
Agro-Physiological and Morphological Responses of Pearl Millet to Varying Water Regimes in Semi-Arid Conditions of Namibia

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
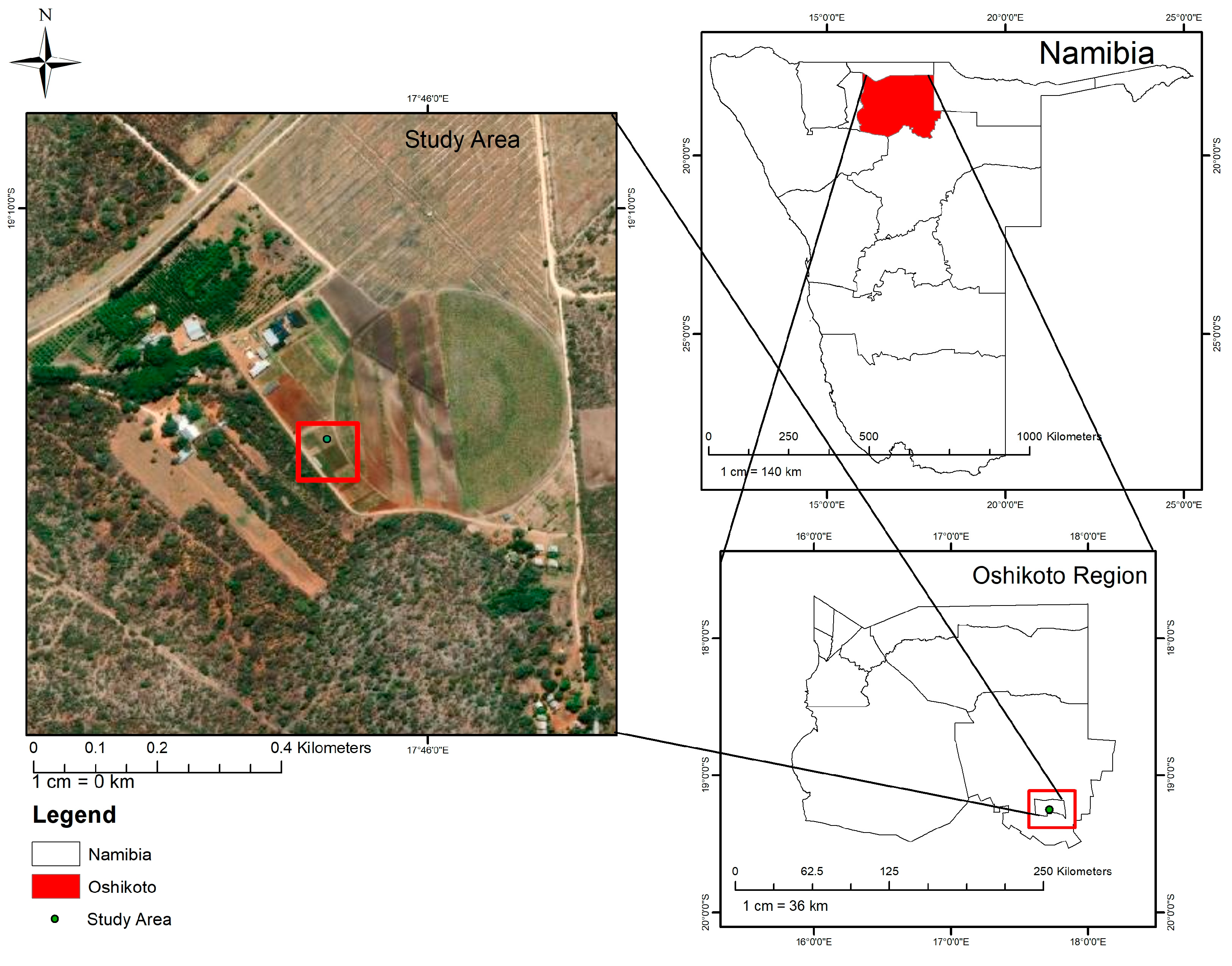
2.1. Study Area
2.2. Methodology
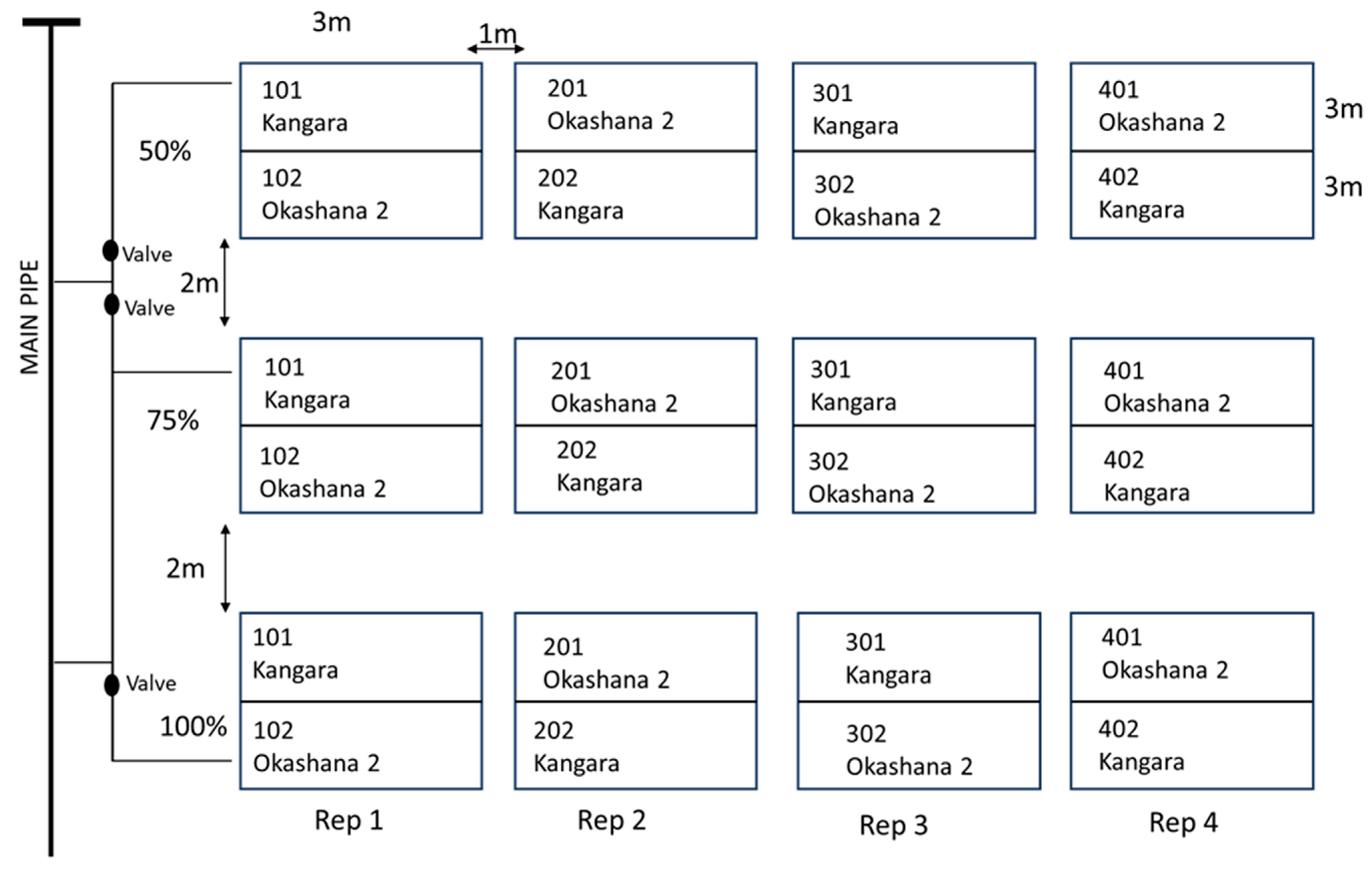
2.2.1. Experimental Design
2.2.2. Treatments Determination
2.2.3. Data Collection
Morphological Traits Data
Stomatal Conductance Measurements
Chlorophyll Content Measurements
Leaf Temperature Measurements
Yield and Yield Attributes Measurements
