
Fermentation Dynamics, Microbial Succession, and Metabolic Shifts in High-Moisture Giant Juncao Silage




Abstract
1. Introduction
2. Materials and Methods
2.1. Silage Production
2.2. Chemical Composition and Fermentation Quality Analyses
2.3. HTS and Functional Prediction Analyses
2.4. Statistical Analysis
3. Results
3.1. Composition of Fresh GJ
3.2. Fermentation Characteristics of NGJ
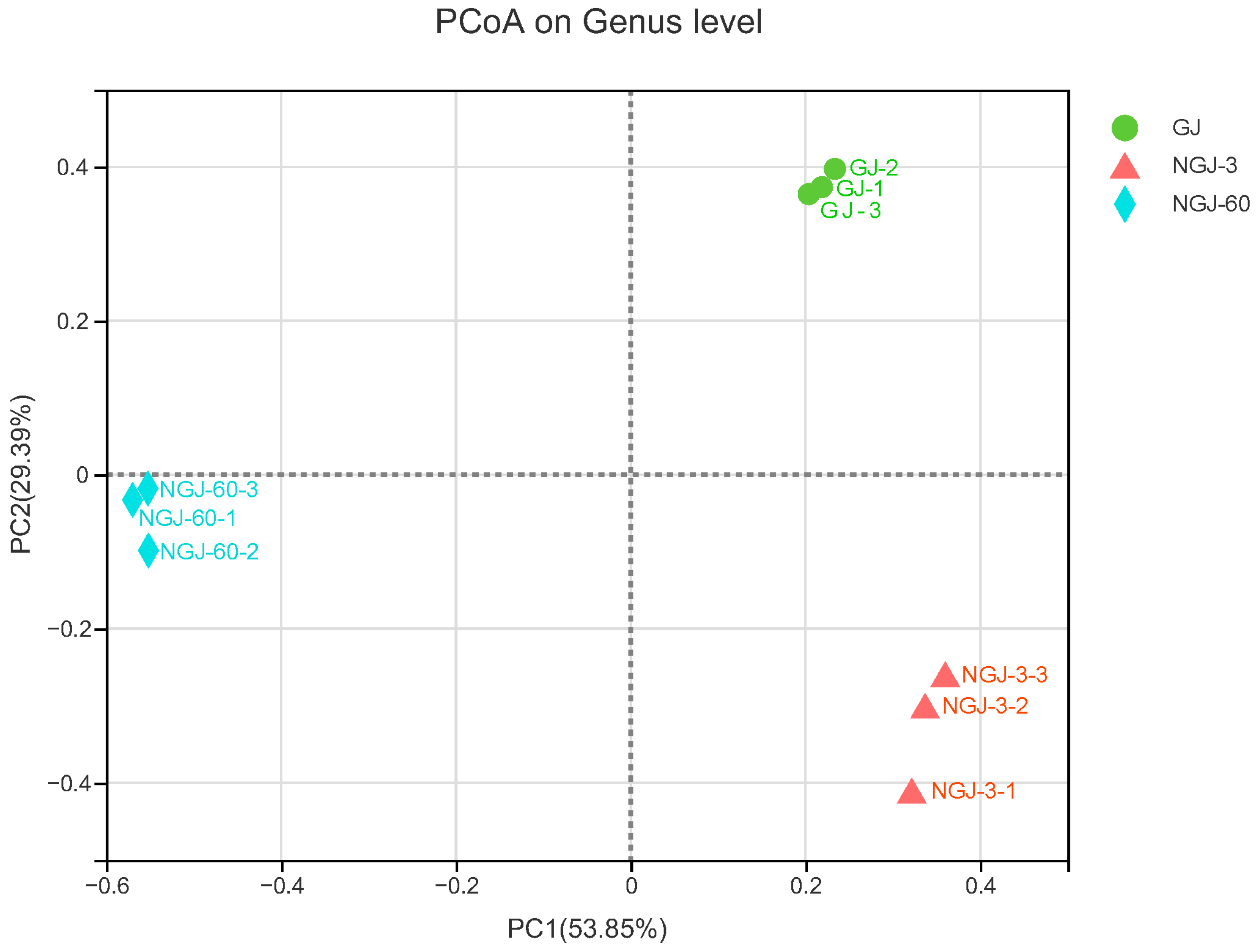
3.3. The Bacterial Community of Fresh GJ and NGJ
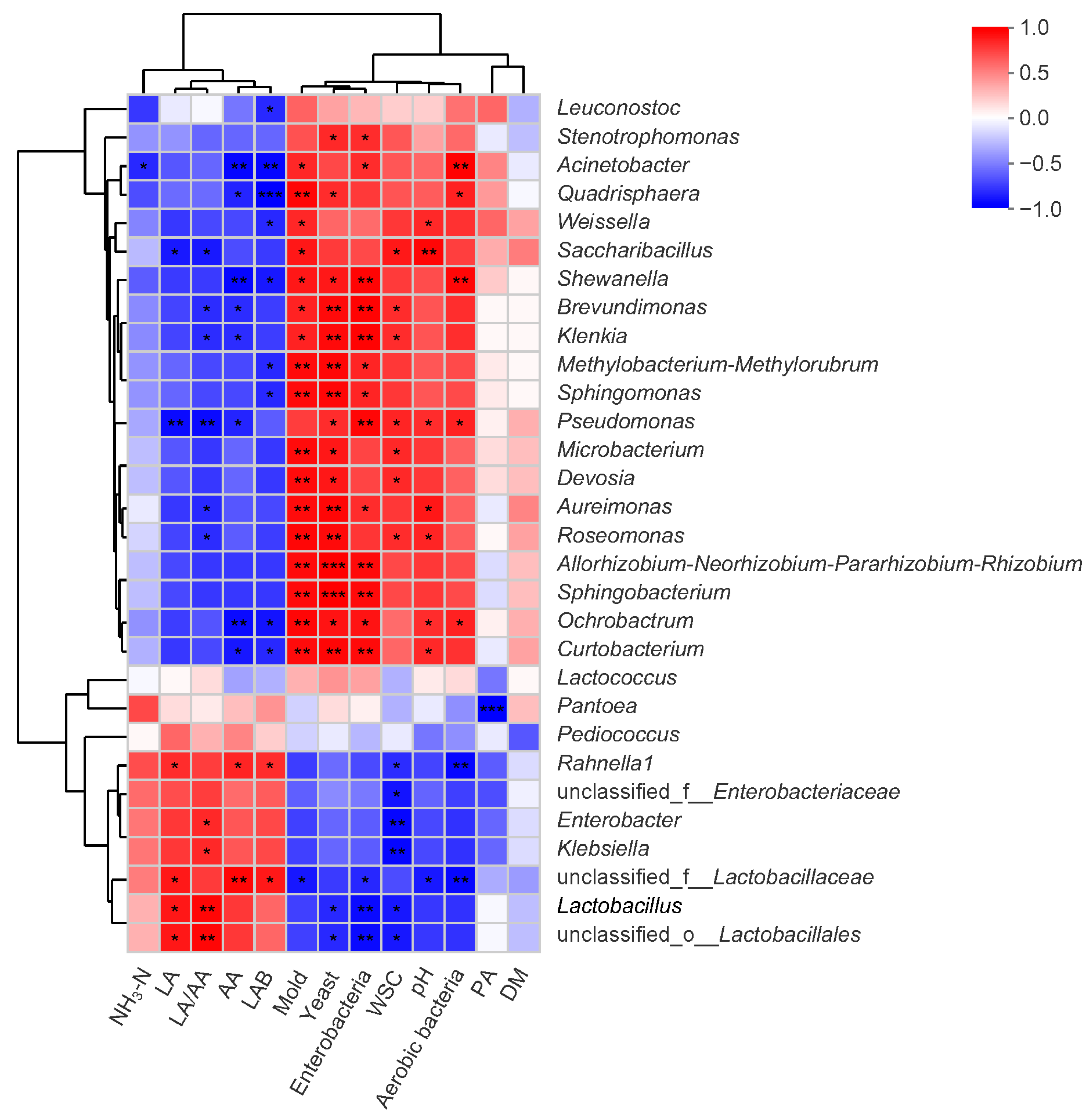
3.4. Interrelationships Between Fermentation Metabolites and Microbial Consortia
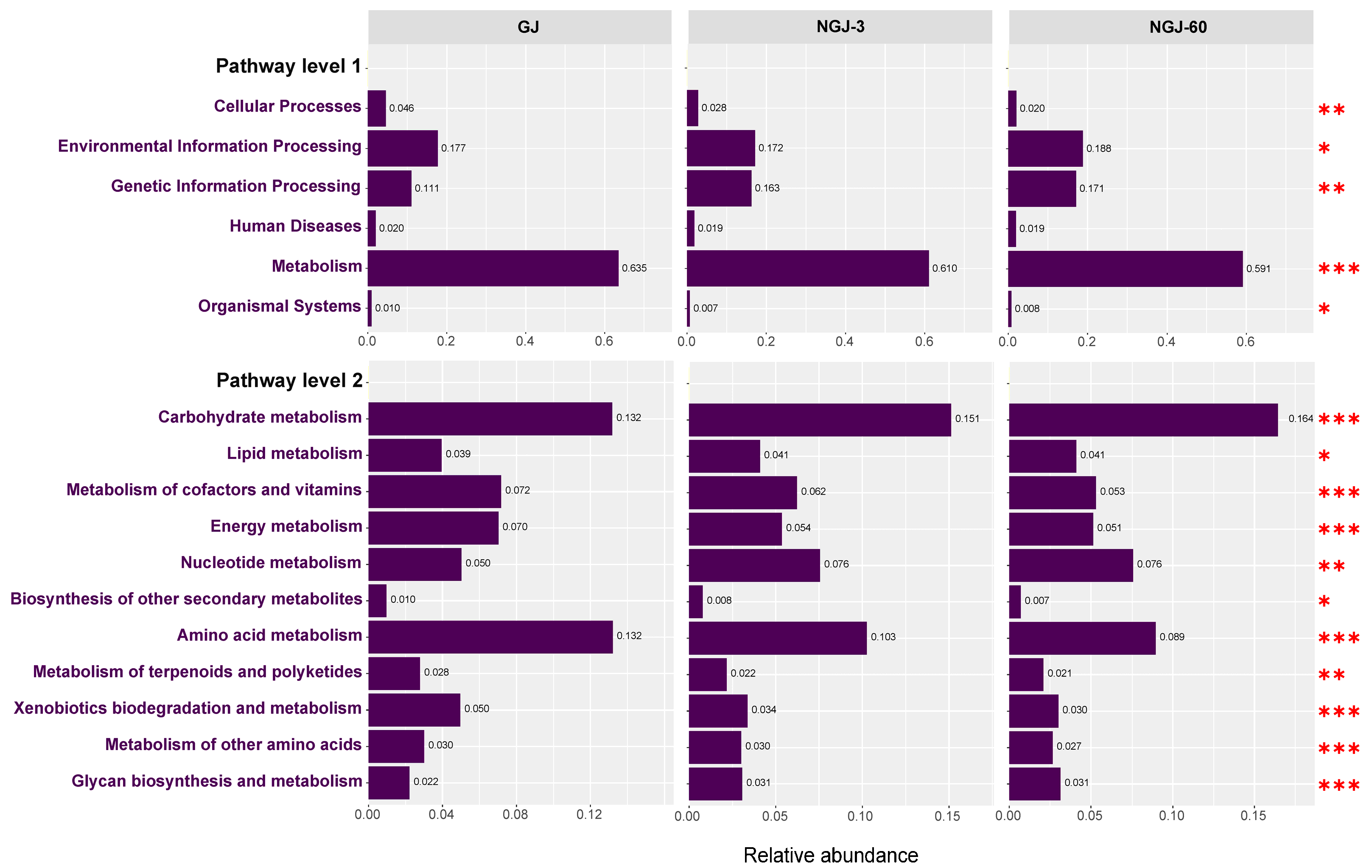
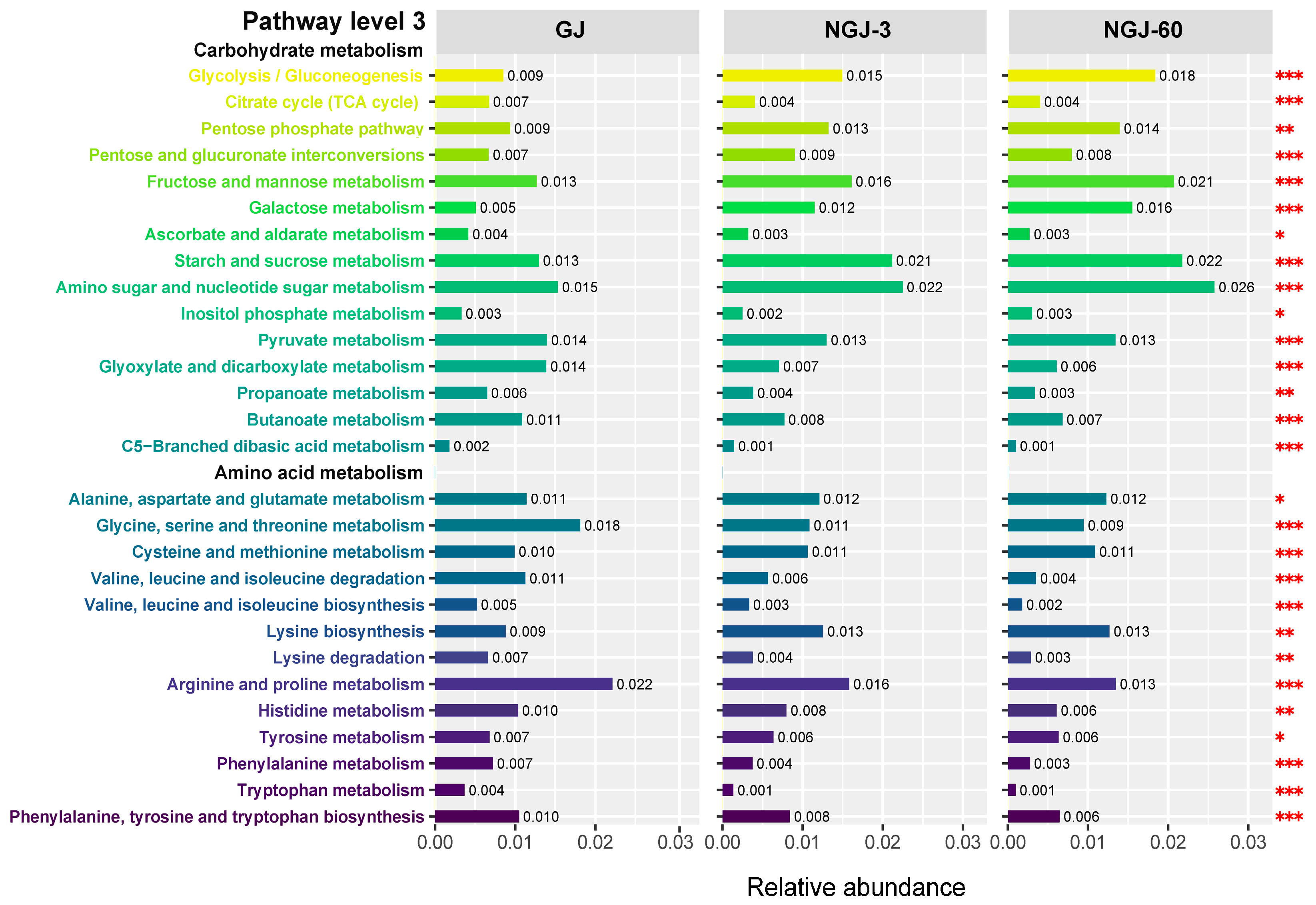
3.5. Functional Profiling of the Bacterial Community
4. Discussion
4.1. Analysis of Fresh GJ
4.2. Effects of Ensiling Days on Fermentation Quality of NGJ
4.3. Effects of Ensiling Days on the Bacterial Community of NGJ
4.4. Relationships Between Fermentation Parameters and Bacterial Communities of NGJ
4.5. Effects of Ensiling Days on the Potential Functions of the Bacterial Community in NGJ
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, F.; Wu, M.; Wang, X. Research Progress in Woody Forage. Feed Industry. 2025, 46, 1–7. (In Chinese) [Google Scholar]
- Lin, B.; Liu, J.; Lv, Z.; Luo, M.; Lin, Z. Preparation and properties of immobilized particles containing highly efficient nitrogen-fixing Klebsiella variicola GN02 cells isolated from the Pennisetum giganteum z. x. lin roots. Appl. Biochem. Microbiol. 2020, 56, 58–63. [Google Scholar] [CrossRef]
