
Enhancing Black Soil Fertility and Microbial Community Structure via Microbial Agents to Reduce Chemical Fertilizer Dependency: A Strategy to Boost Maize Yield

Abstract

1. Introduction
2. Materials and Methods
2.1. Field Experiment Site and Design
2.2. Sample Preparation and Analyses
2.3. Advanced Absolute Quantification 16S-Seq
2.4. Statistical Analyses
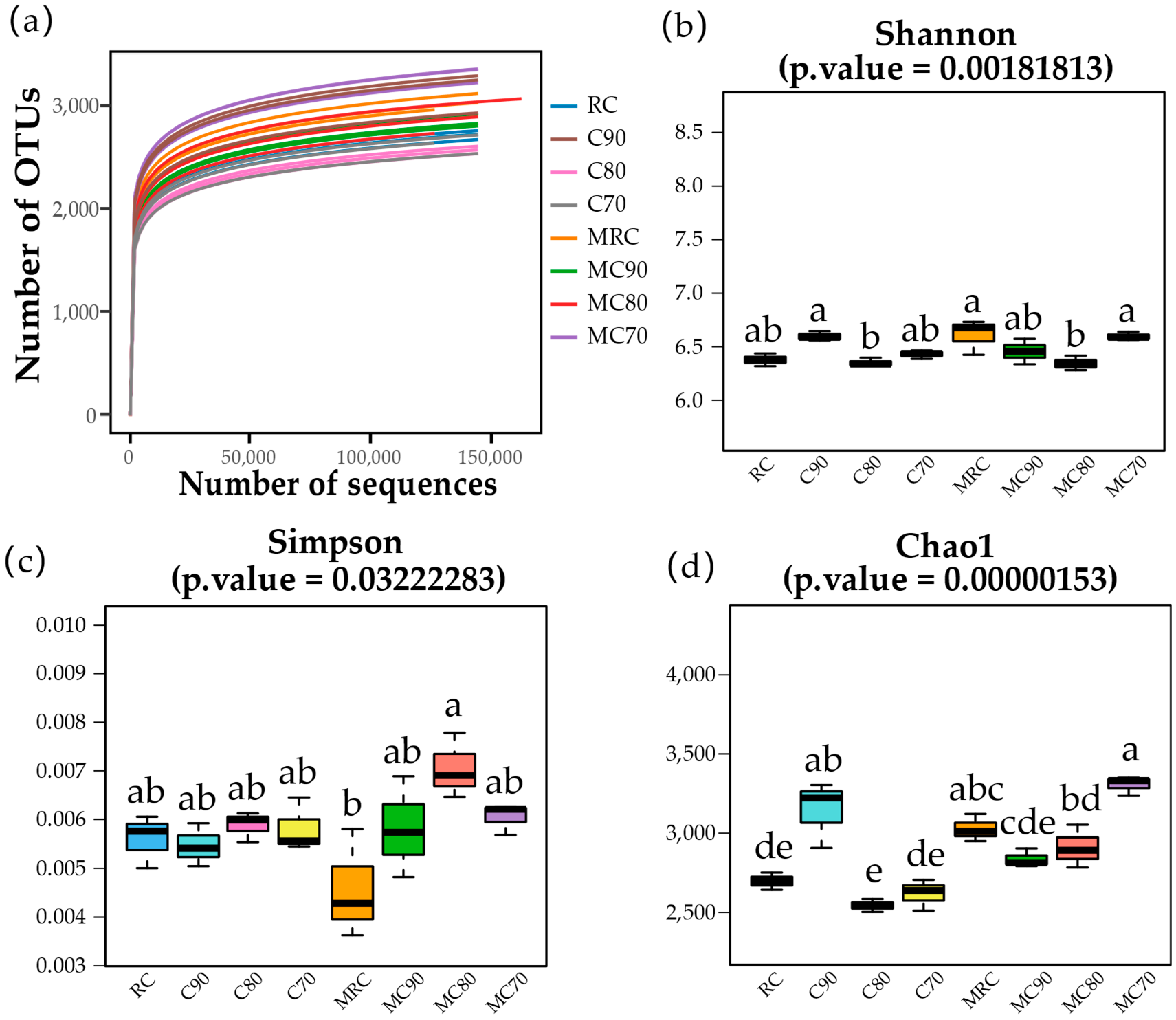
2.4.1. Alpha Diversity
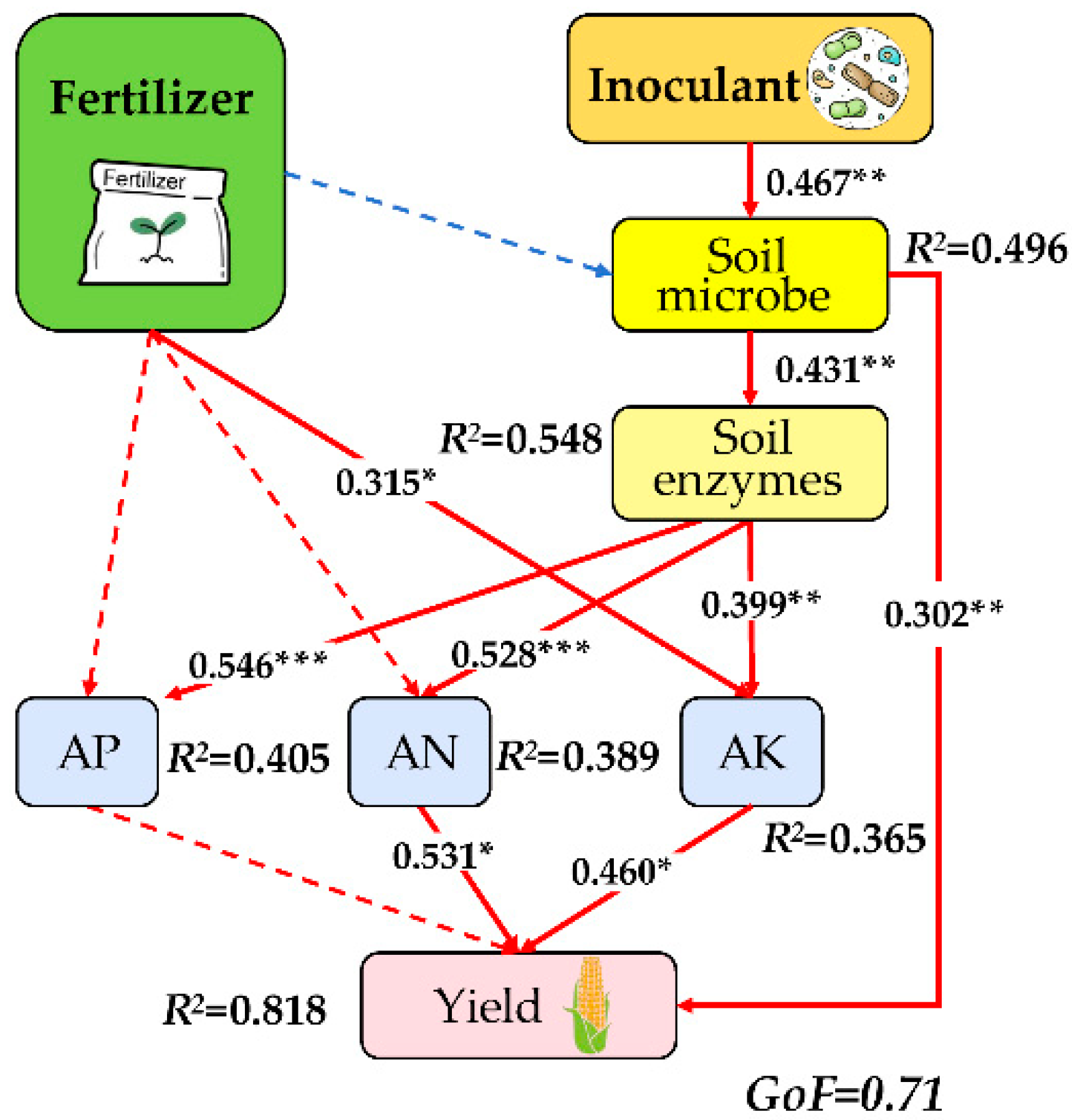
2.4.2. PLS-PM (Partial Least Squares Path Model) Analysis
2.4.3. Principal Component Analysis
2.4.4. Difference Analysis
3. Results
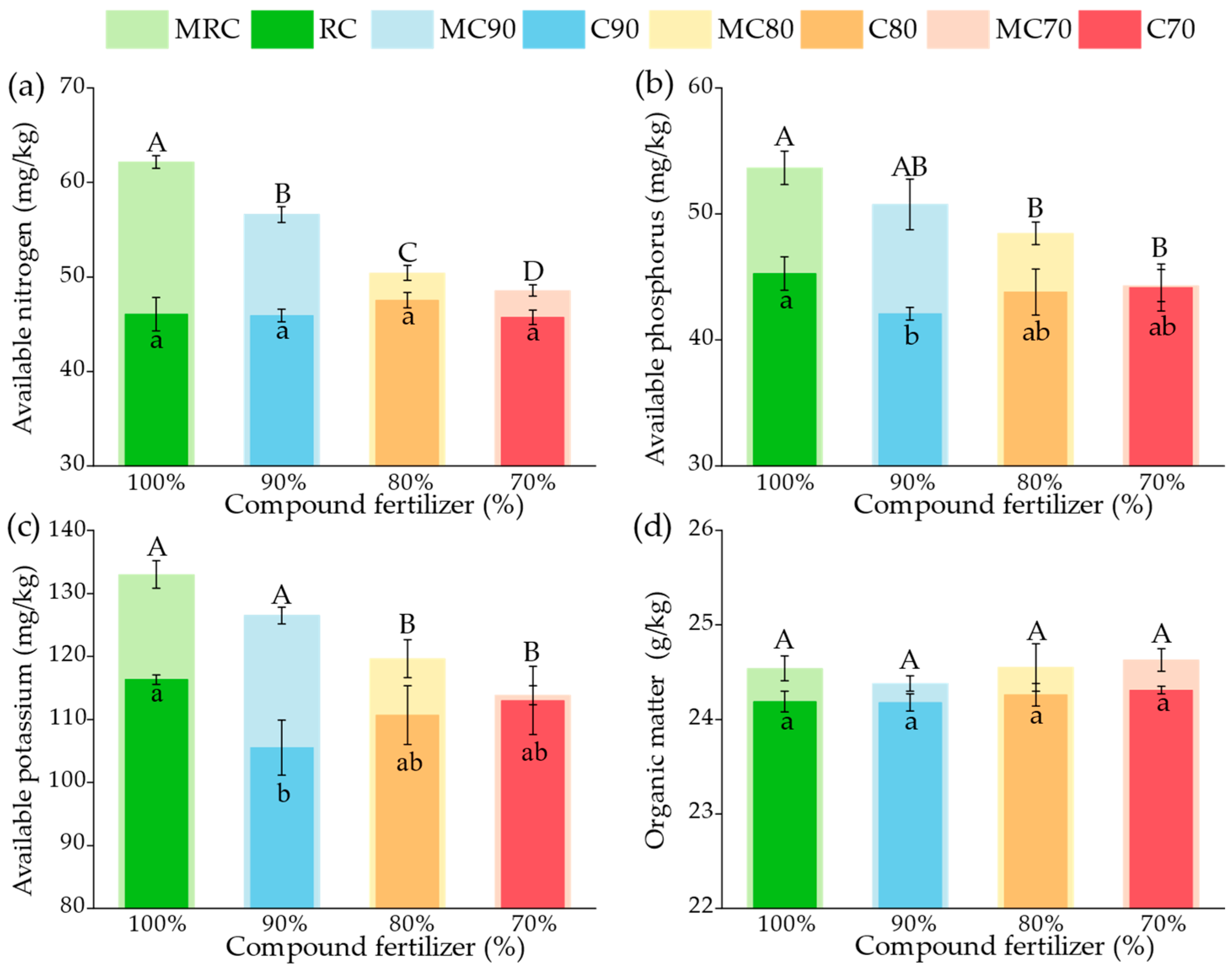
3.1. Soil Physicochemical Properties
3.2. Absolute Quantification of Soil Microbial Community
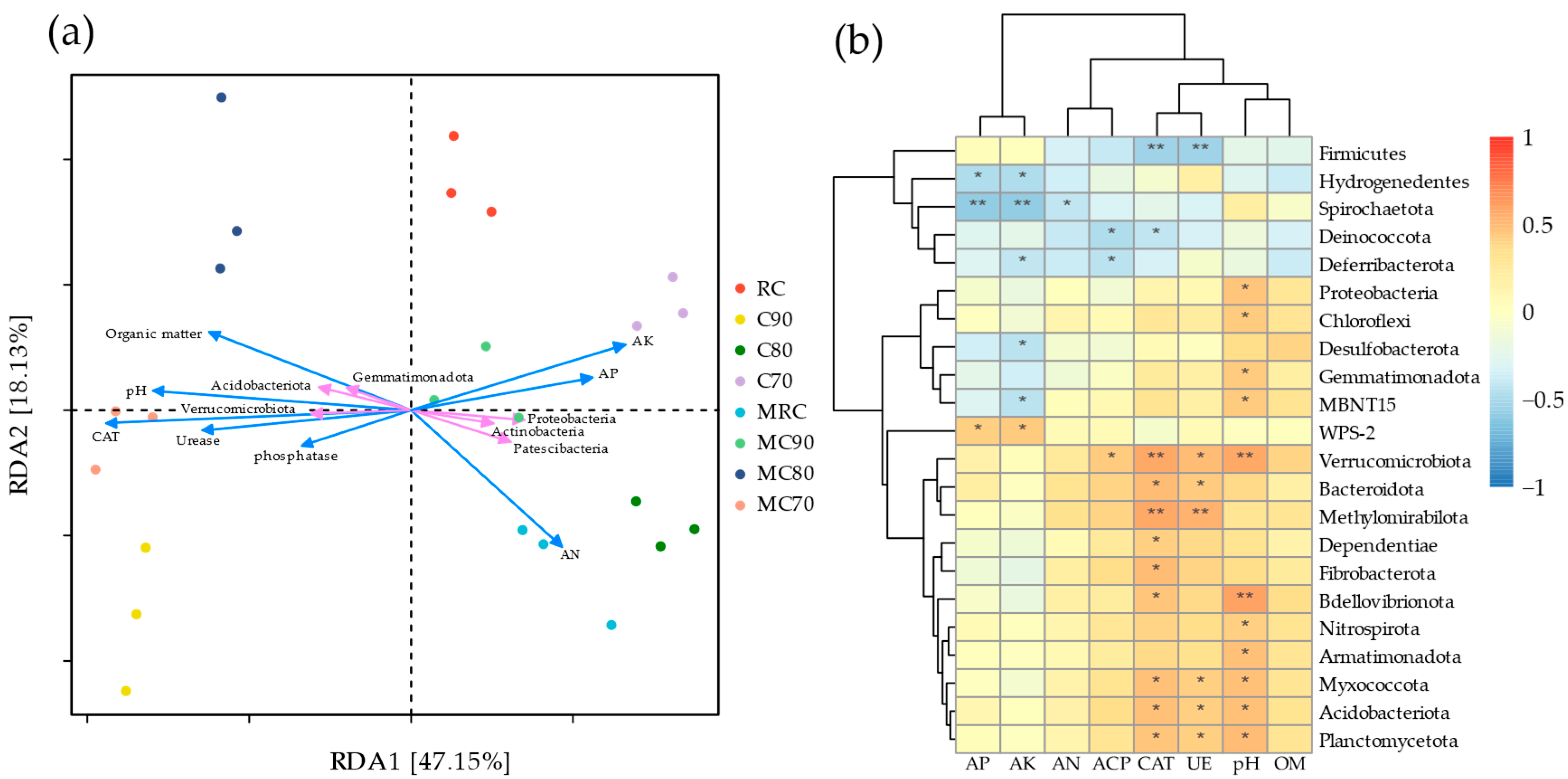
3.3. Relationship Between Rhizosphere Microbial Communities and Soil Environmental Factors
3.4. Economic Benefit and Comprehensive Evaluation
4. Discussion
4.1. Microbial Inoculants Can Partially Replace Chemical Fertilizers and Improve Soil Fertility
4.2. The Combined Application of Chemical Fertilizers and Microbial Inoculants Can Increase Crop Yields and Enhance Economic Benefits
4.3. Fertilizer Application Rates Can Affect the Diversity and Richness of Microorganisms
4.4. Interplay Between Rhizosphere Microenvironment Characteristics and Microbial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Teng, Y.; Pang, B.; Guo, X. Study on the Quality Improvement on Black Land in Northeast China under the Environment of Sustainable Agricultural Development. Kybernetes 2023, 52, 809–827. [Google Scholar] [CrossRef]
- Gu, Z.; Xie, Y.; Gao, Y.; Ren, X.; Cheng, C.; Wang, S. Quantitative Assessment of Soil Productivity and Predicted Impacts of Water Erosion in the Black Soil Region of Northeastern China. Sci. Total Environ. 2018, 637–638, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Dong, K.; Geisen, S.; Yang, W.; Yan, Y.; Gu, D.; Liu, N.; Borisjuk, N.; Luo, Y.; Friman, V.-P. The Effect of Microbial Inoculant Origin on the Rhizosphere Bacterial Community Composition and Plant Growth-Promotion. Plant Soil 2020, 452, 105–117. [Google Scholar] [CrossRef]