2.3. Data Analysis
3. Results
3.1. Effects of Water Regime, Variety, Season, and Their Interactions on the Morpho-Physiological Traits of Pearl Millet Varieties
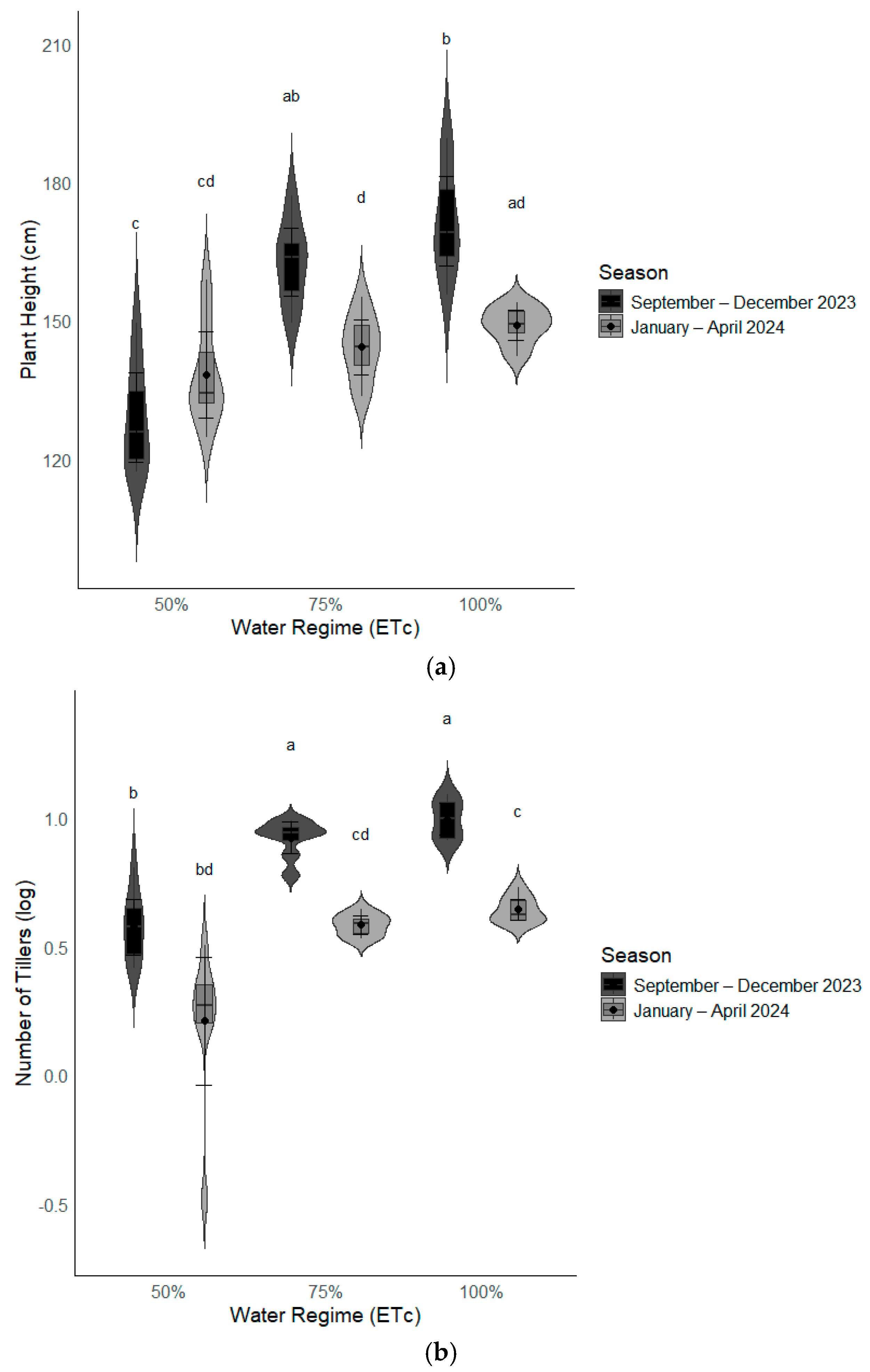
Interaction of Water Regime by Season on Pearl Millet Morpho-Physiological Characteristics
3.2. Effects of Water Regime, Variety, Season, and Their Interactions on the Yield and Yield Attributes of Pearl Millet
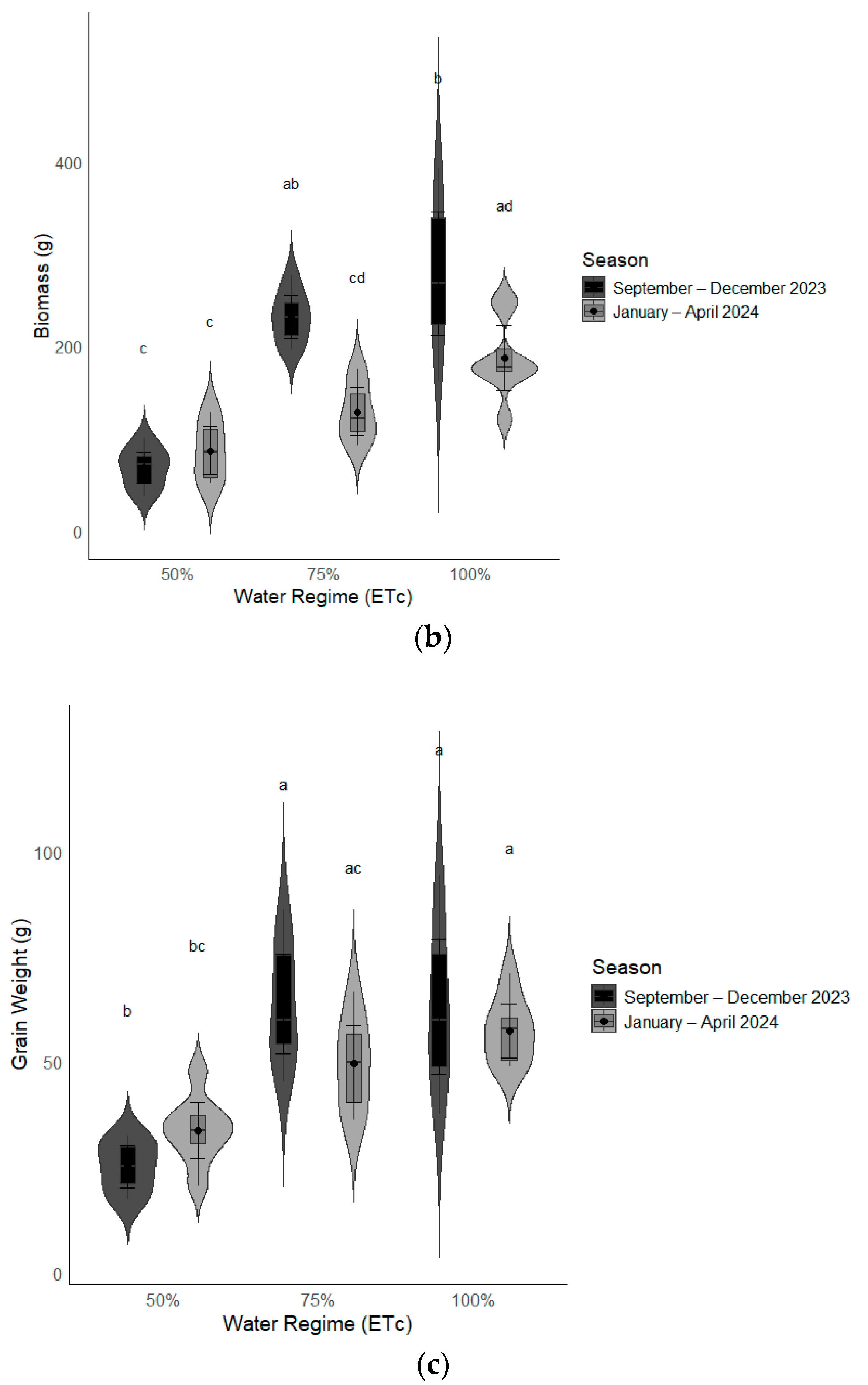
3.2.1. Interaction of Water Regime by Season on Pearl Millet Yield and Yield Characteristics
3.2.2. Changes in Yield and Yield Attributes of Pearl Millet Cultivars During the Two Studied Seasons
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Emediegwu, L.E.; Wossink, A.; Hall, A. The impacts of climate change on agriculture in sub-Saharan Africa: A spatial panel data approach. World Dev. 2022, 158, 105967. [Google Scholar] [CrossRef]
- UN-Water. Water and Climate Change. United Nations. 2024. Available online: https://www.unwater.org/water-facts/water-and-climate-change (accessed on 10 March 2024).