- Li, Q.; Xiang, C.; Xu, L.; Cui, J.; Fu, S.; Chen, B.; Yang, S.; Wang, P.; Xie, Y.; Wei, M.; et al. SMRT sequencing of a full-length transcriptome reveals transcript variants involved in C18 unsaturated fatty acid biosynthesis and metabolism pathways at chilling temperature in Pennisetum giganteum. BMC Genom. 2020, 21, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhang JinGuo, Z.J.; Lei HeXian, L.H.; Li JiFeng, L.J.; Zhao JianGang, Z.J.; Yang GuiFen, Y.G.; Ren MingJin, R.M.; Yang XueSong, Y.X. Growth performance of Pennisetum sp. at different elevation. Guizhou Agricultural Sciences. Sci. 2013, 41, 112–115. (In Chinese) [Google Scholar]
- Zhao, J.; Yin, X.; Dong, Z.; Wang, S.; Li, J.; Dong, D.; Shao, T. Using gamma-ray irradiation and epiphytic microbiota inoculation to separate the effects of chemical and microbial factors on fermentation quality and bacterial community of ensiled Pennisetum giganteum. J. Appl. Microbiol. 2022, 132, 1675–1686. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Bacterial dynamics of wheat silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef]
- Okoye, C.O.; Wang, Y.; Gao, L.; Wu, Y.; Li, X.; Sun, J.; Jiang, J. The performance of lactic acid bacteria in silage production: A review of modern biotechnology for silage improvement. Microbiol. Res. 2023, 266, 127212. [Google Scholar] [CrossRef]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, Z.; Li, J.; Chen, L.; Bai, Y.; Jia, Y.; Shao, T. Ensiling as pretreatment of rice straw: The effect of hemicellulase and Lactobacillus plantarum on hemicellulose degradation and cellulose conversion. Bioresour. Technol. 