
Impact of Phytoplankton Biomass on the Growth and Development of Agricultural Plants

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions, Treatments, and Composition of Active Substances
2.2. Determination of Proline Content
2.3. Determination of Hydrogen Peroxide and Malondialdehyde Content
2.4. Determination of Total Phenol Content
2.5. Antioxidant Activity Determination
2.6. Statistical Analysis
3. Results
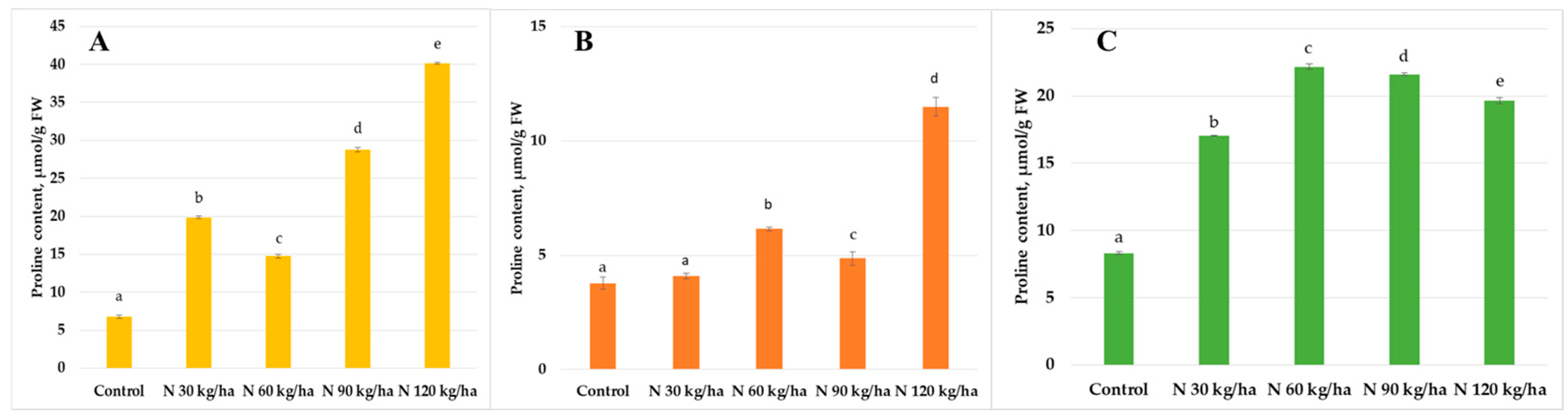
3.1. Proline Content
3.2. Hydrogen Peroxide Content
3.3. MDA Content
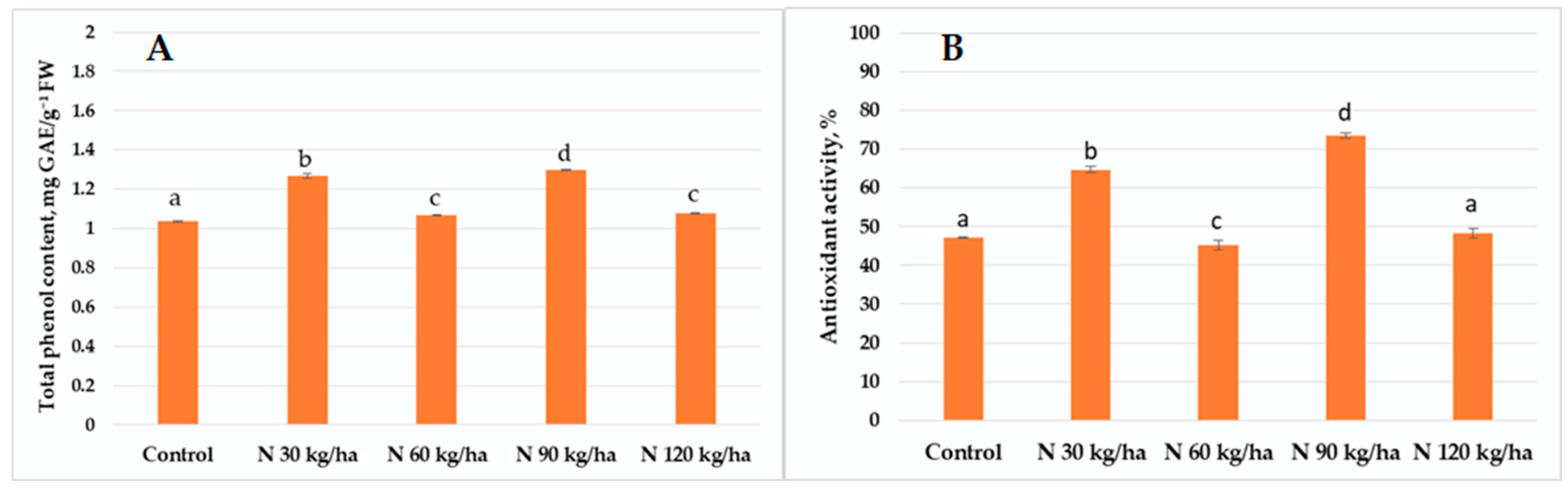
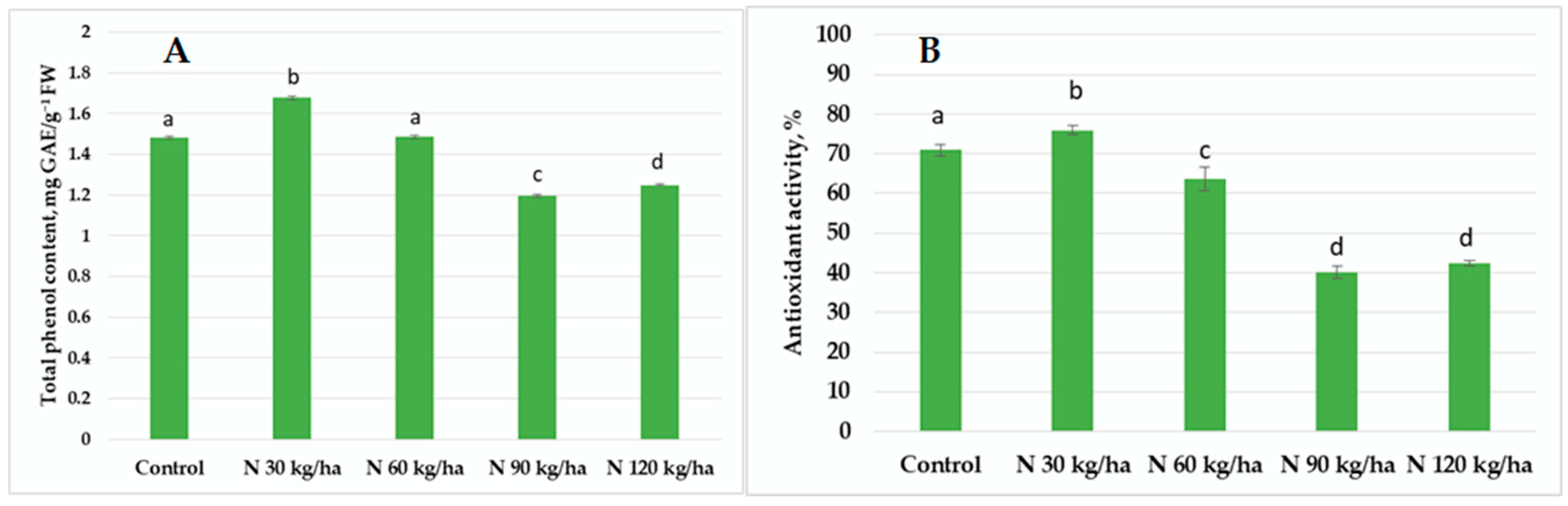
3.4. Total Phenol Content and Antioxidant Activity
4. Discussion
4.1. Proline Content Changes Under Various Phytoplankton Biomass Additions
4.2. Influence of Different Amounts of Phytoplankton Biomass on Hydrogen Peroxide and Malondialdehyde Content
4.3. Activity of the Non-Enzymatic Plant Defence System Under Various Phytoplankton Biomass Additions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chakraborty, S.; Tiwari, P.K.; Sasmal, S.K.; Misra, A.K.; Chattopadhyay, J. Effects of fertilizers used in agricultural fields on algal blooms. Eur. Phys. J. Spec. Top. 2017, 226, 2119–2133. [Google Scholar] [CrossRef]
- Zaib, M.; Zubair, M.; Aryan, M.; Abdullah, M.; Manzoor, S.; Masood, F.; Saeed, S. A review on challenge and opportunities of fertilizer use efficiency and their role in sustainable agriculture with future prospect and recommendations. Curr. Res. Agric. Farming 2023, 4, 1–14. [Google Scholar] [CrossRef]
- Sharma, A.; Chetani, R. A review on the effect of organic and chemical fertilizers on plants. Int. J. Res. Appl. Sci. Eng. Technol. 2017, 5, 677–680. [Google Scholar] [CrossRef]
- Ciero, L. Role of inorganic fertilizers in modern agriculture: Nourishing plants with minerals. Int. J. Manures Fertil. 2023, 11, 1–2. [Google Scholar]
- Verma, B.C.; Pramanik, P.; Bhaduri, D. Organic fertilizers for sustainable soil and environmental management. In Nutrient Dynamics for Sustainable Crop Production; Meena, R.S., Ed.; Springer: Singapore, 2020; pp. 289–313. [Google Scholar] [CrossRef]
- Chittora, D.; Parveen, T.; Yadav, J.; Meena, B.R.; Jain, T.; Sharma, K. Harmful impact of synthetic fertilizers on growing agriculture and environment. Glob. J. Pharmaceu. Sci. 2023, 11, 555804. [Google Scholar] [CrossRef]
- Shanmugavel, D.; Rusyn, I.; Solorza-Feria, O.; Kamaraj, S.-K. Sustainable SMART fertilizers in agriculture systems: A review on fundamentals to in-field applications. Sci. Total Environ. 2023, 904, 166729. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Hasnain, S. Phytostimulation and biofertilization in wheat by cyanobacteria. J. Ind. Microbiol. Biotechnol. 2011, 38, 85–92. [Google Scholar] [CrossRef]