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–16. [Google Scholar]
- FAO. Agricultural Production Statistics 2000–2022. 2023. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/fba4ef43-422c-4d73-886e-3016ff47df52/content#page=2.41 (accessed on 10 December 2023).
- Choudhury, S.; Moulick, D.; Ghosh, D.; Soliman, M.; Alkhedaide, A.; Gaber, A.; Hossain, A. Drought-induced oxidative stress in pearl millet (Cenchrus americanus L.) at seedling stage: Survival mechanisms through alteration of morphophysiological and antioxidants activity. Life 2022, 12, 1171. [Google Scholar] [CrossRef]
- Kapoor, C.; Singh, S.P.; Sankar, S.M.; Singh, N. Enhancing drought tolerance in pearl millet (Pennisetum glaucum L.): Integrating traditional and omics approaches. Euphytica 2022, 218, 104. [Google Scholar] [CrossRef]
- Azare, I.M.; Dantata, I.J.; Abdullahi, M.S.; Adebayo, A.A.; Aliyu, M. Effects of climate change on pearl millet (Pennisetum glaucum [LR Br.]) production in Nigeria. J. Appl. Sci. Environ. Manag. 2020, 24, 157–162. [Google Scholar]
- Siroha, A.K.; Sandhu, K.S.; Kaur, M. Physicochemical, functional and antioxidant properties of flour from pearl millet varieties grown in India. J. Food Meas. Charact. 2016, 10, 311–318. [Google Scholar] [CrossRef]
- Tou, E.H.; Mouquet-Rivier, C.; Picq, C.; Traoré, A.S.; Trèche, S.; Guyot, J.-P. Improving the nutritional quality of ben-saalga, a traditional fermented millet-based gruel, by co-fermenting millet with groundnut and modifying the processing method. LWT-Food Sci. Technol. 2007, 40, 1561–1569. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Reddy, K.N.; Gowda CL, L. Pearl millet germplasm at ICRISAT genebank-status and impact. J. SAT Agric. Res. 2007, 3, 5. [Google Scholar]
- Ateeq, M.; Ahmad, M.; Shahzadi, K.; Ullah, R.; Murad, K.; Ansari, T.A. Minimizing the Effect of Low Moisture Stress in Pearl Millet (Pennisetum glaucum L.) by Regulating Growth, Yield and Antioxidant Defense System Via Foliar Applied Silicon. Asian J. Res. Crop Sci. 2024, 9, 124–138. [Google Scholar] [CrossRef]
- Bello, Z.A.; Walker, S.; Tesfuhuney, W. Water relations and productivity of two lines of pearl millet grown on lysimeter with two different soil types. Agric. Water Manag. 2019, 221, 528–537. [Google Scholar] [CrossRef]
- Lira JB d Andrade AP d Rodrigues Magalhaes, A.L.; Campos, F.S.; Araújo, G.G.L.d.; Deon, D.S.; Gois, G.C.; Regitano Neto, A.; Cunha, D.d.S.; Tabosa, J.N. Production of pearl millet irrigated with different levels of brackish water and organic matter. Commun. Soil Sci. Plant Anal. 2020, 51, 701–709. [Google Scholar]
- Lincoln, N.K.; Haensel, T.P.; Lee, T.M. Modeling Hawaiian Agroecology: Depicting traditional adaptation to the world’s most diverse environment. Front. Sustain. Food Syst. 2023, 7, 1116929. [Google Scholar] [CrossRef]
- Darmaun, M.; Chevallier, T.; Hossard, L.; Lairez, J.; Scopel, E.; Chotte, J.-L.; Lambert-Derkimba, A.; De Tourdonnet, S. Multidimensional and multiscale assessment of agroecological transitions. A review. Int. J. Agric. Sustain. 2023, 21, 2193028. [Google Scholar] [CrossRef]
- Byers, B.A. Environmental Threats and Opportunities in Namibia: A Comprehensive Assessment; Research Discussion Paper 21; Directorate of Environmental Affairs, Ministry of Environment and Tourism: Windhoek, Namibia, 1997. [Google Scholar]
- Lange, G.-M. An approach to sustainable water management in Southern Africa using natural resource accounts: The experience in Namibia. Ecol. Econ. 1998, 26, 299–311. [Google Scholar] [CrossRef]
- Namibia Water Corporation Ltd. (NamWater). Hydrological Services. Water Resources. Available online: https://www.namwater.com.na/index.php/services/56-hydrological-services?showall=1 (accessed on 30 March 2024).
