Nitrogen Dynamics in Sunn Hemp Intercropped Tall Fescue Pastures

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design and Management
2.3. Sampling and Evaluations
2.4. Statistical Analysis
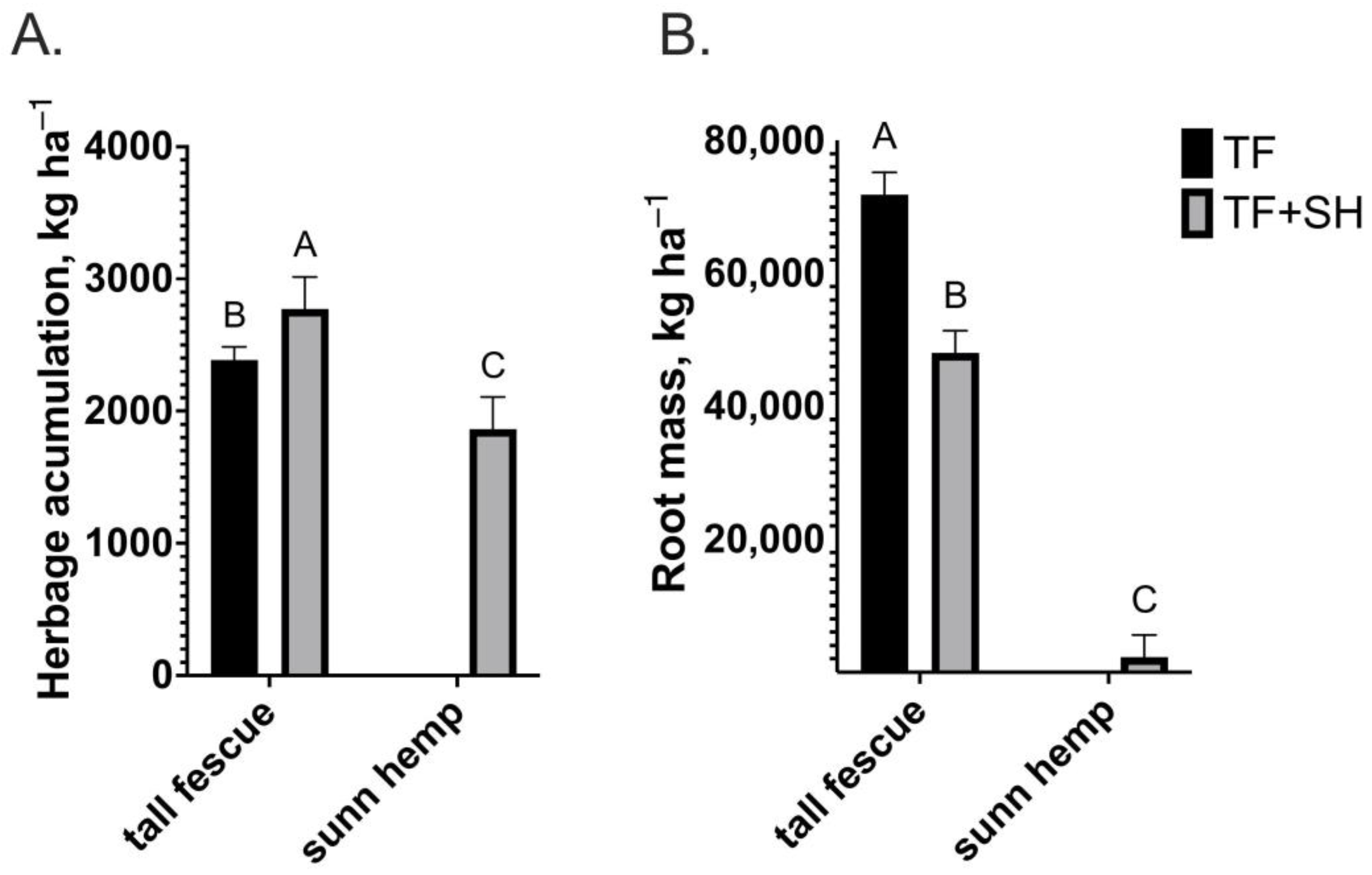
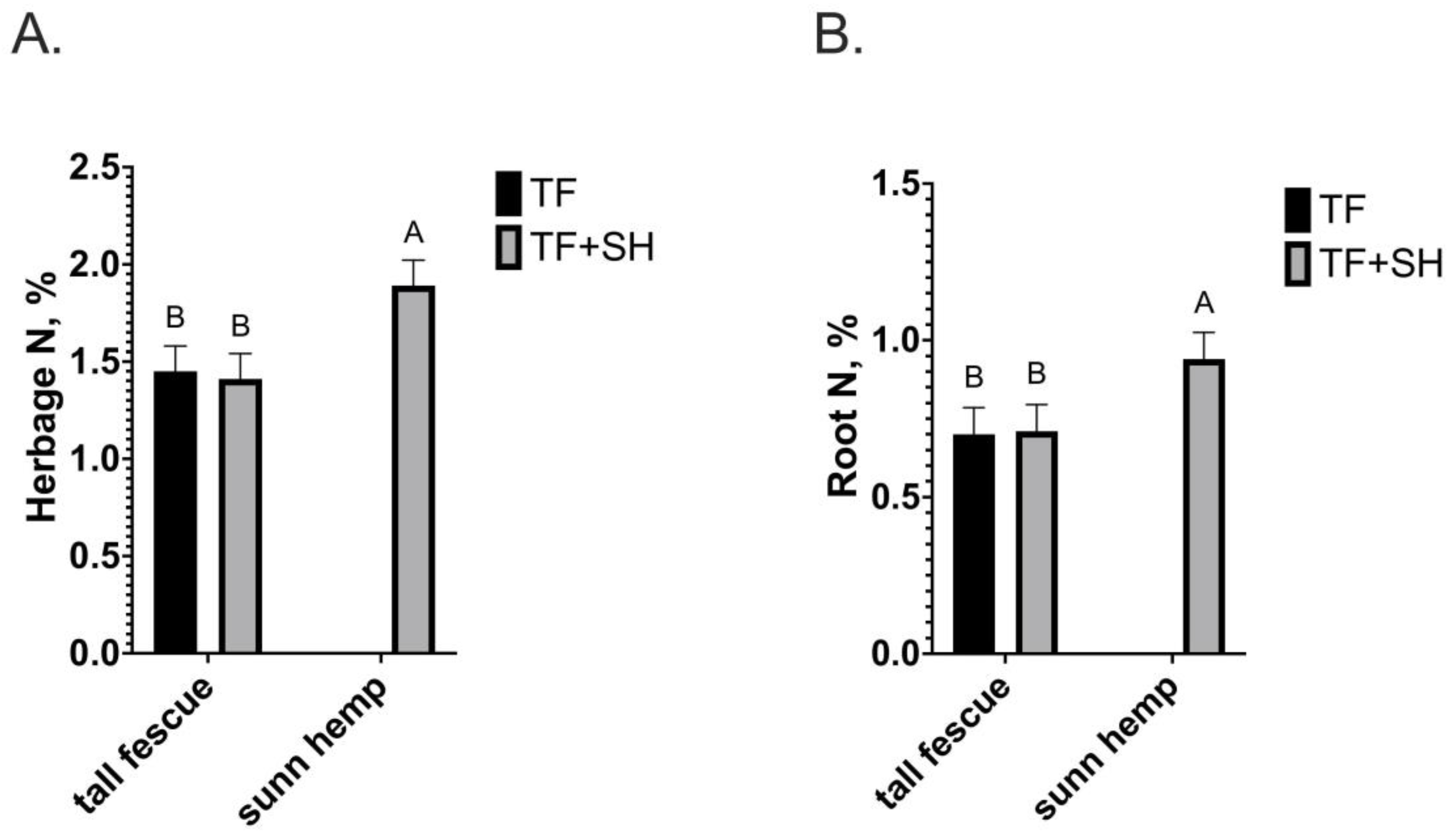
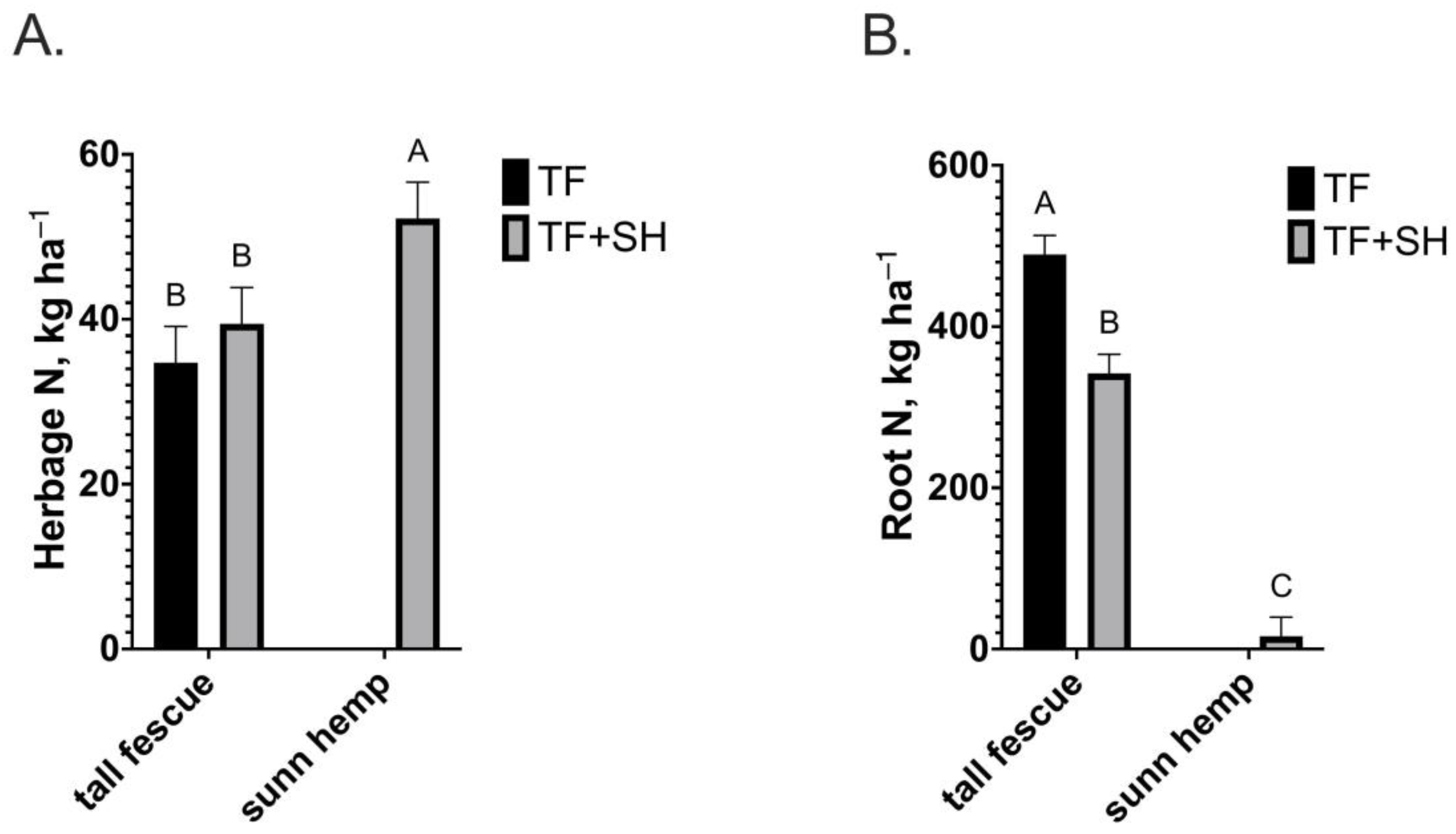
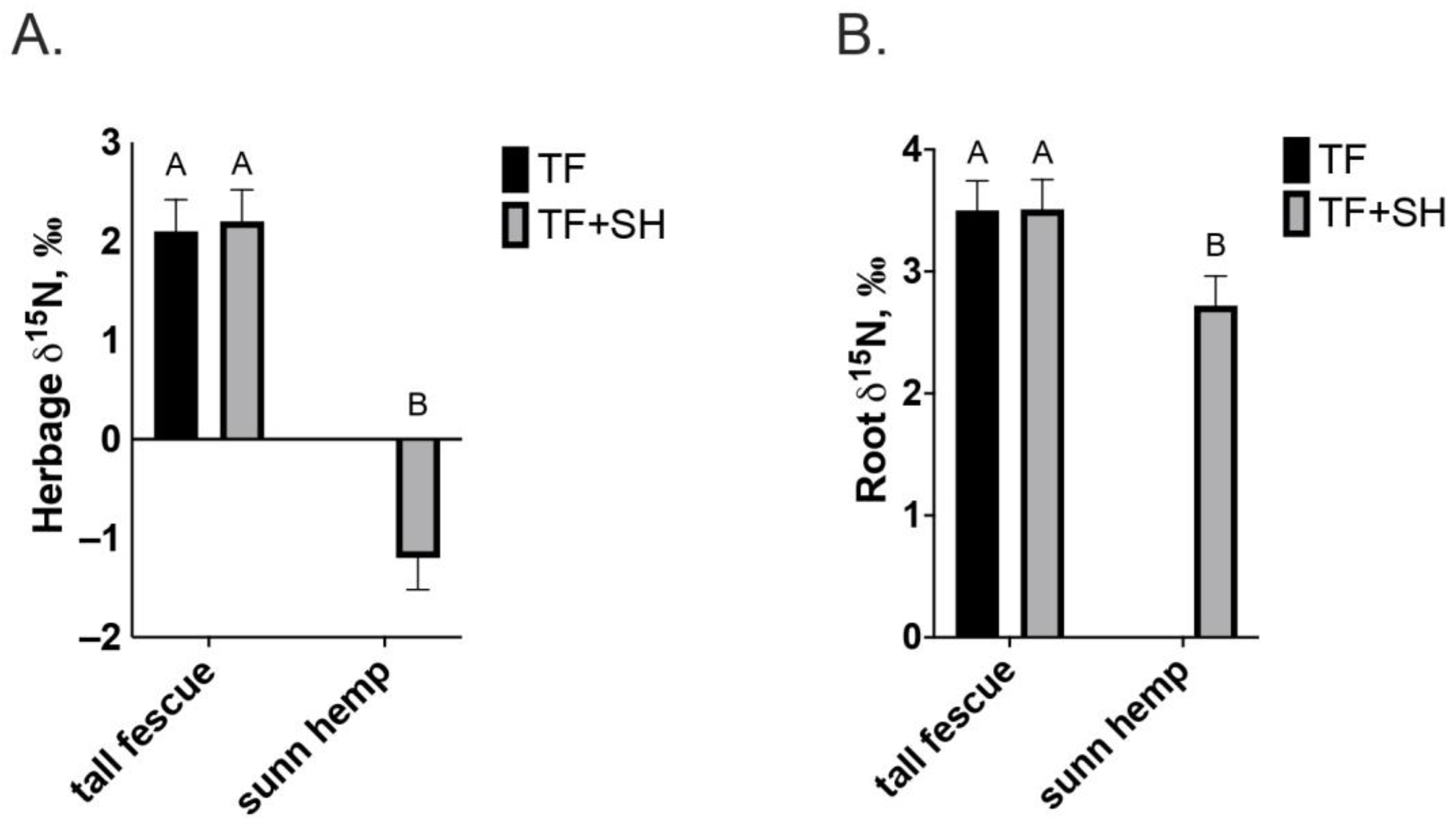
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BNF | Biological nitrogen fixation |
| HA | Herbage accumulation |
| N | Nitrogen |
| NDFA | Nitrogen derived from the atmosphere |
| Ntran | Nitrogen transfer |
| SH | Sunn hemp |
| TF | Tall fescue |
References
- Kubesch, J.O.C.; Nave, R.L.G.; Cui, S.; Bates, G.E.; Butler, D.M.; Pantalone, V. Transitional Organic Forage Systems in the Southeastern U.S.: Production and Nutritive Value. Agron. J. 2022, 114, 1269–1283. [Google Scholar] [CrossRef]
- Min, D.-H.; Moyer, J.L. Establishing Legumes in a Tall Fescue Sward. Am. J. Plant Sci. 2015, 06, 355–361. [Google Scholar] [CrossRef]
- Mullenix, K.; Dillard, S.; Severino da Silva, L.; Dubeux, J., Jr.; Tucker, J.; Baxter, L.; Prevatt, C.; Santos, E.; Garcia, L. Forage Legumes in the Southeast; Clemson University Cooperative Extension Service: Pickens County, SC, USA, 2023. [Google Scholar]
- Evers, G. The Interaction of Annual Ryegrass and Nitrogen on Arrowleaf Clover in the Southeastern United States. Crop Sci. 2011, 51, 1353–1360. [Google Scholar] [CrossRef]
- MacMillan, J.; Adams, C.B.; Hinson, P.O.; DeLaune, P.B.; Rajan, N.; Trostle, C. Biological Nitrogen Fixation of Cool-season Legumes in Agronomic Systems of the Southern Great Plains. Agrosyst. Geosci. Environ. 2022, 5, e20244. [Google Scholar] [CrossRef]
- Rouquette, F.M.; Smith, G.R. Review: Effects of Biological Nitrogen Fixation and Nutrient Cycling on Stocking Strategies for Cow-Calf and Stocker Programs. Prof. Anim. Sci. 2010, 26, 131–141. [Google Scholar] [CrossRef]