- Mazhar, S.; Hasnain, S. Screening of native plant growth promoting cyanobacteria and their impact and Triticum aestivum var. Uqab 2000 growth. Afr. J. Agric. Res. 2011, 6, 3988–3993. [Google Scholar] [CrossRef]
- Eurostat. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Agri-environmental_indicator_-_mineral_fertiliser_consumption#Analysis_at_country_level (accessed on 2 January 2025).
- European Commission. Available online: https://commission.europa.eu/strategy-and-policy/priorities-2019-2024/european-green-deal_en (accessed on 18 January 2025).
- Gustafsson, B.G.; Schenk, F.; Blenckner, T.; Eilola, K.; Meier, H.E.M.; Müller-Karulis, B.; Neumann, T.; Ruoho-Airola, T.; Savchuk, O.P.; Zorita, E. Reconstructing the development of Baltic Sea eutrophication. AMBIO 2012, 41, 534–548. [Google Scholar] [CrossRef]
- Lønborg, C.; Markager, S. Nitrogen in the Baltic Sea: Long-term trends, a budget and decadal time lags in responses to declining inputs. Estuar. Coast. Shelf Sci. 2021, 261, 107529. [Google Scholar] [CrossRef]
- Balasuriya, B.T.G.; Ghose, A.; Gheewala, S.H.; Prapaspongsa, T. Assessment of eutrophication potential from fertiliser application in agricultural systems in Thailand. Sci. Total Environ. 2022, 833, 154993. [Google Scholar] [CrossRef] [PubMed]
- Stakėnienė, R.; Jokšas, K.; Kriaučiūnienė, J.; Jakimavičius, D.; Raudonytė-Svirbutavičienė, E. Nutrient loadings and exchange between the curonian Lagoon and the Baltic Sea: Changes over the past two decades (2001–2020). Water 2023, 15, 4096. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Pliński, M. Do toxic cyanobacteria blooms pose a threat to the Baltic ecosystem? Oceanologia 2009, 51, 293–319. [Google Scholar] [CrossRef]
- Sanseverino, I.; Conduto António, D.S.; Loos, R.; Lettieri, T. Cyanotoxins: Methods and Approaches for Their Analysis and Detection; Publications Office of the European Union: Luxembourg, 2017; p. 70. [Google Scholar] [CrossRef]
- Lyon-Colbert, A.; Su, S.; Cude, C. A Systematic literature review for evidence of Aphanizomenon flos-aquae toxigenicity in recreational waters and toxicity of dietary supplements: 2000–2017. Toxins 2018, 10, 254. [Google Scholar] [CrossRef]
- Agarwal, P.; Soni, R.; Kaur, P.; Madan, A.; Mishra, R.; Pandey, J.; Singh, S.; Singh, G. Cyanobacteria as a Promising Alternative for Sustainable Environment: Synthesis of Biofuel and Biodegradable Plastics. Front. Microbiol. 2022, 13, 939347. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Dong, W.; Zhang, X.; Tyagi, R.D.; Drogui, P.; Surampalli, R.Y. The potential of microalgae in biodiesel production. Renew. Sustain. Energy Rev. 2018, 90, 336–346. [Google Scholar] [CrossRef]