- Walworth, J.L. Soil Sampling and Analysis; The University of Arizona: Tucson, AZ, USA, 2011. [Google Scholar]
- Smith, M.; Allen, R.; Pereira, L. Revised FAO Methodology for Crop-Water Requirements; FAO: Rome, Italy, 1998. [Google Scholar]
- National Aeronautics and Space Administration (NASA). 2023. Available online: https://power.larc.nasa.gov/data-access-viewer/ (accessed on 15 August 2023).
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Italy, Rome, 1998; Volume 300, p. D05109. [Google Scholar]
- Smith, M. CLIMWAT for CROPWAT. A Climatic Database for Irrigation Planning and Management; FAO: Rome, Italy, 1993. [Google Scholar]
- IBPGR and ICRISAT. Descriptors for Pearl millet [Pennisetum glaucum (L.) R. Br.]; International Board for Plant Genetic Resources: Rome, Italy; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 1993; p. 43. [Google Scholar]
- Konica Minolta, Inc. Chlorophyll Meter SPAD-502 Plus. Instruction Manual. 2009-2013. Tokyo. Japan. Available online: https://www.konicaminolta.com/instruments/download/instruction_manual/color/pdf/spad-502plus_instruction_eng.pdf (accessed on 30 August 2023).
- Zhang, K.; Liu, X.; Tahir Ata-Ul-Karim, S.; Lu, J.; Krienke, B.; Li, S.; Cao, Q.; Zhu, Y.; Cao, W.; Tian, Y. Development of chlorophyll-meter-index-based dynamic models for evaluation of high-yield japonica rice production in Yangtze River reaches. Agronomy 2019, 9, 106. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2021. Available online: https://www.r-project.org/ (accessed on 12 June 2024).
- Sakamoto, Y.; Ishiguro, M.; Kitagawa, G. Akaike Information Criterion Statistics; D. Reidel: Dordrecht, The Netherlands, 1986; Volume 81, p. 26853. [Google Scholar]
- Martınez, H.D.R. Analysing Interactions of Fitted Models. 2015. Available online: http://r.meteo.uni.wroc.pl/web/packages/phia/vignettes/phia.pdf (accessed on 10 June 2024).
- Londhe, V.M.; Birajdar, S.G.; Jadhav, V.T.; Jadhav, J.D.; Amrutsagar, V.M. Effect of Weather Parameters on Pearl Millet (Pennisetum glaucum L.) Cultivars under Different Sowing Windows. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 1190–1202. [Google Scholar] [CrossRef]
- Chavarria, G.; dos Santos, H.P. Plant Water Relations: Absorption, Transport and Control Mechanisms; IntechOpen: London, UK, 2012. [Google Scholar]
- Saini, A.K.; Patel, A.M.; Chaudhary, P.P.; Saini, L.H. Impact assessment of irrigation, fertility and hydrogel levels on growth attributes, yield and economics of summer pearl millet (Pennisetum glaucum L.) under North Gujarat conditions. J. Pharmacogn. Phytochem. 2018, 7, 2914–2918. [Google Scholar]
- Yadav, P.; Dagar, C.S.; Kumar, A.; Dahiya, P. Effect of soil moisture on growth and yield of pearl millet (Pennisetum glaucum L.) under different dates of sowing. Environ. Ecol. 2022, 40, 1489–1496. [Google Scholar]
- Ausiku, A.P.; Annandale, J.G.; Steyn, J.M.; Sanewe, A.J. Improving Pearl Millet (Pennisetum glaucum) productivity through adaptive management of water and nitrogen. Water 2020, 12, 422. [Google Scholar] [CrossRef]
- Phoura, Y.; Ohtomo, R.; Nakanishi, H.; Kamoshita, A. Effects of arbuscular mycorrhizal fungi inoculation on infection and growth of rice and pearl millet in upland fields with three water regimes. Plant Prod. Sci. 2023, 26, 350–363. [Google Scholar] [CrossRef]
- Yadav, N.; Kumar, A.; Kumar, N.; Kumar, S.; Arya, S.S. Impacts on Plant Growth and Development Under Stress. In Plant Stress Mitigators; Springer: Berlin/Heidelberg, Germany, 2022; pp. 61–100. [Google Scholar]
- Beveridge, C.A.; Rameau, C.; Wijerathna-Yapa, A. Lessons from a century of apical dominance research. J. Exp. Bot. 2023, 74, 3903–3922. [Google Scholar] [CrossRef] [PubMed]