- Dave, K.; Kumar, A.; Dave, N.; Jain, M.; Dhanda, P.S.; Yadav, A.; Kaushik, P. Climate Change Impacts on Legume Physiology and Ecosystem Dynamics: A Multifaceted Perspective. Sustainability 2024, 16, 6026. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Randall, H.C. Nitrogen and Biomass Accumulation by Alfalta under High Temperatures of Late Summer. Field Crops Res. 1993, 31, 287–294. [Google Scholar] [CrossRef]
- Ashworth, A.J.; West, C.P.; Allen, F.L.; Keyser, P.D.; Weiss, S.A.; Tyler, D.D.; Taylor, A.M.; Warwick, K.L.; Beamer, K.P. Biologically Fixed Nitrogen in Legume Intercropped Systems: Comparison of Nitrogen-Difference and Nitrogen-15 Enrichment Techniques. Agron. J. 2015, 107, 2419–2430. [Google Scholar] [CrossRef]
- Blesh, J. Functional Traits in Cover Crop Mixtures: Biological Nitrogen Fixation and Multifunctionality. J. Appl. Ecol. 2018, 55, 38–48. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology and Development, 6th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2015; ISBN 978-1-60535-255-8. [Google Scholar]
- Gadberry, M.S.; Hawley, J.; Beck, P.A.; Jennings, J.A.; Kegley, E.B.; Coffey, K.P. BILL E. KUNKLE INTERDISCIPLINARY BEEF SYMPOSIUM: A Meta-Analysis of Research Efforts Aimed at Reducing the Impact of Fescue Toxicosis on Cattle Weight Gain and Feed Intake1. J. Anim. Sci. 2015, 93, 5496–5505. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.G.; White, G.A. Crotalaria Juncea: A Potential Multi-Purpose Fiber Crop. In Progress in New Crops; ASHS Press: Arlington, VA, USA, 1996; pp. 389–394. [Google Scholar]
- Jaramillo, D.M.; Dubeux, J.C.B.; Vendramini, J.M.B.; Queiroz, L.M.D.; Santos, E.R.S.; Ruiz-Moreno, M.; Garcia, L.; De Abreu, D.S.; De Miranda, L.R.; De Siqueira, M.C.F. Establishment Techniques Affect Productivity, Nutritive Value and Atmospheric N2 Fixation of Two Sunn Hemp Cultivars. Grass Forage Sci. 2020, 75, 153–158. [Google Scholar] [CrossRef]
- Eberle, C.; Shortnacy, L. Sunn Hemp Planting Date Effect on Growth, Biomass Accumulation, and Nutritive Value in Southeastern Wyoming. Crop Sci. 2021, 61, 4447–4457. [Google Scholar] [CrossRef]