- Sitther, V.; Tabatabai, B.; Fathabad, S.G.; Gichuki, S.; Chen, H.; Christy, A.; Arumanayagam, S. Cyanobacteria as a biofuel source: Advances and applications. In Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press: London, UK; Elsevier: Amsterdam, The Netherlands, 2020; pp. 269–289. [Google Scholar] [CrossRef]
- Arias, D.M.; Ortíz-Sánchez, E.; Okoye, P.U.; Rodríguez-Rangel, H.; Ortega, A.B.; Longoria, A.; Domínguez-Espíndola, R.; Sebastian, P.J. A review on cyanobacteria cultivation for carbohydrate-based biofuels: Cultivation aspects, polysaccharides accumulation strategies, and biofuels production scenarios. Sci. Total Environ. 2021, 794, 148636. [Google Scholar] [CrossRef]
- Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P.; et al. Biomolecules from microalgae and cyanobacteria: Applications and market survey. Appl. Sci. 2022, 12, 1924. [Google Scholar] [CrossRef]
- Scoglio, G.; Jackson, H.O.; Purton, S. The commercial potential of Aphanizomenon flosaquae, a nitrogenfixing edible cyanobacterium. J. Appl. Phycol. 2024, 36, 1593–1617. [Google Scholar] [CrossRef]
- Mader, P.; Kaiser, F.; Adholeya, A.; Singh, R.; Uppal, H.S.; Sharma, A.K.; Srivastava, R.; Sahai, V.; Aragno, M.; Wiemken, A. Inoculation of root microorganisms for sustainable wheat-rice and wheat-black gram rotations in India. Soil Biol. Biochem. 2011, 43, 609–619. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Asif, M.; Zaheer, A.; Malik, A.; Qasim, A.Q.; Rasool, M. Plant growth promoting rhizobacteria and sustainable agriculture: A review. Afr. J. Microbiol. Res. 2013, 7, 704–709. [Google Scholar] [CrossRef]
- Chittora, D.; Meena, M.; Barupal, T.; Swapnil, P.; Sharma, K. Cyanobacteria as a source of biofertilizers for sustainable agriculture. Biochem. Biophys. Rep. 2020, 22, 100737. [Google Scholar] [CrossRef]
- Prasanna, R.; Babu, S.; Rana, A.; Kabi, S.R.; Chaudhary, V.; Gupta, V.; Kumar, A.; Shivay, Y.S.; Nain, L.; Pal, R.K. Evaluating the establishment and agronomic proficiency of cyanobacterial consortia as organic options in wheat–rice cropping sequence. Exp. Agric. 2013, 49, 416–434. [Google Scholar] [CrossRef]
- Jankauskienė, J.; Buzytė, K.; Paškauskas, R. Melsvabakterių biomasės kaip biotrąšos naudojimo galimybės žemės ūkio augalų augimo ir vystymosi procesams valdyti. In Proceedings of the Conference for Junior Researchers “Science–Future of Lithuania”, Aplinkos Apsaugos Inžinerija/Environmental Protection Engineering, Vilnius, Lithuania, 19 March 2021; Volume 24, pp. 71–78. [Google Scholar] [CrossRef]
- Komárek, J.; Komárková, J. Diversity of Aphanizomenon-like cyanobacteria. Czech Phycol. 2006, 6, 1–32. [Google Scholar]
- Komárek, J. Cyanoprokaryota 3. Teil: Heterocytous Genera. In Süßwasserflora Von Mitteleuropa Bd. 19/3; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spectrum: Berlin/Heidelberg, Germany, 2013; p. 1130. [Google Scholar]