- Shen, M.; Li, J.; Dong, Y.; Zhang, Z.; Zhao, Y.; Li, Q.; Dang, K.; Peng, J.; Liu, H. The Effects of Microbial Inoculants on Bacterial Communities of the Rhizosphere Soil of Maize. Agriculture 2021, 11, 389. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Omidvari, M.; Abbaszadeh-Dahaji, P.; Omidvar, R.; Kariman, K. Mechanisms Underlying the Protective Effects of Beneficial Fungi against Plant Diseases. Biol. Control 2018, 117, 147–157. [Google Scholar] [CrossRef]
- Khoshnevisan, B.; Rafiee, S.; Pan, J.; Zhang, Y.; Liu, H. A Multi-Criteria Evolutionary-Based Algorithm as a Regional Scale Decision Support System to Optimize Nitrogen Consumption Rate; a Case Study in North China Plain. J. Clean. Prod. 2020, 256, 120213. [Google Scholar] [CrossRef]
- Xun, W.; Zhao, J.; Xue, C.; Zhang, G.; Ran, W.; Wang, B.; Shen, Q.; Zhang, R. Significant Alteration of Soil Bacterial Communities and Organic Carbon Decomposition by Different Long-Term Fertilization Management Conditions of Extremely Low-Productivity Arable Soil in South China. Environ. Microbiol. 2016, 18, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jiang, L.; Yang, S.; Wang, Z.; Tian, R.; Peng, Z.; Chen, Y.-L.; Zhang, X.; Kuang, J.; Ling, N.; et al. Critical Transition of Soil Bacterial Diversity and Composition Triggered by Nitrogen Enrichment. Ecology 2020, 101, e03053. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Kloepper, J.W. Plant–Microbes Interactions in Enhanced Fertilizer-Use Efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef]
- Cong, P.; Ouyang, Z.; Hou, R.; Han, D. Effects of Application of Microbial Fertilizer on Aggregation and Aggregate-Associated Carbon in Saline Soils. Soil Tillage Res. 2017, 168, 33–41. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.; Li, J.; Zou, Z.; Cao, K. Bio-Organic Fertilizer with Reduced Rates of Chemical Fertilization Improves Soil Fertility and Enhances Tomato Yield and Quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Win, K.T.; Okazaki, K.; Ohkama-Ohtsu, N.; Yokoyama, T.; Ohwaki, Y. Short-Term Effects of Biochar and Bacillus Pumilus TUAT-1 on the Growth of Forage Rice and Its Associated Soil Microbial Community and Soil Properties. Biol. Fertil. Soils 2020, 56, 481–497. [Google Scholar] [CrossRef]