- Kaneko, M.; Da Silva, H.M.S.; Vendramini, J.M.B.; Sanchez, J.M.D.; De Sousa, J.L.; De Oliveira, R.A. Hybrid Brachiariagrass (Urochloa Spp.) Establishment with Annual Sorghum Sudangrass (Sorghum bicolor (L.) Moench) and Sunn Hemp (Crotalaria juncea L.) Mixtures at Half and Full Recommended Seeding Rates. Grassl. Sci. 2023, 69, 87–95. [Google Scholar] [CrossRef]
- Corbin, M.D.; Nave, R.L.G.; Naumann, H.D.; Bates, G.E.; Boyer, C.; De Almeida, O.G. Inclusion of Cool- and Warm-season Species in Tall Fescue Swards for Increased Productivity. Agron. J. 2024, agj2.21690. [Google Scholar] [CrossRef]
- Garzon, J.; Vendramini, J.M.B.; Silveira, M.L.; Moriel, P.; Da Silva, H.M.S.; Dubeux, J.C.B.; Kaneko, M.; Carnelos, C.C.; Mamede, P.A. Harvest Management and Genotype Effects on Sunn Hemp Forage Characteristics. Agron. J. 2021, 113, 298–307. [Google Scholar] [CrossRef]
- Mansoer, Z.; Reeves, D.W.; Wood, C.W. Suitability of Sunn Hemp as an Alternative Late-Summer Legume Cover Crop. Soil Sci. Soc. Amer. J. 1997, 61, 246–253. [Google Scholar] [CrossRef]
- Balkcom, K.S.; Reeves, D.W. Sunn-Hemp Utilized as a Legume Cover Crop for Corn Production. Agron. J. 2005, 97, 26–31. [Google Scholar] [CrossRef]
- Rotar, P.P.; Joy, R.J. “Tropic Sun” Sunn Hemp; Crotalaria Juncea L.; Research Extension Series; RES-36; University of Hawaii: Honolulu, HI, USA, 1983; 7p. [Google Scholar]
- Schomberg, H.H.; Martini, N.L.; Diaz-Perez, J.C.; Phatak, S.C.; Balkcom, K.S.; Bhardwaj, H.L. Potential for Using Sunn Hemp as a Source of Biomass and Nitrogen for the Piedmont and Coastal Plain Regions of the Southeastern USA. Agron. J. 2007, 99, 1448–1457. [Google Scholar] [CrossRef]
- Brophy, L.S.; Heichel, G.H.; Russelle, M.P. Nitrogen Transfer from Forage Legumes to Grass in a Systematic Planting Design1. Crop Sci. 1987, 27, 753–758. [Google Scholar] [CrossRef]