- Hawkesford, M.; Riche, A.B. Impacts of G × E × M on nitrogen use efficiency in wheat and future prospects. Front. Plant Sci. 2020, 11, 1157. [Google Scholar] [CrossRef]
- Meier, U. Growth stages of mono and dicotyledonous plants. In BBCH Monograph; Meier, U., Ed.; Julius Kühn-Institut: Quedlinburg, Germany, 2018; p. 564. [Google Scholar] [CrossRef]
- Mattioli, R.; Falasca, G.; Sabatini, S.; Altamura, M.M.; Costantino, P.; Trovato, M. The proline biosynthetic genes P5CS1 and P5CS2 play overlapping roles in Arabidopsis flower transition but not in embryo development. Physiol. Plant 2009, 137, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M.W. Short communication proline accumulation is a general response to abiotic stress in the date palm tree (Phoenix dactylifera L.). Genet. Mol. Res. 2015, 14, 9943–9950. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Kong, W.; Liu, F.; Zhang, C.; Feng, H. Non-destructive determination of Malondialdehyde (MDA) distribution in oilseed rape leaves by laboratory scale NIR hyperspectral imaging. Sci. Rep. 2016, 6, 35393. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.; Norman, H.; Krizek, D.; Mirecki, R. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Zagoskina, N.V.; Zubova, M.Y.; Nechaeva, T.L.; Kazantseva, V.V.; Goncharuk, E.A.; Katanskaya, V.M.; Baranova, E.N.; Aksenova, M.A. Polyphenols in plants: Structure, biosynthesis, abiotic stress regulation, and practical applications (Review). Int. J. Mol. Sci. 2023, 24, 13874. [Google Scholar] [CrossRef] [PubMed]
- Swain, T.; Goldstein, L. Methods in Polyphenol Chemistry; Pridham, J.B., Ed.; Pergamon Press: Oxford, UK, 1964; pp. 131–146. [Google Scholar]
- Ali, A.; Alqurainy, F.; Motohashi, N. Activities of antioxidants in plants under environmental stress. Environ. Sci. Biol. 2006, 1–32. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Karthikeyan, N.; Prasanna, R.; Sood, A.; Jaiswal, P.; Nayak, S.; Kaushik, B.D. Physiological characterization and electron microscopic investigation of cyanobacteria associated with wheat rhizosphere. Folia Microbiol. 2009, 54, 43–51. [Google Scholar] [CrossRef]
- Maqubela, M.P.; Mnkeni, P.N.S.; Malam Issa, O.; Pardo, M.T.; D’Acqui, L.P. Nostoc cyanobacterial inoculation in South African agricultural soils enhances soil structure, fertility, and maize growth. Plant Soil 2009, 315, 79–92. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Smith, S.M. Hormone Metabolism and Signaling in Plants, 1st ed.; Academic Press: London, UK; Elsevier: Amsterdam, The Netherlands, 2017; p. 597. [Google Scholar]
- Řezanka, T.; Palyzová, A.; Sigler, K. Isolation and identification of siderophores produced by cyanobacteria. Folia Microbiol. 2018, 63, 569–579. [Google Scholar] [CrossRef]
