
Pesticides Toxicity, Removal and Detoxification in Plants: A Review



Abstract
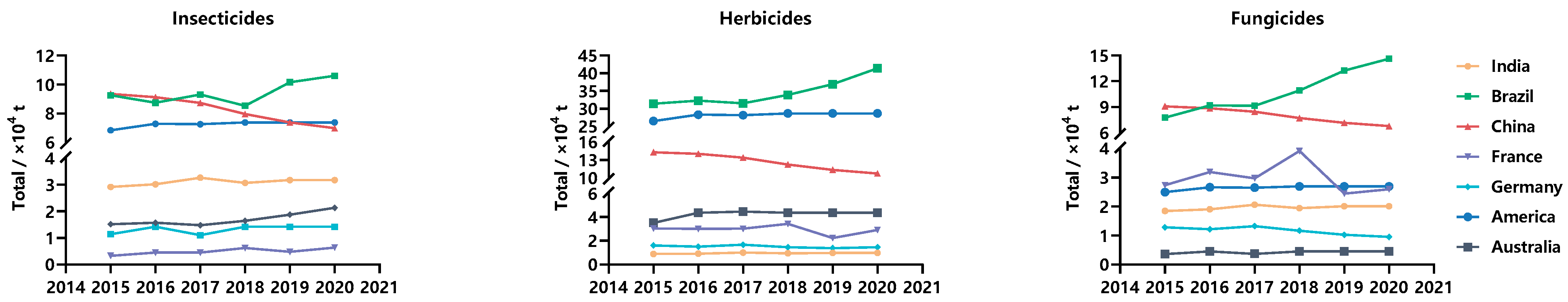
1. Introduction
2. The Physiological and Biochemical Effects of Pesticides on Plants
2.1. Pesticides Affect Plant Photosynthesis
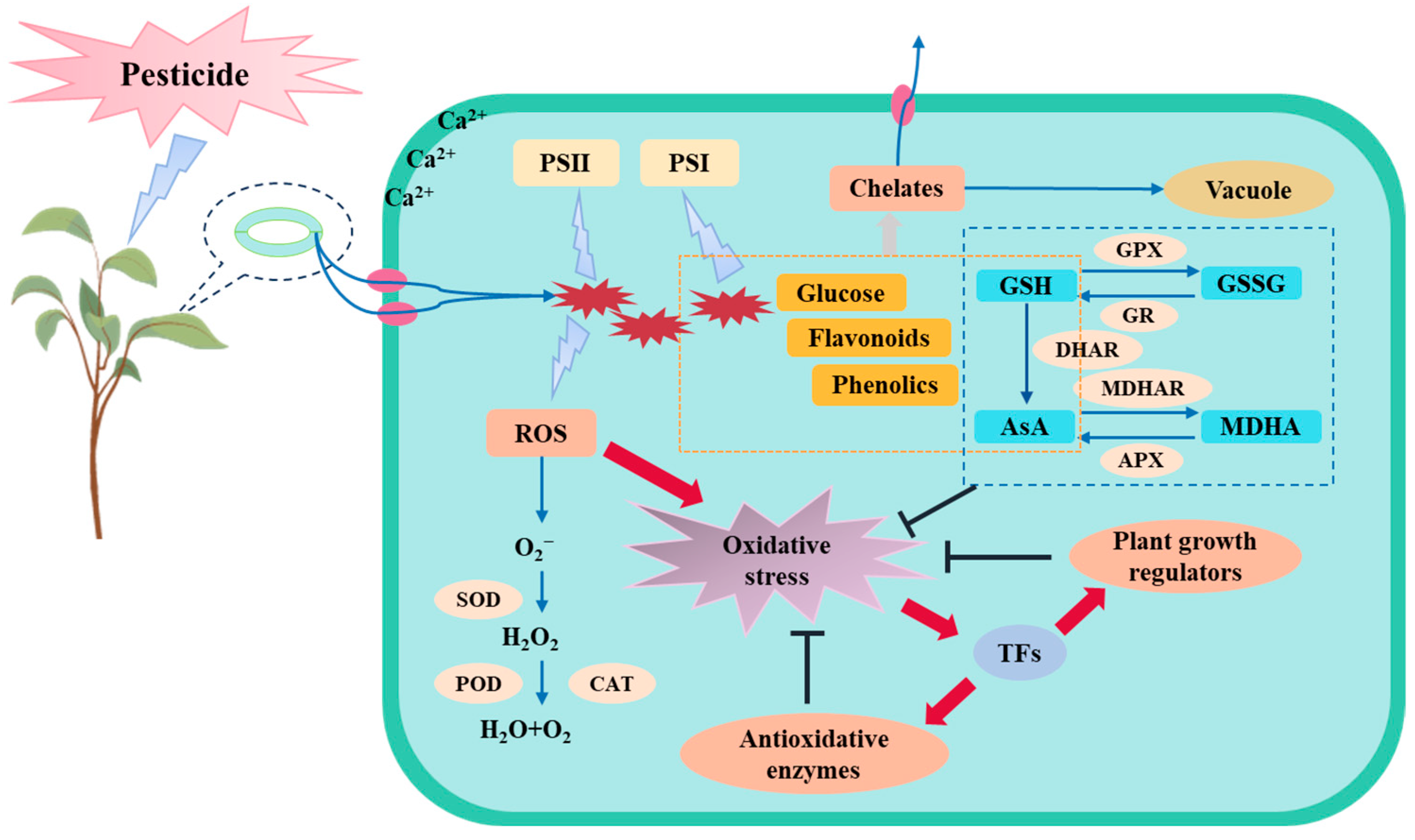
2.2. Pesticides Affect the Plant Active Oxygen Scavenging System
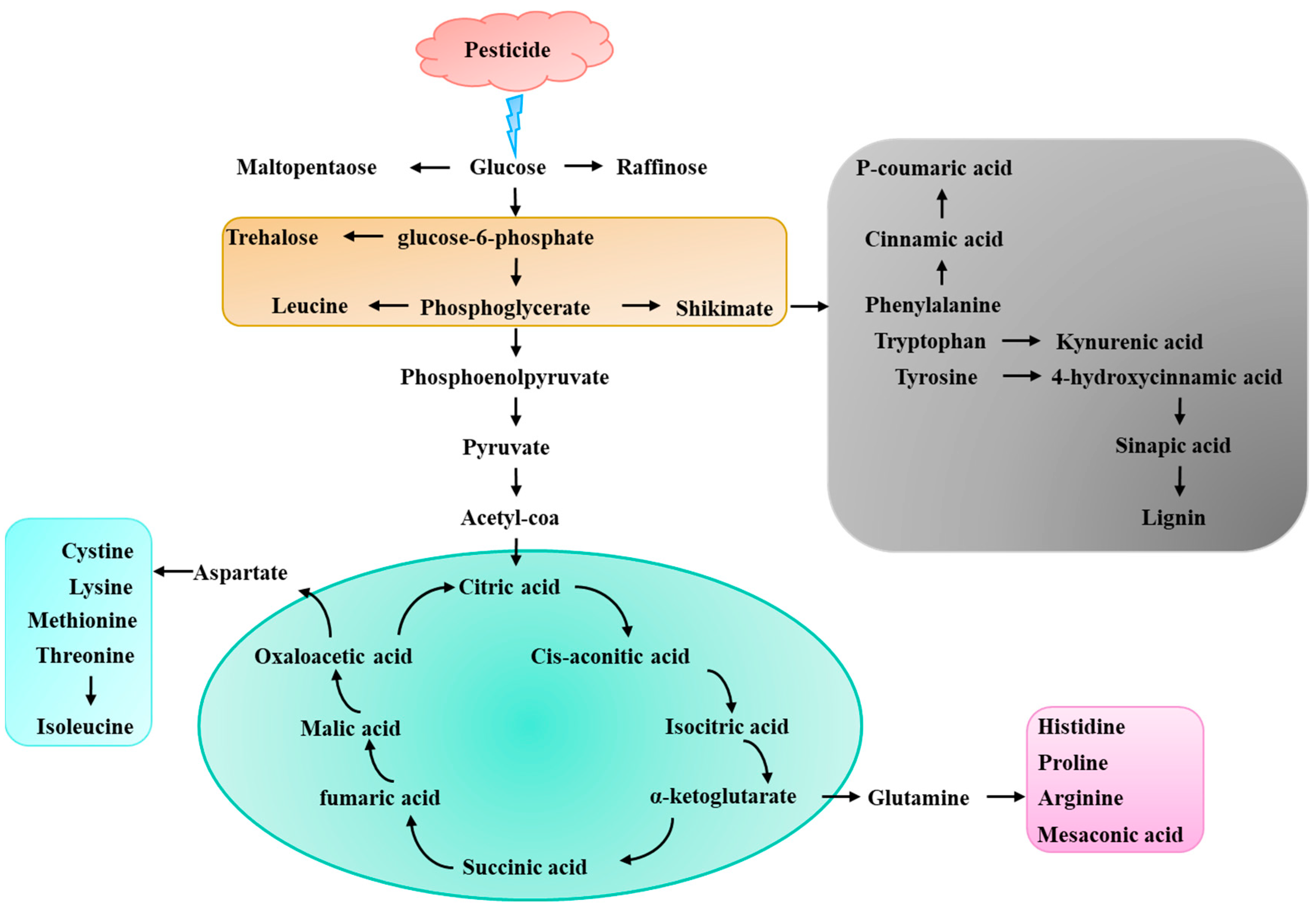
2.3. Pesticides Affect Plant Growth and Metabolites
3. The Degradation of Pesticides Outside of the Plant
3.1. Non-Biological Degradation
3.2. Biological Degradation
4. Pesticides Degradation in Plants
4.1. A Brief Description of Pesticides Detoxification
4.2. Main Factors in the Three-Step Detoxification Process