- Na, X.; Yu, H.; Wang, P.; Zhu, W.; Niu, Y.; Huang, J. Vegetation Biomass and Soil Moisture Coregulate Bacterial Community Succession under Altered Precipitation Regimes in a Desert Steppe in Northwestern China. Soil Biol. Biochem. 2019, 136, 107520. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Kent, A.D.; Brisson, V.L.; Gaudin, A.C.M. Agricultural Management and Plant Selection Interactively Affect Rhizosphere Microbial Community Structure and Nitrogen Cycling. Microbiome 2019, 7, 146. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Caruso, T. Soil Microbial Community Responses to Climate Extremes: Resistance, Resilience and Transitions to Alternative States. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190112. [Google Scholar] [CrossRef] [PubMed]
- Coban, O.; De Deyn, G.B.; Van Der Ploeg, M. Soil Microbiota as Game-Changers in Restoration of Degraded Lands. Science 2022, 375, abe0725. [Google Scholar] [CrossRef]
- Domene, X.; Hanley, K.; Enders, A.; Lehmann, J. Short-Term Mesofauna Responses to Soil Additions of Corn Stover Biochar and the Role of Microbial Biomass. Appl. Soil Ecol. 2015, 89, 10–17. [Google Scholar] [CrossRef]
- Sun, L.; Gao, J.; Huang, T.; Kendall, J.R.A.; Shen, Q.; Zhang, R. Parental Material and Cultivation Determine Soil Bacterial Community Structure and Fertility. FEMS Microbiol. Ecol. 2015, 91, 1–10. [Google Scholar] [CrossRef]
- Ye, L.; Wu, X.; Wu, C.; Zhang, Y.; Meng, L.; Bao, E.; Cao, K. Response of Soil Bacterial Community to Agricultural Reclamation in the Tengger Desert, Northwestern China. Appl. Soil Ecol. 2022, 169, 104189. [Google Scholar] [CrossRef]
- Tian, J.; He, N.; Hale, L.; Niu, S.; Yu, G.; Liu, Y.; Blagodatskaya, E.; Kuzyakov, Y.; Gao, Q.; Zhou, J. Soil Organic Matter Availability and Climate Drive Latitudinal Patterns in Bacterial Diversity from Tropical to Cold Temperate Forests. Funct. Ecol. 2017, 32, 61–70. [Google Scholar] [CrossRef]
- Fierer, N.; Wood, S.A.; Bueno De Mesquita, C.P. How Microbes Can, and Cannot, Be Used to Assess Soil Health. Soil Biol. Biochem. 2021, 153, 108111. [Google Scholar] [CrossRef]
- Yang, L.; Lou, J.; Wang, H.; Wu, L.; Xu, J. Use of an improved high-throughput absolute abundance quantification method to characterize soil bacterial community and dynamics. Sci. Total Environ. 2018, 633, 360–371. [Google Scholar] [CrossRef]