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable sources of plant biostimulation: Microalgae as a sustainable means to improve crop performance. Front. Plant Sci. 2018, 9, 1782. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Wood, E.E.; Llewellyn, C.A. Algae biostimulants: A critical look at microalgal biostimulants for sustainable agricultural practices. Biotechnol. Adv. 2021, 49, 107754. [Google Scholar] [CrossRef] [PubMed]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643. [Google Scholar] [CrossRef] [PubMed]
- Gavelienė, V.; Jurkonienė, S. Probiotics enhance cereal yield and quality and modify agrochemical soil properties. Microorganisms 2022, 10, 1277. [Google Scholar] [CrossRef] [PubMed]
- Moradbeygi, H.; Jamei, R.; Heidari, R.; Darvishzadeh, R. Investigating the enzymatic and non-enzymatic antioxidant defense by applying iron oxide nanoparticles in Dracocephalum moldavica L. plant under salinity stress. Sci. Hortic. 2020, 272, 109537. [Google Scholar] [CrossRef]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. A review. Agron. Sustain. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Renzetti, M.; Funck, D.; Trovato, M. Proline and ROS: A unified mechanism in plant development and stress response? Plants 2024, 14, 2. [Google Scholar] [CrossRef]
- El-Arroussi, H.; Elbaouchi, A.; Benhima, R.; Bendaou, N.; Smouni, A.; Wahby, I. Halophilic microalgae Dunaliella salina extracts improve seed germination and seedling growth of Triticum aestivum L. under salt stress. Acta Hortic. 2016, 1148, 13–26. [Google Scholar] [CrossRef]
- Brito, Â.; Rocha, M.; Kaštovský, J.; Viera, J.; Viera, C.P.; Ramos, V.; Correia, M.; Santos, M.; Mota, R.; Roque, J.; et al. A new cyanobacterial species with a protective effect on lettuce grown under salinity stress: Envisaging sustainable agriculture practices. J. Appl. Phycol. 2022, 34, 915–928. [Google Scholar] [CrossRef]
- Jajić, I.; Sarna, T.; Szewczyk, G.; Strzałka, K. Changes in production of reactive oxygen species in illuminated thylakoids isolated during development and senescence of barley. J. Plant Physiol. 2015, 184, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Hill, A.L.; Shirsekar, G.; Afzal, A.J.; Wang, G.-L.; Mackey, D.; Bonello, P. Quantification of hydrogen peroxide in plant tissues using Amplex Red. Methods 2016, 109, 105–113. [Google Scholar] [CrossRef]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Yang, S.L.; Lan, S.S.; Gong, M. Hydrogen peroxide-induced proline and metabolic pathway of its accumulation in maize seedlings. J. Plant Physiol. 2009, 166, 1694–1699. [Google Scholar] [CrossRef]