2018, 266, 158–165. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Horiguchi, K.; Goto, M. Effect of crushing unhulled rice and the addition of fermented juice of epiphytic lactic acid bacteria on the fermentation quality of whole crop rice silage, and its digestibility and rumen fermentation status in sheep. Anim. Sci. J. 2005, 76, 353–358. [Google Scholar] [CrossRef]
- Jing, Y.; Mu, C.; Wang, H.; Shen, J.; Zoetendal, E.G.; Zhu, W. Amino acid utilization allows intestinal dominance of Lactobacillus amylovorus. ISME J. 2022, 16, 2491–2502. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of applying lactic acid bacteria isolated from forage crops on fermentation characteristics and aerobic deterioration of silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef]
- Kung, L.; Shaver, R. Interpretation and use of silage fermentation analysis reports. Focus Forage 2001, 3, 1–5. [Google Scholar]
- van Niekerk, W.A.; Hassen, A.; Bechaz, F.M. Influence of growth stage at harvest on fermentative characteristics of Panicum maximum silage. S. Afr. J. Anim. Sci. 2010, 40, 334–341. [Google Scholar] [CrossRef]
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Pian, R.; Chen, X.; Zhang, Q. Ensiling characteristics, proteolysis and bacterial community of high-moisture corn stalk and stylo silage prepared with Bauhinia variegate flower. Bioresour. Technol. 2020, 296, 122336. [Google Scholar] [CrossRef]