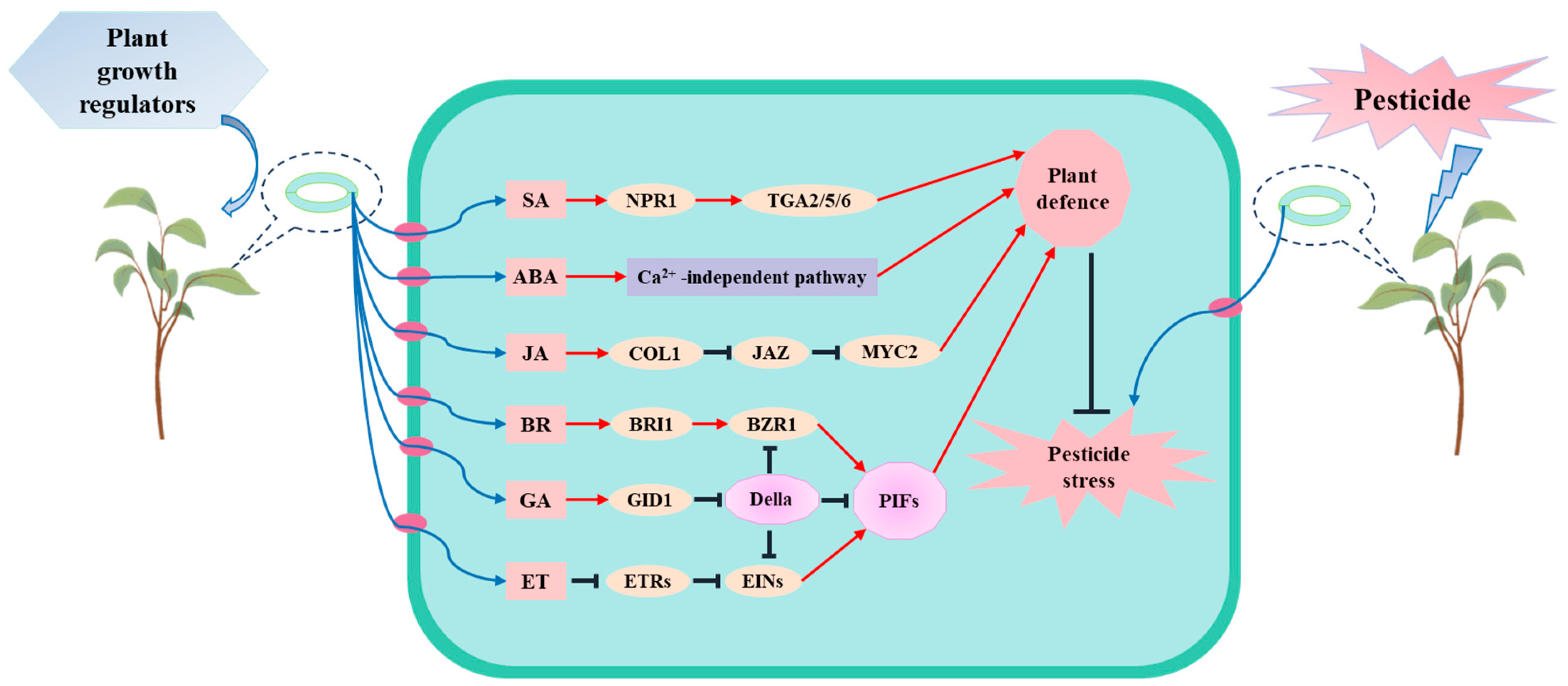
5. Exogenous Plant Growth Regulators Alleviate Pesticide Stress and Promote the Degradation of Pesticides by Plants
5.1. Pesticides Influence the Synthesis and Metabolism of Plant Signaling Molecules by Affecting Genes, Thereby Altering Hormone Levels
5.2. Exogenous Plant Growth Regulators Have Been Found to Alleviate the Damage That Pesticides Cause to the Photosystem
5.3. Exogenous Plant Growth Regulators Are Known to Enhance Enzymatic Activity, Thereby Reducing Cellular Damage Caused by Pesticides
5.4. Exogenous Plant Growth Regulators Reduce Pesticide Residues and Exhibit a Close Correlation with Secondary Metabolites