- Luo, F.; Mi, W.; Liu, W. Legume–Grass Mixtures Improve Biological Nitrogen Fixation and Nitrogen Transfer by Promoting Nodulation and Altering Root Conformation in Different Ecological Regions of the Qinghai–Tibet Plateau. Front. Plant Sci. 2024, 15, 1375166. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; McElroy, M.S.; Chapagain, T.; Papadopoulos, Y.A.; Raizada, M.N. Belowground Nitrogen Transfer from Legumes to Non-Legumes under Managed Herbaceous Cropping Systems. A Review. Agron. Sustain. Dev. 2016, 36, 58. [Google Scholar] [CrossRef]
- Chalk, P.M.; Peoples, M.B.; McNeill, A.M.; Boddey, R.M.; Unkovich, M.J.; Gardener, M.J.; Silva, C.F.; Chen, D. Methodologies for Estimating Nitrogen Transfer between Legumes and Companion Species in Agro-Ecosystems: A Review of 15N-Enriched Techniques. Soil Biol. Biochem. 2014, 73, 10–21. [Google Scholar] [CrossRef]
- Moyer-Henry, K.A.; Burton, J.W.; Israel, D.W.; Rufty, T.W. Nitrogen Transfer Between Plants: A 15N Natural Abundance Study with Crop and Weed Species. Plant Soil 2006, 282, 7–20. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon and Organic Matter. In Methods of Soil Analysis. Part 2; Page, A.L., Miller, R.H., Kenney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 539–579. [Google Scholar]
- Measuring Plant-Associated Nitrogen Fixation in Agricultural Systems; Unkovich, M., Cadisch, G., Australian Centre for International Agricultural Research, Eds.; ACIAR monograph series; ACIAR: Canberra, Australia, 2008; ISBN 978-1-921531-26-2. [Google Scholar]
- Okito, A.; Alves, B.R.J.; Urquiaga, S.; Boddey, R.M. Isotopic Fractionation during N2 Fixation by Four Tropical Legumes. Soil Biol. Biochem. 2004, 36, 1179–1190. [Google Scholar] [CrossRef]
- Frankow-Lindberg, B.E.; Dahlin, A.S. N2 Fixation, N Transfer, and Yield in Grassland Communities Including a Deep-Rooted Legume or Non-Legume Species. Plant Soil 2013, 370, 567–581. [Google Scholar] [CrossRef]
- Corbin, M.D.; Nave, R.L.G.; Bates, G.E.; Butler, D.M.; Hawkins, S.A. Alternatives to Conventional Nitrogen Fertilization on Tall Fescue and Bermudagrass. Agron. J. 2019, 111, 275–286. [Google Scholar] [CrossRef]
- Dhamala, N.R.; Eriksen, J.; Carlsson, G.; Søegaard, K.; Rasmussen, J. Highly Productive Forage Legume Stands Show No Positive Biodiversity Effect on Yield and N2-Fixation. Plant Soil 2017, 417, 169–182. [Google Scholar] [CrossRef]