- Jiang, S.-Q.; Yu, Y.-N.; Gao, R.-W.; Wang, H.; Zhang, J.; Li, R.; Long, X.-H.; Shen, Q.-R.; Chen, W.; Cai, F. High-Throughput Absolute Quantification Sequencing Reveals the Effect of Different Fertilizer Applications on Bacterial Community in a Tomato Cultivated Coastal Saline Soil. Sci. Total Environ. 2019, 687, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Jin, X.-Y.; Zhang, X.-C.; Chen, L.; Liu, J.-L.; Zhang, H.-M.; Zhang, X.; Zhang, Y.-F.; Zhao, J.-H.; Ma, Z.-S.; et al. Comparative Metagenomics of Two Distinct Biological Soil Crusts in the Tengger Desert, China. Soil Biol. Biochem. 2020, 140, 107637. [Google Scholar] [CrossRef]
- Smets, W.; Leff, J.W.; Bradford, M.A.; McCulley, R.L.; Lebeer, S.; Fierer, N. A Method for Simultaneous Measurement of Soil Bacterial Abundances and Community Composition via 16S rRNA Gene Sequencing. Soil Biol. Biochem. 2016, 96, 145–151. [Google Scholar] [CrossRef]
- Pysarenko, P.; Samojlik, M.; Galytskaya, M.; Tsova, Y.; Mostoviak, I. Influence of Bacillus Subtilis on Soil Microbiocenosis. Ecol. Quest. 2023, 34, 127–133. [Google Scholar] [CrossRef]
- Malvestiti, G.S. Resposta Técnica e Econômica para Adubação Com N, P e K em Milho Convencional e Geneticamente Modificado; Mestrado em Qualidade e Produtividade Animal; Universidade de São Paulo: São Paulo, Brazil, 2014. [Google Scholar]
- Paolinelli, A.; Dourado Neto, D.; Mantovani, E.C. (Eds.) Agricultura Irrigada no Brasil: Ciência e Tecnología; Universidade de São Paulo. Escola Superior de Agricultura Luiz de Queiroz; Universidade Federal de Viçosa; Universidade de São Paulo: São Paulo, Brazil, 2022; ISBN 978-65-87391-23-6. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis; China Agriculture Press: Beijing, China, 2000; ISBN 7-109-06644-4. [Google Scholar]
- Ma, Z.; Xu, W.; Chen, Y.; Liu, M.; Wen, J. A Study of the Influence of the Type of Land Use on the Enzymatic Activity of Soils in Southwestern China. Forests 2024, 15, 581. [Google Scholar] [CrossRef]
- Sun, J.; Ma, J.; Zhu, M.; Yuan, Z.; Yao, J. Effects of Nitrogen Application on the Soil Microbial Activity, Enzyme Activities and Properties and Their Relationships in a Maize Field. Fresenius Environ. Bull. 2016, 25, 852–861. [Google Scholar]
- Mekonnen, T.W. Effects of Planting Date, Environments and Their Interaction on Grain Yield and Quality Traits of Maize Hybrids. Heliyon 2023, 9, e21660. [Google Scholar] [CrossRef]
- Kataoka, T.; Ooki, A.; Nomura, D. Production of Dibromomethane and Changes in the Bacterial Community in Bromoform-Enriched Seawater. Microbes Environ. 2019, 34, 215–218. [Google Scholar] [CrossRef]