- Vadez, V.; Hash, T.; Bidinger, F.R.; Kholova, J., II. 1.5 Phenotyping pearl millet for adaptation to drought. Front. Physiol. 2012, 3, 386. [Google Scholar] [CrossRef]
- Muller, J.D.; Rotenberg, E.; Tatarinov, F.; Oz, I.; Yakir, D. Evidence for efficient nonevaporative leaf-to-air heat dissipation in a pine forest under drought conditions. New Phytol. 2021, 232, 2254–2266. [Google Scholar] [CrossRef]
- Karthika, K.J.D.; Manivannan, V.; Dheebakaran, G.; Kokilavani, S.; Djanaguiraman, M. Effect of Microclimate Modification on Growth and Yield of Pearl Millet. Int. J. Environ. Clim. Change 2022, 12, 893–898. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Shukla, S.S.; Panda, S.K. Nitrogen Use Efficiency Regulates Drought Stress in Pearl Millet Genotypes: Morpho-Physiological Evaluation. Agriculture 2023, 13, 680. [Google Scholar] [CrossRef]
- Wasaya, A.; Hassan, J.; Yasir, T.A.; Ateeq, M.; Raza, M.A. Foliar application of silicon improved physiological indicators, yield attributes, and yield of pearl millet (Pennisetum glaucum L.) under terminal drought stress. J. Soil Sci. Plant Nutr. 2022, 22, 4458–4472. [Google Scholar] [CrossRef]
- Shanker, A.K.; Amirineni, S.; Bhanu, D.; Yadav, S.K.; Jyothilakshmi, N.; Vanaja, M.; Singh, J.; Sarkar, B.; Maheswari, M.; Singh, V.K. High-resolution dissection of photosystem II electron transport reveals differential response to water deficit and heat stress in isolation and combination in pearl millet [Pennisetum glaucum (L.) R. Br.]. Front. Plant Sci. 2022, 13, 892676. [Google Scholar] [CrossRef]
- Del Pozo, A.; Méndez-Espinoza, A.M.; Romero-Bravo, S.; Garriga, M.; Estrada, F.; Alcaíno, M.; Camargo-Rodriguez, A.V.; Corke FM, K.; Doonan, J.H.; Lobos, G.A. Genotypic variations in leaf and whole-plant water use efficiencies are closely related in bread wheat genotypes under well-watered and water-limited conditions during grain filling. Sci. Rep. 2020, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- El-Sabagh, A.; Barutcular, C.; Islam, M.S. Relationships between stomatal conductance and yield under deficit irrigation in maize (Zea mays L.). J. Exp. Biol. Agric. Sci. 2017, 5, 14–21. [Google Scholar] [CrossRef]
- El-Tigani, S. Effect of Different Irrigation Levels on Growth and Photosynthetic Capacity of Four Pearl Millet Genotypes. NeuroQuantology 2022, 20, 2514. [Google Scholar]
- Kayalvizhi, A.; Karthikeyan, R.; Selvakumar, T.; Senthil, A.; Ramalakshmi, A. Effect of stress ameliorants on pearl millet (Pennisetum glaucum L.) growth and productivity under different moisture regimes. Pharma Innov. 2021, 10, 1723–1727. [Google Scholar]
- Salem EM, M.; Shoman, H.E.-D.A. Impact of Irrigation Water Quantities and Soil Mulching on Pearl Millet Performance under Heat Stress Conditions. Egypt. J. Agron. 2021, 43, 333–345. [Google Scholar] [CrossRef]
- Bakheit, B.; Ali MB, M.; Mahdy RE, E.; Kandeel, M.N. Assessment of Pearl Millet Genotypes for Seed Yield and its Attributes under Normal and Water Stress Conditions. Assiut J. Agric. Sci. 2021, 52, 28–52. [Google Scholar]
- Farooqi, Z.U.R.; Ayub, M.A.; ur Rehman, M.Z.; Sohail, M.I.; Usman, M.; Khalid, H.; Naz, K. Regulation of drought stress in plants. In Plant Life Under Changing Environment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 77–104. [Google Scholar]
- de Almeida, A.M.; Coelho, R.D.; da Silva Barros, T.H.; de Oliveira Costa, J.; Quiloango-Chimarro, C.A.; Moreno-Pizani, M.A.; Farias-Ramírez, A.J. Water productivity and canopy thermal response of pearl millet subjected to different irrigation levels. Agric. Water Manag. 2022, 272, 107829. [Google Scholar] [CrossRef]