- Kubesch, J.O.C.; Greiner, S.P.; Pent, G.J.; Reid, J.L.; Tracy, B.F. Diversifying Tall Fescue Pastures with Native Warm-season Grasses and White Clover Improves Beef Cattle Performance. Crop Sci. 2025, 65, e70030. [Google Scholar] [CrossRef]
- Sheaffer, C.C.; Seguin, P.; Cuomo, G.J. Sward Characteristics and Management Effects on Cool-Season Grass Forage Quality. In Grass for Dairy Catrtle; CABI Publishing: New York, NY, USA, 1998; pp. 73–100. ISBN 978-0-85199-288-4. [Google Scholar]
- Kendall, W.A.; Shaffer, J.A.; Hill, R.R. Effect of Temperature and Water Variables on the Juvenile Growth of Lucerne and Red Clover. Grass Forage Sci. 1994, 49, 264–269. [Google Scholar] [CrossRef]
- Lepcha, I.; Naumann, H.D.; Fritschi, F.B.; Kallenbach, R.L. Herbage Accumulation, Nutritive Value, and Regrowth Potential of Sunn Hemp at Different Harvest Regimens and Maturity. Crop Sci. 2019, 59, 413–421. [Google Scholar] [CrossRef]
- Luo, C.L.; Duan, H.X.; Wang, Y.L.; Liu, H.J.; Xu, S.X. Complementarity and Competitive Trade-Offs Enhance Forage Productivity, Nutritive Balance, Land and Water Use, and Economics in Legume-Grass Intercropping. Field Crops Res. 2024, 319, 109642. [Google Scholar] [CrossRef]
- Spasiani, P.P.; Homem, B.G.C.; de Lima, I.B.G.; Guimarães, B.C.; de Medeiros, E.S.; Muir, J.P.; de Oliveira, M.S.; Boddey, R.M.; Casagrande, D.R. Light Competition Is the Key Factor Determining Spatio-Temporal Variability in Legume Proportion within Marandu Palisadegrass–Forage Peanut Mixed Pastures. Crop Pasture Sci. 2023, 74, 898–910. [Google Scholar] [CrossRef]
- Huang, B.; Gao, H. Root Physiological Characteristics Associated with Drought Resistance in Tall Fescue Cultivars. Crop Sci. 2000, 40, 196–203. [Google Scholar] [CrossRef]
- Abdul-baki, A.A.; Bryan, H.H.; Zinati, G.M.; Klassen, W.; Codallo, M.; Heckert, N. Biomass Yield and Flower Production in Sunn Hemp: Effect of Cutting the Main Stem. J. Veg. Crop Prod. 2001, 7, 83–104. [Google Scholar] [CrossRef]
