; Takai, Y.; Ishikawa, K. Microbial reduction mechanism of ferric iron in paddy soils (Part I). Soil Sci. Plant Nutr. 2012, 9, 5–9. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J. Organic matter mineralization with reduction of ferric iron in anaerobic sediments. Appl. Environ. Microbiol. 1986, 51, 683–689. [Google Scholar] [CrossRef]
- He, J.; Qu, D. Dissimilatory Fe(III) reduction characteristics of paddy soil extract cultures treated with glucose or fatty acids. J. Environ. Sci. 2008, 20, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Buettner, S.W.; Kramer, M.G.; Chadwick, O.A.; Thompson, A. Mobilization of colloidal carbon during iron reduction in basaltic soils. Geoderma 2014, 221–222, 139–145. [Google Scholar] [CrossRef]
- Pan, W.; Kan, J.; Inamdar, S.; Chen, C.; Sparks, D. Dissimilatory microbial iron reduction release DOC (dissolved organic carbon) from carbon-ferrihydrite association. Soil Biol. Biochem. 2016, 103, 232–240. [Google Scholar] [CrossRef]
- Kleber, M.; Bourg, I.C.; Coward, E.K.; Hansel, C.M.; Myneni, S.C.B.; Nunan, N. Dynamic interactions at the mineral–organic matter interface. Nat. Rev. Earth Environ. 2021, 2, 402–421. [Google Scholar] [CrossRef]
- Chen, C.; Hall, S.J.; Coward, E.; Thompson, A. Iron-mediated organic matter decomposition in humid soils can counteract protection. Nat. Commun. 2020, 11, 2255. [Google Scholar] [CrossRef]
- Luo, M.; Zhu, W.; Huang, J.; Liu, Y.; Duan, X.; Wu, J.; Tong, C. Anaerobic organic carbon mineralization in tidal wetlands along a low-level salinity gradient of a subtropical estuary: Rates, pathways, and controls. Geoderma 2019, 337, 1245–1257. [Google Scholar] [CrossRef]
- Widdel, F.; Schnell, S.; Heising, S.; Ehrenreich, A.; Assmus, B.; Schink, B. Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature 1993, 362, 834–836. [Google Scholar] [CrossRef]
- Wang, X.; Sun, L.; Chen, Z.; Guo, D.; Fan, H.; Xu, X.; Shi, Z.; Chen, X. Light inhibition of carbon mineralization associated with iron redox processes in calcareous paddy soil. J. Soils Sediments 2020, 20, 3171–3180. [Google Scholar] [CrossRef]
- Davín, A.A.; Woodcroft, B.J.; Soo, R.M.; Morel, B.; Murali, R.; Schrempf, D.; Clark, J.W.; álvarez-Carretero, S.; Boussau, B.; Moody, E.R.R.; et al. A geological timescale for bacterial evolution and oxygen adaptation. Science 2025, 388, eadp1853. [Google Scholar] [CrossRef]
- Soo, R.M.; Hemp, J.; Parks, D.H.; Fischer, W.W.; Hugenholtz, P. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. Science 2017, 355, 1436. [Google Scholar] [CrossRef]
- Thompson, K.J.; Kenward, P.A.; Bauer, K.W.; Warchola, T.; Gauger, T.; Martinez, R.; Simister, R.L.; Michiels, C.C.; Lliros, M.; Reinhard, C.T.; et al. Photoferrotrophy, deposition of banded iron formations, and methane production in Archean oceans. Sci. Adv. 2019, 5, eaav2869. [Google Scholar] [CrossRef] [PubMed]
- Nikeleit, V.; Maisch, M.; Byrne, J.M.; Harwood, C.; Kappler, A.; Bryce, C. Phototrophic Fe(II) oxidation by Rhodopseudomonas palustris TIE-1 in organic and Fe(II)-rich conditions. Environ. Microbiol. 2024, 26, e16608. [Google Scholar] [CrossRef] [PubMed]
- Hegler, F.; Posth, N.R.; Jiang, J.; Kappler, A. Physiology of phototrophic iron(II)-oxidizing bacteria: Implications for modern and ancient environments. FEMS Microbiol. Ecol. 2008, 66, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583. [Google Scholar] [CrossRef]
- Kushkevych, I.; Procházka, V.; Vítězová, M.; Dordević, D.; Abd El-Salam, M.; Rittmann, S.K.M.R. Anoxygenic photosynthesis with emphasis on green sulfur bacteria and a perspective for hydrogen sulfide detoxification of anoxic environments. Front. Microbiol. 2024, 15, 1417714. [Google Scholar] [CrossRef]