- Zhao, J.; Yin, X.J.; Wang, S.R.; Li, J.F.; Shao, T. Separating the effects of chemical and microbial factors on fermentation quality and bacterial community of Napier grass silage by using gamma-ray irradiation and epiphytic microbiota transplantation. Anim. Feed Sci. Technol. 2021, 280, 115082. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, J.; Liu, J.; Yang, F.; Zhu, W.; Yuan, X.; Hu, Y.; Cui, Z.; Wang, X. Effect of ensiling and silage additives on biogas production and microbial community dynamics during anaerobic digestion of switchgrass. Bioresour. Technol. 2017, 241, 349–359. [Google Scholar] [CrossRef]
- Pang, H.; Zhang, M.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Identification of lactic acid bacteria isolated from corn stovers. Anim. Sci. J. 2011, 82, 642–653. [Google Scholar] [CrossRef]
- Muck, R.E. Recent advances in silage microbiology. Agric. Food Sci. 2013, 22, 3–15. [Google Scholar] [CrossRef]
- Graf, K.; Ulrich, A.; Idler, C.; Klocke, M. Bacterial community dynamics during ensiling of perennial ryegrass at two compaction levels monitored by terminal restriction fragment length polymorphism. J. Appl. Microbiol. 2016, 120, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef]
- McGarvey, J.A.; Franco, R.B.; Palumbo, J.D.; Hnasko, R.; Stanker, L.; Mitloehner, F.M. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol. 2013, 114, 1661–1670. [Google Scholar] [CrossRef]
- Bai, J.; Ding, Z.; Ke, W.; Xu, D.; Wang, M.; Huang, W.; Zhang, Y.; Liu, F.; Guo, X. Different lactic acid bacteria and their combinations regulated the fermentation process of ensiled alfalfa: Ensiling characteristics, dynamics of bacterial community and their functional shifts. Microb. Biotechnol. 