- Garzon, J.; Vendramini, J.M.B.; Silveira, M.L.; Dubeux, J.C.; Liao, H.; Sollenberger, L.E.; Moriel, P.; Silva, H.M.S.; Gomes, V.C.; Ferreira, I.M.; et al. Residue Management and Genotype Effect on Sunn Hemp Organic Matter and Nitrogen Decomposition. Agron. J. 2023, 115, 261–272. [Google Scholar] [CrossRef]
- Sant’Anna, S.A.C.; Martins, M.R.; Goulart, J.M.; Araújo, S.N.; Araújo, E.S.; Zaman, M.; Jantalia, C.P.; Alves, B.J.R.; Boddey, R.M.; Urquiaga, S. Biological Nitrogen Fixation and Soil N2O Emissions from Legume Residues in an Acrisol in SE Brazil. Geoderma Reg. 2018, 15, e00196. [Google Scholar] [CrossRef]
- Dubeux, J.C.B.; Blount, A.R.S.; Mackowiak, C.; Santos, E.R.S.; Pereira Neto, J.D.; Riveros, U.; Garcia, L.; Jaramillo, D.M.; Ruiz-Moreno, M. Biological N 2 Fixation, Belowground Responses, and Forage Potential of Rhizoma Peanut Cultivars. Crop Sci. 2017, 57, 1027–1038. [Google Scholar] [CrossRef]
- Dubeux, J.C.B.; Sollenberger, L.E.; Mathews, B.W.; Scholberg, J.M.; Santos, H.Q. Nutrient Cycling in Warm-Climate Grasslands. Crop Sci. 2007, 47, 915–928. [Google Scholar] [CrossRef]
- Russelle, M.P. Nitrogen Cycling in Pasture and Range. J. Prod. Agric. 1992, 5, 13–23. [Google Scholar] [CrossRef]
- Lepcha, I.; Naumann, H.D. Partitioning of Forage Mass and Nutritive Value in Sunn Hemp Leaf and Stem Components. Int. J. Agron. 2021, 2021, 5547120. [Google Scholar] [CrossRef]
- Stallings, A.M.; Balkcom, K.S.; Wood, C.W.; Guertal, E.A.; Weaver, D.B. Nitrogen Mineralization from ‘AU Golden’ Sunn Hemp Residue. J. Plant Nutr. 2017, 40, 50–62. [Google Scholar] [CrossRef]
- Høgh-Jensen, H.; Schjoerring, J.K. Interactions between White Clover and Ryegrass under Contrasting Nitrogen Availability: N2 Fixation, N Fertilizer Recovery, N Transfer and Water Use Efficiency. Plant Soil 1997, 197, 187–199. [Google Scholar] [CrossRef]