- Ozaki, K.; Thompson, K.J.; Simister, R.L.; Crowe, S.A.; Reinhard, C.T. Anoxygenic photosynthesis and the delayed oxygenation of Earth’s atmosphere. Nat. Commun. 2019, 10, 3026. [Google Scholar] [CrossRef]
- Sun, Y.; Casella, S.; Fang, Y.; Huang, F.; Faulkner, M.; Barrett, S.; Liu, L.N. Light modulates the biosynthesis and organization of cyanobacterial carbon fixation machinery through photosynthetic electron flow. Plant Physiol. 2016, 171, 530–541. [Google Scholar] [CrossRef]
- Bengtsson, M.M.; Wagner, K.; Schwab, C.; Urich, T.; Battin, T.J. Light availability impacts structure and function of phototrophic stream biofilms across domains and trophic levels. Mol. Ecol. 2018, 27, 2913–2925. [Google Scholar] [CrossRef]
- Jodłowska, S.; Sliwińska, S. Effects of light intensity and temperature on the photosynthetic irradiance response curves and chlorophyll fluorescence in three picocyanobacterial strains of Synechococcus. Photosynthetica 2014, 52, 223–232. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Nikeleit, V.; Roth, L.; Maisch, M.; Kappler, A.; Bryce, C. Phototrophic Fe(II) oxidation benefits from light/dark cycles. Environ. Microbiol. Rep. 2024, 16, e13239. [Google Scholar] [CrossRef] [PubMed]
- Melton, E.D.; Schmidt, C.; Kappler, A. Microbial iron(II) oxidation in littoral freshwater lake sediment: The potential for competition between phototrophic vs. nitrate-reducing iron(II)-oxidizers. Front. Microbiol. 2012, 3, 197. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Rothman, D.; Grant, K.; Rosengard, S.; Eglinton, T.; Derry, L.; Galy, V. Mineral protection regulates the long-term global preservation of natural organic carbon. Nature 2019, 570, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Li, L.; Guo, H.; Xie, M. Important role of Fe oxides in global soil carbon stabilization and stocks. Nat. Commun. 2024, 15, 10318. [Google Scholar] [CrossRef]
- Eusterhues, K.; Rennert, T.; Knicker, H.; Kögel-Knabner, I.; Totsche, K.U.; Schwertmann, U. Fractionation of organic matter due to reaction with ferrihydrite: Coprecipitation versus adsorption. Environ. Sci. Technol. 2011, 45, 527–533. [Google Scholar] [CrossRef]
- Muehe, E.M.; Obst, M.; Hitchcock, A.; Tyliszczak, T.; Behrens, S.; Schröder, C.; Byrne, J.M.; Michel, F.M.; Krämer, U.; Kappler, A. Fate of Cd during microbial Fe(III) mineral reduction by a novel and Cd-tolerant geobacter species. Environ. Sci. Technol. 2013, 47, 14099–14109. [Google Scholar] [CrossRef]
- Glodowska, M.; Stopelli, E.; Schneider, M.; Lightfoot, A.; Rathi, B.; Straub, D.; Patzner, M.; Duyen, V.T.; Berg, M.; Kleindienst, S.; et al. Role of in situ natural organic matter in mobilizing As during microbial reduction of FeIII-Mineral-Bearing Aquifer Sediments from Hanoi (Vietnam). Environ. Sci. Technol. 2020, 54, 4149–4159. [Google Scholar] [CrossRef]
- Qiu, G.; Koedooder, C.; Qiu, B.; Shaked, Y.; Keren, N. Iron transport in cyanobacteria—From molecules to communities. Trends Microbiol. 2022, 30, 309. [Google Scholar] [CrossRef]
- Molot, L.A.; Li, G.; Findlay, D.L.; Watson, S.B. Iron-mediated suppression of bloom-forming cyanobacteria by oxine in a eutrophic lake. Freshw. Biol. 2010, 55, 1102–1117. [Google Scholar] [CrossRef]
- Tania, L.; Grace, M.W.; Elizabeth, D.S. Iron availability allows sustained cyanobacterial growth: A dual-lake case study. Inland Waters 2021, 11, 417–429. [Google Scholar] [CrossRef]