- Su, X.; Zhang, L.; Meng, H.; Wang, H.; Zhao, J.; Sun, X.; Song, X.; Zhang, X.; Mao, L. Long-Term Conservation Tillage Increase Cotton Rhizosphere Sequestration of Soil Organic Carbon by Changing Specific Microbial CO2 Fixation Pathways in Coastal Saline Soil. J. Environ. Manag. 2024, 358, 120743. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, D.; Ren, Y.; Wang, Z.; Zhou, Y. Soil Quality Assessment of Croplands in the Black Soil Zone of Jilin Province, China: Establishing a Minimum Data Set Model. Ecol. Indic. 2019, 107, 105251. [Google Scholar] [CrossRef]
- Masto, R.E.; Chhonkar, P.K.; Singh, D.; Patra, A.K. Soil Quality Response to Long-Term Nutrient and Crop Management on a Semi-Arid Inceptisol. Agric. Ecosyst. Environ. 2007, 118, 130–142. [Google Scholar] [CrossRef]
- Masto, R.E.; Chhonkar, P.K.; Singh, D.; Patra, A.K. Alternative Soil Quality Indices for Evaluating the Effect of Intensive Cropping, Fertilisation and Manuring for 31 Years in the Semi-Arid Soils of India. Environ. Monit. Assess. 2007, 136, 419–435. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Zheng, M.; Jiang, L.; Luo, Y. Patterns and Mechanisms of Responses by Soil Microbial Communities to Nitrogen Addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, Y.; Yang, Y.; Zhang, M.; Mao, X.; Guo, Y.; Li, X.; Tao, B.; Qi, Y.; Ma, L.; et al. Co-Application of Biochar and Microbial Inoculants Increases Soil Phosphorus and Potassium Fertility and Improves Soil Health and Tomato Growth. J. Soils Sediments 2023, 23, 947–957. [Google Scholar] [CrossRef]
- Rose, M.T.; Phuong, T.L.; Nhan, D.K.; Cong, P.T.; Hien, N.T.; Kennedy, I.R. Up to 52% N Fertilizer Replaced by Biofertilizer in Lowland Rice via Farmer Participatory Research. Agron. Sustain. Dev. 2014, 34, 857–868. [Google Scholar] [CrossRef]
- Da Costa, P.B.; Beneduzi, A.; De Souza, R.; Schoenfeld, R.; Vargas, L.K.; Passaglia, L.M.P. The Effects of Different Fertilization Conditions on Bacterial Plant Growth Promoting Traits: Guidelines for Directed Bacterial Prospection and Testing. Plant Soil 2013, 368, 267–280. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant Growth-Promoting Rhizobacteria Allow Reduced Application Rates of Chemical Fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef]