- Tharanya, M.; Kholova, J.; Sivasakthi, K.; Thirunalasundari, T.; Vadez, V. Water-Conservation Traits to Increase Crop Yields in Water-Deficit Environments: Case Studies; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Qiao, M.; Hong, C.; Jiao, Y.; Hou, S.; Gao, H. Impacts of Drought on Photosynthesis in Major Food Crops and the Related Mechanisms of Plant Responses to Drought. Plants 2024, 13, 1808. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Chen, Y.; Meng, S.; Duan, M.; Zhang, J.; Ye, N. Environmental stimuli: A major challenge during grain filling in cereals. Int. J. Mol. Sci. 2023, 24, 2255. [Google Scholar] [CrossRef]
- Halli, H.M.; Angadi, S.; Kumar, A.; Govindasamy, P.; Madar, R.; El-Ansary, D.O.; Rashwan, M.A.; Abdelmohsen SA, M.; Abdelbacki AM, M.; Mahmoud, E.A. Influence of planting and irrigation levels as physical methods on maize root morphological traits, grain yield and water productivity in semi-arid region. Agronomy 2021, 11, 294. [Google Scholar] [CrossRef]
- Zou, Y.; Saddique, Q.; Ali, A.; Xu, J.; Khan, M.I.; Qing, M.; Azmat, M.; Cai, H.; Siddique KH, M. Deficit irrigation improves maize yield and water use efficiency in a semi-arid environment. Agric. Water Manag. 2021, 243, 106483. [Google Scholar] [CrossRef]
- Allamine, H.M.; Buyuktas, D.; Karaca, C.; Aydinsakir, K.; Erdurmus, C. Effect of regulated deficit irrigation on productivity, evapotranspiration and quality of grain sorghum. Irrig. Sci. 2023, 41, 277–293. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (ETc) | Total Water Applied (mm)—Season 1 (September to December 2023) | Total Water Applied (mm)—Season 2 (January to December 2024) |
|---|---|---|
| 50% | 304.77 | 239.34 |
| 75% | 457.16 | 359.02 |
| 100% | 609.55 | 478.69 |
| Model Term | DF | Plant Height (cm) | No. of Leaves | No. of Tillers | Chlorophyll Content (SPAD value) | Stomatal Conductance (mmol_m_2_s_) | Leaf Temperature (°C) |
|---|---|---|---|---|---|---|---|
| Water Regime (WR) | 2 | 3078.48 (<0.001) | 131.94 (<0.001) | 81.39 (<0.001) | 687.61 (<0.001) | 90,047.00 (<0.001) | 398.58 (<0.001) |
| Variety (V) | 1 | 349.25 (0.05) | 0.15 (0.807) | 0.18 (0.72) | 0.47 (0.88) | 4178.00 (0.26) | 0.25 (0.84) |
| Season (S) | 1 | 1333.47 (<0.001) | 234.53 (<0.001) | 201.38 (<0.001) | 1021.36 (<0.0010) | 2346.00 (0.40) | 47.73 (0.005) |
| WR × V | 2 | 71.67 (0.43) | 0.26 (0.90) | 0.08 (0.94) | 24.75 (0.27) | 2068.00 (0.53) | 0.15 (0.97) |
| WR × S | 2 | 1199.87 (<0.001) | 4.63 (0.172) | 13.47 (<0.001) | 35.35 (0.16) | 2300.00 (0.49) | 21.66 (0.03) |
| V × S | 1 | 191.82 (0.13) | 1.02 (0.53) | 2.12 (0.21) | 0.02 (0.93) | 1100.00 (0.56) | 0.43 (0.78) |
| WR × V × S | 2 | 76.71 (0.40) | 1.61 (0.53) | 0.17 (0.88) | 0.75 (0.96) | 334.00 (0.90) | 0.92 (0.85) |
| Residual | 36 | 81.94 | 2.51 | 1.32 | 18.42 | 3201.00 | 5.56 |
| Grand Mean | 149.18 | 10.97 | 5.43 | 44.45 | 297.61 | 34.27 | |
| CV (%) | 6.1 | 14.4 | 21.1 | 9.7 | 19.0 | 6.9 | |
| S.E± | 9.05 | 1.58 | 1.15 | 4.29 | 56.58 | 2.36 |
| Model Term | DF | ST (cm) | NP | PD (cm) | PL (cm) | DPW (g) | BM (g) | GW (g) | 1000-SW (g) |
|---|---|---|---|---|---|---|---|---|---|
| Water Regime (WR) | 2 | 0.12 (<0.001) | 18.51 (<0.001) | 0.34 (<0.001) | 5.71 (0.07) | 10,043.0 (<0.001) | 99,558 (<0.001) | 4521.0 (<0.001) | 80.58 (0.001) |
| Variety (V) | 1 | 0.01 (0.28) | 0.81 (0.15) | 0.01 (0.46) | 36.38 (<0.001) | 109.0 (0.56) | 2576 (0.25) | 21.0 (0.71) | 6.98 (0.03) |
| Season (S) | 1 | 0.0008 (0.76) | 7.32 (<0.001) | 0.58 (<0.001) | 1.08 (0.47) | 391.0 (0.28) | 41,167 (<0.001) | 169.0 (0.30) | 48.04 (<0.001) |