- Amarala, J.S.; Seabra, R.M.; Andradea, P.B.; Valentaoa, P.; Pereirac, L.A.; Ferreresd, F. Phenolic profile in the quality control of walnut (Juglans regia L.) leaves. Food Chem. 2004, 88, 373–379. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Extractios and analysis of phenolics in food. J. Chromatogr. 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Cazzonelli, I. Carotenoids in nature: Insights from plants and beyond. Funct. Plant Biol. 2011, 38, 833–847. [Google Scholar] [CrossRef]
- Battke, F.; Ernst, D.; Halbach, S. Ascorbate promotes mercury vapour emission from plants. Plant Cell Environ. 2005, 28, 1487–1495. [Google Scholar] [CrossRef]
- Podsedek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT—Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Yandigeri, M.S.; Arora, D.K. Cyanobacteria-mediated phenylpropanoids and phytohormones in rice (Oryza sativa) enhance plant growth and stress tolerance. Antonie Van Leeuwenhoek 2011, 100, 557–568. [Google Scholar] [CrossRef] [PubMed]
- El Arroussi, H.; Benhima, R.; El-Baouchi, A.; Sijilmassi, B.; El Mernissi, N.; Aafsar, A.; Meftah- Kadmiri, I.; Bendaou, N.; Smouni, A. Dunaliella salina exopolysaccharides: A promising biostimulant for salt stress tolerance in tomato (Solanum lycopersicum). J. Appl. Phycol. 2018, 30, 2929–2941. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Treatments | Control | N 30 kg/ha | N 60 kg/ha | N 90 kg/ha | N 120 kg/ha | F Statistic | p Value |
|---|---|---|---|---|---|---|---|---|
| Oilseed rape | AVERAGE | 1.21 a * | 0.68 b | 0.80 c | 0.90 d | 2.14 e | 3503.4 | 0.000 |
| SD | 0.01 | 0.02 | 0.01 | 0.01 | 0.01 | |||
| Common wheat | AVERAGE | 0.03 a | 0.18 b | 0.09 c | 0.54 d | 0.33 e | 203.5 | 0.000 |
| SD | 0.01 | 0.01 | 0.02 | 0.03 | 0.01 | |||
| Pea | AVERAGE | 0.31 a | 0.70 b | 0.50 c | 0.45 d | 0.58 e | 1677.1 | 0.000 |
| SD | 0.01 | 0.01 | 0.01 | 0.01 | 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankauskienė, J.; Janušaitienė, K.; Karosienė, J.; Paškauskas, R.; Jurkonienė, S.; Mockevičiūtė, R. Impact of Phytoplankton Biomass on the Growth and Development of Agricultural Plants. Agronomy 2025, 15, 1120. https://doi.org/10.3390/agronomy15051120
Jankauskienė J, Janušaitienė K, Karosienė J, Paškauskas R, Jurkonienė S, Mockevičiūtė R. Impact of Phytoplankton Biomass on the Growth and Development of Agricultural Plants. Agronomy. 2025; 15(5):1120. https://doi.org/10.3390/agronomy15051120
Chicago/Turabian StyleJankauskienė, Jurga, Kornelija Janušaitienė, Jūratė Karosienė, Ričardas Paškauskas, Sigita Jurkonienė, and Rima Mockevičiūtė. 2025. "Impact of Phytoplankton Biomass on the Growth and Development of Agricultural Plants" Agronomy 15, no. 5: 1120. https://doi.org/10.3390/agronomy15051120
APA StyleJankauskienė, J., Janušaitienė, K., Karosienė, J., Paškauskas, R., Jurkonienė, S., & Mockevičiūtė, R. (2025). Impact of Phytoplankton Biomass on the Growth and Development of Agricultural Plants. Agronomy, 15(5), 1120. https://doi.org/10.3390/agronomy15051120