- Zhang, C.; Song, Z.; Zhuang, D.; Wang, J.; Xie, S.; Liu, G. Urea Fertilization Decreases Soil Bacterial Diversity, but Improves Microbial Biomass, Respiration, and N-Cycling Potential in a Semiarid Grassland. Biol. Fertil. Soils 2019, 55, 229–242. [Google Scholar] [CrossRef]
- Xia, Z.; Yang, J.; Sang, C.; Wang, X.; Sun, L.; Jiang, P.; Wang, C.; Bai, E. Phosphorus Reduces Negative Effects of Nitrogen Addition on Soil Microbial Communities and Functions. Microorganisms 2020, 8, 1828. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Fang, F.; Wu, L.; Gao, F.; Li, M.; Li, B.; Wu, K.; Hu, X.; Wang, S.; Wei, Z.; et al. The Microbial Community, Nutrient Supply and Crop Yields Differ along a Potassium Fertilizer Gradient under Wheat–Maize Double-Cropping Systems. J. Integr. Agric. 2024, 23, 3592–3609. [Google Scholar] [CrossRef]
- Liu, L.; Gao, Z.; Liu, W.; Li, H.; Wang, Z.; Liu, J. Phosphorus Fertilizer Input Threshold Shifts Bacterial Community Structure and Soil Multifunctionality to Maintain Dryland Wheat Production. Soil Tillage Res. 2024, 243, 106174. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Huber, K.J.; Pester, M.; Eichorst, S.A.; Navarrete, A.A.; Foesel, B.U. Editorial: Acidobacteria—Towards Unraveling the Secrets of a Widespread, Though Enigmatic, Phylum. Front. Microbiol. 2022, 13, 960602. [Google Scholar] [CrossRef]
- Trojan, D.; Garcia-Robledo, E.; Meier, D.V.; Hausmann, B.; Revsbech, N.P.; Eichorst, S.A.; Woebken, D. Microaerobic Lifestyle at Nanomolar O2 Concentrations Mediated by Low-Affinity Terminal Oxidases in Abundant Soil Bacteria. mSystems 2021, 6, e0025021. [Google Scholar] [CrossRef]
- Falagán, C.; Foesel, B.; Johnson, B. Acidicapsa ferrireducens Sp. Nov., Acidicapsa acidiphila Sp. Nov., and Granulicella acidiphila Sp. Nov.: Novel Acidobacteria Isolated from Metal-Rich Acidic Waters. Extremophiles 2017, 21, 459–469. [Google Scholar] [CrossRef]
- Flieder, M.; Buongiorno, J.; Herbold, C.W.; Hausmann, B.; Rattei, T.; Lloyd, K.G.; Loy, A.; Wasmund, K. Novel Taxa of Acidobacteriota Implicated in Seafloor Sulfur Cycling. ISME J. 2021, 15, 3159–3180. [Google Scholar] [CrossRef]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three Genomes from the Phylum Acidobacteria Provide Insight into the Lifestyles of These Microorganisms in Soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef]