| WR × V | 2 | 0.004 (0.64) | 0.60 (0.22) | 0.002 (0.91) | 0.50 (0.78) | 80.0 (0.78) | 4814 (0.08) | 72.0 (0.63) | 3.24 (0.10) |
| WR × S | 2 | 0.009 (0.31) | 4.67 (<0.001) | 0.020.32 | 5.36 (0.09) | 681.0 (0.14) | 17863 (<0.001) | 533.0 (0.04) | 0.39 (0.74) |
| V ×S | 1 | 0.01 (0.22) | 0.03 (0.79) | 0.04 (0.13) | 2.05 (0.32) | 196.0 (0.44) | 68 (0.85) | 26.0 (0.68) | 1.67 (0.26) |
| WR × V × S | 2 | 0.001 (0.88) | 0.34 (0.42) | 0.02 (0.40) | 0.26 (0.88) | 416.0 (0.29) | 90 (0.95) | 138.0 (0.41) | 0.37 (0.75) |
| Residual | 36 | 0.008 | 0.38 | 0.02 | 2.05 | 321.0 | 1856 | 151.0 | 1.29 |
| Grand Mean | 0.58 | 2.95 | 2.51 | 20.71 | 70.75 | 164.17 | 48.88 | 9.72 | |
| CV (%) | 15.0 | 21.0 | 5.1 | 6.9 | 25.3 | 26.2 | 25.2 | 11.7 | |
| S.E ± | 0.09 | 0.62 | 0.13 | 1.43 | 17.93 | 43.08 | 12.30 | 1.14 |
| Parameter | Stem Thickness (cm) | Number of Productive Tillers | Panicle Diameter (cm) | Panicle Length (cm) | ||||
|---|---|---|---|---|---|---|---|---|
| Season | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 |
| A. Water Regime (ETc) | ||||||||
| 100 | 0.69 b | 0.66 b | 4.27 b | 3.14 b | 2.57 b | 2.75 b | 21.27 b | 20.71 |
| 75 | 0.55 a | 0.59 b | 4.24 b | 2.60 ab | 2.37 a | 2.67 b | 21.19 b | 21.03 |
| 50 | 0.52 a | 0.48 a | 1.50 a | 1.93 a | 2.28 a | 2.46 a | 19.21 a | 20.83 |
| F-test | 0.004 | <0.001 | <0.001 | 0.002 | 0.003 | <0.001 | 0.02 | 0.90 |
| B. Cultivar | ||||||||
| Kangara | 0.56 | 0.58 | 3.49 | 2.66 | 2.42 | 2.58 | 19.48 | 20.19 |
| Okashana 2 | 0.62 | 0.58 | 3.18 | 2.45 | 2.39 | 2.67 | 21.64 | 21.52 |
| F-test | 0.15 | 0.91 | 0.28 | 0.36 | 0.63 | 0.08 | 0.002 | 0.04 |
| C. Interaction | ||||||||
| A × B | 0.66 | 0.92 | 0.16 | 0.88 | 0.80 | 0.38 | 0.89 | 0.78 |
| Parameter | Dry Panicles Weight (g) | Biomass (g) | Grain Weight (g) | 1000-Seed Weight (g) | ||||
| Season | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 |
| A. Water Regime (ETc) | ||||||||
| 100 | 89.65 b | 88.89 b | 278.90 b | 187.59 b | 63.16 b | 57.52 b | 10.26 b | 12.04 b |
| 75 | 90.98 b | 70.48 b | 232.40 b | 129.68 a | 63.83 b | 49.61 b | 9.69 b | 12.05 b |
| 50 | 40.16 a | 44.31 a | 69.1 a | 87.38 a | 25.27 a | 33.90 a | 6.19 a | 8.07 a |
| F-test | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | <0.001 |
| B. Cultivar | ||||||||
| Kangara | 74.11 | 64.36 | 184.94 | 128.75 | 52.16 | 46.93 | 9.28 | 10.91 |
| Okashana 2 | 73.09 | 71.42 | 201.98 | 141.02 | 49.35 | 47.08 | 8.15 | 10.52 |
| F-test | 0.90 | 0.30 | 0.42 | 0.38 | 0.64 | 0.97 | 0.02 | 0.42 |
| C. Interaction | ||||||||
| A × B | 0.35 | 0.78 | 0.42 | 0.13 | 0.41 | 0.89 | 0.13 | 0.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moseki, O.; Kangueehi, G.; Chiteculo, V.; Zink, M.; Wanga, M.A. Agro-Physiological and Morphological Responses of Pearl Millet to Varying Water Regimes in Semi-Arid Conditions of Namibia. Agronomy 2025, 15, 381. https://doi.org/10.3390/agronomy15020381
Moseki O, Kangueehi G, Chiteculo V, Zink M, Wanga MA. Agro-Physiological and Morphological Responses of Pearl Millet to Varying Water Regimes in Semi-Arid Conditions of Namibia. Agronomy. 2025; 15(2):381. https://doi.org/10.3390/agronomy15020381
Chicago/Turabian StyleMoseki, Ofentse, Grace Kangueehi, Vasco Chiteculo, Matthias Zink, and Maliata Athon Wanga. 2025. "Agro-Physiological and Morphological Responses of Pearl Millet to Varying Water Regimes in Semi-Arid Conditions of Namibia" Agronomy 15, no. 2: 381. https://doi.org/10.3390/agronomy15020381
APA StyleMoseki, O., Kangueehi, G., Chiteculo, V., Zink, M., & Wanga, M. A. (2025). Agro-Physiological and Morphological Responses of Pearl Millet to Varying Water Regimes in Semi-Arid Conditions of Namibia. Agronomy, 15(2), 381. https://doi.org/10.3390/agronomy15020381