- Lu, R. Analytical Method of Soil Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 2000; pp. 13, 47–55, 65, 107–108. (In Chinese) [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Saviozzi, A.; Vanni, G.; Cardelli, R. Carbon mineralization kinetics in soils under urban environment. Appl. Soil Ecol. 2014, 73, 64–69. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, X.; Sun, L.; Dong, L.; Guo, D.; Shi, Z. Nitrate-dependent and photosynthetic Fe(II) oxidation processes in a calcareous paddy soil. Acta Pedol. Sin. 2023, 60, 127–137. (In Chinese) [Google Scholar] [CrossRef]
- Hamilton, T.L.; Bennett, A.C.; Murugapiran, S.K.; Havig, J.R. Anoxygenic phototrophs span geochemical gradients and diverse morphologies in terrestrial geothermal springs. Msystems 2019, 4, e00498-19. [Google Scholar] [CrossRef]
- Melton, E.D.; Swanner, E.D.; Behrens, S.; Schmidt, C.; Kappler, A. The interplay of microbially mediated and abiotic reactions in the biogeochemical Fe cycle. Nat. Rev. Microbiol. 2014, 12, 797–808. [Google Scholar] [CrossRef]
- Hamilton, T.L. The trouble with oxygen: The ecophysiology of extant phototrophs and implications for the evolution of oxygenic photosynthesis. Free Radic. Biol. Med. 2019, 140, 233–249. [Google Scholar] [CrossRef]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Xia, Y. Practical Manual for Laboratory Technicians; Chemical Industry Press: Beijing, China, 2015; pp. 208–209. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Final Fe(II) Oxidation (mg g−1) | Oxidation Capacity, a (mg g−1) | Oxidation Rate Constant, k (d−1) | Correlation Coefficient, R2 | Probability of Significance, p | Number of Samples Fitted, n |
|---|---|---|---|---|---|---|
| L1 | 3.12 ± 0.10 aA | 3.10 ± 0.17 aA | 0.22 ± 0.04 b | 0.97 | <0.001 | 9 |
| L2 | 3.50 ± 0.23 aA | 3.35 ± 0.23 aA | 0.20 ± 0.04 b | 0.95 | <0.001 | 9 |
| L3 | 3.53 ± 0.35 aA | 3.29 ± 0.29 aA | 0.87 ± 0.35 a | 0.74 | <0.001 | 10 |
| Treatment | Decrease Rate Constant, k, d−1 | Correlation Coefficient, R2 | Probability of Significance, p | Number of Samples Fitted, n |
|---|---|---|---|---|
| L1 | 0.14 ± 0.02 b | 0.99 | <0.001 | 8 |
| L2 | 0.18 ± 0.03 b | 0.98 | <0.001 | 8 |
| L3 | 0.26 ± 0.05 a | 0.94 | <0.001 | 10 |
| Treatment | Initial DOC Concentration, C0 (µmol g−1) | Decrease Rate Constant, k, d−1 | Correlation Coefficient, R2 | Probability of Significance, p | Number of Samples Fitted, n |
|---|---|---|---|---|---|
| Dark | 12.30 ± 0.35 a | 0.012 ± 0.002 d | 0.84 | <0.001 | 11 |
| L1 | 12.29 ± 0.44 a | 0.024 ± 0.003 c | 0.91 | <0.001 | 11 |
| L2 | 12.35 ± 0.38 a | 0.034 ± 0.003 b | 0.96 | <0.001 | 11 |
| L3 | 11.61 ± 0.86 a | 0.042 ± 0.007 a | 0.83 | <0.001 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Jiang, M.; Li, M.; Wang, X.; Han, Y.; Chen, X. Increased Light Intensity Mitigates CO2 and CH4 Emissions from Paddy Soil by Mediating Iron Redox Cycling Coupled with Organic Carbon Transformation. Agronomy 2025, 15, 1137. https://doi.org/10.3390/agronomy15051137
Sun L, Jiang M, Li M, Wang X, Han Y, Chen X. Increased Light Intensity Mitigates CO2 and CH4 Emissions from Paddy Soil by Mediating Iron Redox Cycling Coupled with Organic Carbon Transformation. Agronomy. 2025; 15(5):1137. https://doi.org/10.3390/agronomy15051137
Chicago/Turabian StyleSun, Lirong, Mengxiao Jiang, Meng Li, Xugang Wang, Yafeng Han, and Xianni Chen. 2025. "Increased Light Intensity Mitigates CO2 and CH4 Emissions from Paddy Soil by Mediating Iron Redox Cycling Coupled with Organic Carbon Transformation" Agronomy 15, no. 5: 1137. https://doi.org/10.3390/agronomy15051137
APA StyleSun, L., Jiang, M., Li, M., Wang, X., Han, Y., & Chen, X. (2025). Increased Light Intensity Mitigates CO2 and CH4 Emissions from Paddy Soil by Mediating Iron Redox Cycling Coupled with Organic Carbon Transformation. Agronomy, 15(5), 1137. https://doi.org/10.3390/agronomy15051137