2021, 14, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Flythe, M.D.; Russell, J.B. The effect of pH and a bacteriocin (bovicin HC5) on Clostridium sporogenes MD1, a bacterium that has the ability to degrade amino acids in ensiled plant materials. FEMS Microbiol. Ecol. 2004, 47, 215–222. [Google Scholar] [CrossRef]
- Liu, Q.H.; Wu, J.X.; Shao, T. Roles of microbes and lipolytic enzymes in changing the fatty acid profile, α-tocopherol and β-carotene of whole-crop oat silages during ensiling and after exposure to air. Anim. Feed Sci. Technol. 2019, 253, 81–92. [Google Scholar] [CrossRef]
- Xu, D.; Wang, N.; Rinne, M.; Ke, W.; Weinberg, Z.G.; Da, M.; Bai, J.; Zhang, Y.; Li, F.; Guo, X. The bacterial community and metabolome dynamics and their interactions modulate fermentation process of whole crop corn silage prepared with or without inoculants. Microb. Biotechnol. 2021, 14, 561–576. [Google Scholar] [CrossRef]
- Yao, Z.; Guo, Y.; Wang, H.; Chen, Y.; Wang, Q.; Nielsen, J.; Dai, Z. Integrated microbiology and metabolomic analysis reveal the improvement of rice straw silage quality by inoculation of Lactobacillus brevis. Biotechnol. Biofuels Bioprod. 2023, 16, 25. [Google Scholar] [CrossRef]
- Kilstrup, M.; Hammer, K.; Ruhdal Jensen, P.; Martinussen, J. Nucleotide metabolism and its control in lactic acid bacteria. FEMS Microbiol. Rev. 2005, 29, 555–590. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Lactic acid production from lignocellulose-derived sugars using lactic acid bacteria: Overview and limits. J. Biotechnol. 2011, 156, 286–301. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | GJ |