- Shelyakin, P.V.; Semenkov, I.N.; Tutukina, M.N.; Nikolaeva, D.D.; Sharapova, A.V.; Sarana, Y.V.; Lednev, S.A.; Smolenkov, A.D.; Gelfand, M.S.; Krechetov, P.P.; et al. The influence of kerosene on microbiomes of diverse soils. Life 2022, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The Interplay between Microbial Communities and Soil Properties. Nat. Rev. Microbiol. 2024, 22, 226–239. [Google Scholar] [CrossRef]
- Wang, S.; Heal, K.V.; Zhang, Q.; Yu, Y.; Tigabu, M.; Huang, S.; Zhou, C. Soil Microbial Community, Dissolved Organic Matter and Nutrient Cycling Interactions Change along an Elevation Gradient in Subtropical China. J. Environ. Manag. 2023, 345, 118793. [Google Scholar] [CrossRef]
- Wang, X.; Bian, Q.; Jiang, Y.; Zhu, L.; Chen, Y.; Liang, Y.; Sun, B. Organic Amendments Drive Shifts in Microbial Community Structure and Keystone Taxa Which Increase C Mineralization across Aggregate Size Classes. Soil Biol. Biochem. 2021, 153, 108062. [Google Scholar] [CrossRef]
- Huet, S.; Romdhane, S.; Breuil, M.-C.; Bru, D.; Mounier, A.; Spor, A.; Philippot, L. Experimental Community Coalescence Sheds Light on Microbial Interactions in Soil and Restores Impaired Functions. Microbiome 2023, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.; Buddolla, V.; Lian, B. Oxalate Carbonate Pathway—Conversion and Fixation of Soil Carbon—A Potential Scenario for Sustainability. Front. Plant Sci. 2020, 11, 591297. [Google Scholar] [CrossRef]
- Bargaz, A.; Elhaissoufi, W.; Khourchi, S.; Benmrid, B.; Borden, K.A.; Rchiad, Z. Benefits of Phosphate Solubilizing Bacteria on Belowground Crop Performance for Improved Crop Acquisition of Phosphorus. Microbiol. Res. 2021, 252, 126842. [Google Scholar] [CrossRef]
- Ikoyi, I.; Fowler, A.; Schmalenberger, A. One-Time Phosphate Fertilizer Application to Grassland Columns Modifies the Soil Microbiota and Limits Its Role in Ecosystem Services. Sci. Total Environ. 2018, 630, 849–858. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Inoculant, Fertilizers, and Other Inputs (Yuan hm−2) | Yield (kg hm−2) | Gross Product (Yuan hm−2) | Net Income (Yuan hm−2) | Input–Output Ratio |
|---|---|---|---|---|---|
| RC | 7600 | 10,710.03 b | 13,923.04 | 6323.04 | 1:1.41 |
| C90 | 7250 | 10,473.49 bc | 13,615.53 | 6365.53 | 1:1.44 |