- Tracy, B.F.; Albrecht, K.; Flores, J.; Hall, M.; Islam, A.; Jones, G.; Lamp, W.; MacAdam, J.W.; Skinner, H.; Teutsch, C. Evaluation of Alfalfa–Tall Fescue Mixtures across Multiple Environments. Crop Sci. 2016, 56, 2026–2034. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Steele, K.W. Biological Nitrogen Fixation in Mixed Legume/Grass Pastures. In Biological Nitrogen Fixation for Sustainable Agriculture; Ladha, J.K., George, T., Bohlool, B.B., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1992; pp. 137–153. ISBN 978-90-481-4164-7. [Google Scholar]
- Carlsson, G.; Huss-Danell, K. Nitrogen Fixation in Perennial Forage Legumes in the Field. Plant Soil 2003, 253, 353–372. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Midega, C.A.O.; Awuor, R.; Nyagol, D.; Khan, Z.R. Perennial Legume Intercrops Provide Multiple Belowground Ecosystem Services in Smallholder Farming Systems. Agric. Ecosyst. Environ. 2021, 320, 107566. [Google Scholar] [CrossRef]
- Meza, K.; Vanek, S.J.; Sueldo, Y.; Olivera, E.; Ccanto, R.; Scurrah, M.; Fonte, S.J. Grass–Legume Mixtures Show Potential to Increase Above- and Belowground Biomass Production for Andean Forage-Based Fallows. Agronomy 2022, 12, 142. [Google Scholar] [CrossRef]
- Reilly, E.C.; Gutknecht, J.L.; Tautges, N.E.; Sheaffer, C.C.; Jungers, J.M. Nitrogen Transfer and Yield Effects of Legumes Intercropped with the Perennial Grain Crop Intermediate Wheatgrass. Field Crops Res. 2022, 286, 108627. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Drinkwater, L.E. Nitrogen Fixation in Annual and Perennial Legume-Grass Mixtures across a Fertility Gradient. Plant Soil 2012, 357, 147–159. [Google Scholar] [CrossRef]
- Santos, E.R.S.; Dubeux, J.C.B.; Mackowiak, C.L.; Blount, A.R.S.; Sollenberger, L.E.; Jaramillo, D.M.; Garcia, L.; Abreu, D.S.; Souza, R.T.; Ruiz-Moreno, M. Herbage Responses and Nitrogen Agronomic Efficiency of Bermudagrass–Legume Mixtures. Crop Sci. 2021, 61, 3815–3829. [Google Scholar] [CrossRef]
- Belesky, D.P.; Wilkinson, S.R.; Dawson, R.N.; Elsner, J.E. Forage Production of a Tall Fescue Sod Intercropped with Sorghum × Sudangrass and Rye. Agron. J. 1981, 73, 657–660. [Google Scholar] [CrossRef]