|---|---|
| pH | 5.68 ± 0.09 |
| DM (% FM) | 16.6 ± 0.31 |
| WSC (% DM) | 8.57 ± 0.12 |
| CP (% DM) | 14.6 ± 0.44 |
| BC (mEq/kg DM) | 39.6 ± 2.03 |
| NDF (% DM) | 53.5 ± 1.23 |
| ADF (% DM) | 29.6 ± 1.44 |
| LAB (Log10 CFU/g FM) | 4.60 ± 0.17 |
| Aerobic bacteria (Log10 CFU/g FM) | 5.85 ± 0.16 |
| Yeast (Log10 CFU/g FM) | 4.76 ± 0.12 |
| Molds (Log10 CFU/g FM) | 4.60 ± 0.02 |
| Enterobacteria (Log10 CFU/g FM) | 5.70 ± 0.15 |
| Items 1 | Ensiling Days 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 15 | 30 | 60 | |||
| pH | 5.57 a | 5.64 a | 5.62 a | 4.76 b | 4.03 c | 4.16 c | 0.165 | <0.001 |
| DM (% FM) | 16.0 a | 15.2 a | 14.8 ab | 14.6 ab | 14.3 ab | 13.1 b | 0.248 | 0.006 |
| LA (% DM) | ND b | 0.04 b | 0.09 b | 1.57 ab | 4.23 a | 3.00 ab | 0.475 | 0.014 |
| AA (% DM) | 0.25 d | 0.54 d | 0.36 d | 1.26 c | 2.12 b | 2.97 a | 0.238 | <0.001 |
| LA/AA | — b | 0.07 b | 0.23 b | 1.25 ab | 2.01 a | 1.01 ab | 0.214 | 0.012 |
| PA (% DM) | 0.04 b | 0.18 a | 0.15 a | 0.09 ab | 0.14 a | 0.02 b | 0.018 | 0.046 |
| BA (% DM) | ND | ND | ND | ND | ND | 0.01 | — | — |
| VFA (% DM) | 0.29 d | 0.71 cd | 0.51 bc | 1.35 ab | 2.2 ab | 3.00 a | 0.234 | <0.001 |
| Ethanol (% DM) | ND | ND | ND | 0.24 | 0.10 | 0.50 | — | — |
| 1,2-Propanediol (% DM) | ND | ND | ND | ND | ND | 0.15 | — | — |
| WSC (% DM) | 8.21 a | 8.05 ab | 7.71 ab | 6.74 b | 5.02 c | 4.16 c | 0.377 | <0.001 |
| NH3-N (% TN) | 7.35 b | 8.28 b | 9.20 b | 11.9 ab | 12.5 ab | 16.7 a | 0.836 | 0.001 |
| V-score | 95 | 89 | 89 | 74 | 70 | 52 | — | — |