| C80 | 6900 | 10,348.03 bc | 13,452.44 | 6552.44 | 1:1.50 |
| C70 | 6550 | 10,088.83 c | 13,115.48 | 6565.48 | 1:1.54 |
| MRC | 7860 | 11,769.77 a | 15,300.70 | 7440.70 | 1:1.50 |
| MC90 | 7510 | 11,431.49 a | 14,860.93 | 7350.93 | 1:1.52 |
| MC80 | 7160 | 11,326.42 a | 14,724.35 | 7564.35 | 1:1.58 |
| MC70 | 6810 | 10,707.05 b | 13,919.16 | 7109.16 | 1:1.57 |
| Treatment | PC1 Score | Ranking | PC2 Score | Ranking | PC3 Score | Ranking | Comprehensive Score | Integrated Ranking |
|---|---|---|---|---|---|---|---|---|
| RC | −0.228 | 5 | −0.292 | 4 | −1.381 | 8 | −0.551 | 7 |
| C90 | −1.386 | 6 | 1.111 | 2 | −0.379 | 6 | −0.412 | 5 |
| C80 | −0.303 | 7 | −0.806 | 7 | −0.602 | 7 | −0.525 | 6 |
| C70 | −0.535 | 8 | −1.262 | 8 | −0.102 | 4 | −0.627 | 8 |
| MRC | 1.585 | 1 | −0.505 | 6 | 0.395 | 2 | 0.677 | 2 |
| MC90 | 1.222 | 2 | 0.401 | 3 | −0.255 | 5 | 0.598 | 3 |
| MC80 | 0.361 | 3 | 1.702 | 1 | 0.253 | 3 | 0.713 | 1 |
| MC70 | −0.716 | 4 | −0.35 | 5 | 2.072 | 1 | 0.125 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Wang, N.; Zhao, C.; Yang, L.; Zhao, X.; Gao, H.; Zhang, F.; Wang, H.; Huang, N. Enhancing Black Soil Fertility and Microbial Community Structure via Microbial Agents to Reduce Chemical Fertilizer Dependency: A Strategy to Boost Maize Yield. Agronomy 2025, 15, 1029. https://doi.org/10.3390/agronomy15051029
Zhang F, Wang N, Zhao C, Yang L, Zhao X, Gao H, Zhang F, Wang H, Huang N. Enhancing Black Soil Fertility and Microbial Community Structure via Microbial Agents to Reduce Chemical Fertilizer Dependency: A Strategy to Boost Maize Yield. Agronomy. 2025; 15(5):1029. https://doi.org/10.3390/agronomy15051029
Chicago/Turabian StyleZhang, Fenglin, Nan Wang, Chenyu Zhao, Luze Yang, Xingmin Zhao, Hongjun Gao, Fugui Zhang, Hongbin Wang, and Ning Huang. 2025. "Enhancing Black Soil Fertility and Microbial Community Structure via Microbial Agents to Reduce Chemical Fertilizer Dependency: A Strategy to Boost Maize Yield" Agronomy 15, no. 5: 1029. https://doi.org/10.3390/agronomy15051029
APA StyleZhang, F., Wang, N., Zhao, C., Yang, L., Zhao, X., Gao, H., Zhang, F., Wang, H., & Huang, N. (2025). Enhancing Black Soil Fertility and Microbial Community Structure via Microbial Agents to Reduce Chemical Fertilizer Dependency: A Strategy to Boost Maize Yield. Agronomy, 15(5), 1029. https://doi.org/10.3390/agronomy15051029