- Reinbott, T.M.; Blevins, D.G. Multiyear Use of Killed Strips for Forage and Grain Sorghum Production in a Tall Fescue Pasture. J. Prod. Agric. 1995, 8, 354–359. [Google Scholar] [CrossRef]
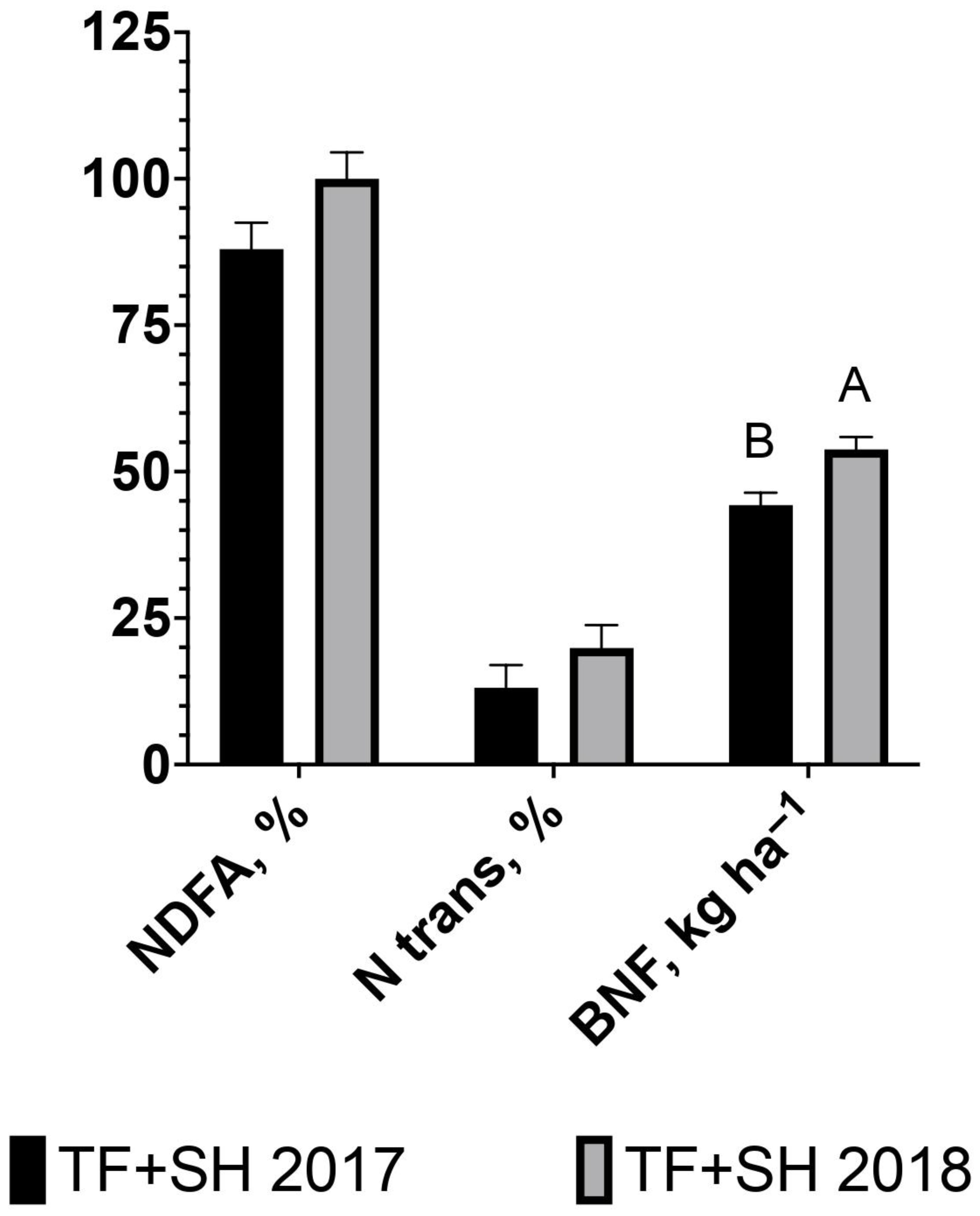

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Mean Rainfall (mm) | Mean Temperature (°C) | ||||
|---|---|---|---|---|---|---|
| 2017 | 2018 | 10-yr | 2017 | 2018 | 10-yr | |
| January | 30.5 | 22.9 | 29.7 | −1.1 | −3.5 | −2.9 |
| February | 3.6 | 66.3 | 50.4 | 5.5 | −0.7 | −2.2 |
| March | 98.6 | 49.3 | 76.2 | 7.4 | 4.6 | 6.5 |
| April | 122.4 | 18.3 | 101.2 | 13.6 | 7.5 | 11.9 |
| May | 69.3 | 119.6 | 108.4 | 16.7 | 21.9 | 17.1 |
| June | 182.9 | 89.2 | 162.7 | 22.4 | 24.4 | 22.6 |
| July | 92.2 | 39.4 | 117.3 | 24.9 | 24.8 | 24.2 |
| August | 80.8 | 148.3 | 95.9 | 21.1 | 24.5 | 23.6 |
| September | 21.1 | 95.3 | 109.5 | 20.8 | 20.7 | 19.1 |
| October | 126.2 | 146.6 | 76.6 | 13.7 | 12.2 | 12.6 |
| November | 9.4 | 41.7 | 62.4 | 5.7 | 1.0 | 6.3 |
| December | 13.7 | 79.2 | 50.3 | −1.3 | 0.8 | −0.7 |
| Total | 850.7 | 916.1 | 1040.6 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naumann, H.D.; Dubeux, J.C.B., Jr.; Tooley, J.A.; Lory, J.A.; Bretas, I.L. Nitrogen Dynamics in Sunn Hemp Intercropped Tall Fescue Pastures. Agronomy 2025, 15, 1027. https://doi.org/10.3390/agronomy15051027
Naumann HD, Dubeux JCB Jr., Tooley JA, Lory JA, Bretas IL. Nitrogen Dynamics in Sunn Hemp Intercropped Tall Fescue Pastures. Agronomy. 2025; 15(5):1027. https://doi.org/10.3390/agronomy15051027
Chicago/Turabian StyleNaumann, Harley D., José C. B. Dubeux, Jr., Joshua A. Tooley, John A. Lory, and Igor L. Bretas. 2025. "Nitrogen Dynamics in Sunn Hemp Intercropped Tall Fescue Pastures" Agronomy 15, no. 5: 1027. https://doi.org/10.3390/agronomy15051027
APA StyleNaumann, H. D., Dubeux, J. C. B., Jr., Tooley, J. A., Lory, J. A., & Bretas, I. L. (2025). Nitrogen Dynamics in Sunn Hemp Intercropped Tall Fescue Pastures. Agronomy, 15(5), 1027. https://doi.org/10.3390/agronomy15051027