| Items | Ensiling Days 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 15 | 30 | 60 | |||
| LAB (log10 CFU/g FM) | 3.80 d | 4.53 cd | 4.68 c | 6.37 b | 7.94 a | 7.07 b | 0.357 | <0.001 |
| Aerobic bacteria (log10 CFU/g FM) | 5.48 a | 4.34 ab | 3.50 ab | 2.63 ab | <2.00 b | 2.60 ab | 0.364 | 0.037 |
| Yeast (log10 CFU/g FM) | 4.34 a | 4.27 a | 4.30 a | 4.04 ab | 3.47 b | 3.52 b | 0.112 | 0.039 |
| Molds (log10 CFU/g FM) | 4.54 a | 4.50 ab | 3.88 b | 3.04 c | ND d | ND d | 0.461 | <0.001 |
| Enterobacteria (log10 CFU/g FM) | 5.71 a | 5.56 ab | 5.29 ab | 3.81 bc | <2.00 d | 2.60 cd | 0.436 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.-Y.; Shao, T.; Li, J.-F.; Dong, Z.-H.; Zhao, J. Fermentation Dynamics, Microbial Succession, and Metabolic Shifts in High-Moisture Giant Juncao Silage. Agronomy 2025, 15, 1028. https://doi.org/10.3390/agronomy15051028
Liang X-Y, Shao T, Li J-F, Dong Z-H, Zhao J. Fermentation Dynamics, Microbial Succession, and Metabolic Shifts in High-Moisture Giant Juncao Silage. Agronomy. 2025; 15(5):1028. https://doi.org/10.3390/agronomy15051028
Chicago/Turabian StyleLiang, Xin-Yu, Tao Shao, Jun-Feng Li, Zhi-Hao Dong, and Jie Zhao. 2025. "Fermentation Dynamics, Microbial Succession, and Metabolic Shifts in High-Moisture Giant Juncao Silage" Agronomy 15, no. 5: 1028. https://doi.org/10.3390/agronomy15051028
APA StyleLiang, X.-Y., Shao, T., Li, J.-F., Dong, Z.-H., & Zhao, J. (2025). Fermentation Dynamics, Microbial Succession, and Metabolic Shifts in High-Moisture Giant Juncao Silage. Agronomy, 15(5), 1028. https://doi.org/10.3390/agronomy15051028