6. Conclusions and Prospect
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| APX | ascorbic acid peroxidase |
| AsA | ascorbic acid |
| CAT | catalase |
| DNA | deoxyribo nucleic acid |
| DZ | dichlorophen |
| EBL | brassinolide |
| EC | ethyl carbamate |
| ET | ethylene |
| FAO | Food and Agriculture Organization |
| FE | fenpropathrin |
| FEN | fenvalerate |
| Fm | fluorescence maximum |
| Fv | variable fluorescence |
| GGT | gamma-glutamyl transpeptidase |
| GPX | glutathione peroxidase |
| GR | glutathione reductase |
| GRXS | glutaredoxin |
| Gs | stomatal conductance |
| GSH | glutathione |
| GSSG | glutathione oxidized |
| GST | glutathione s-transferase |
| GUN | genomes uncoupled |
| H2O2 | hydrogen peroxide |
| IAA | indole acetic acid |
| ICS | isochromatic synthase |
| IMD | imidacloprid |
| IMI | imidacloprid |
| IPU | isoproturon |
| JA | jasmonic acid |
| KAT | potassium channel KAT1-like protein |
| kGy | kilogray |
| LCT | lambda-cyhalothrin |
| MAPK | mitogen-activated protein kinases |
| MBC | carbendazim |
| Md | malate dehydrogenase |
| MDA | malondialdehyde |
| MeJA | methyl jasmonate |
| MOF | magnetic nanocomposites of coated ferrites |
| MT | melatonin |
| MV | methyl viologen |
| NADPH | triphosphopyridine nucleotide |
| NF | norflurazon |
| NO | nitric oxide |
| O2 | oxygen |
| O2− | superoxide |
| -OH | hydroxyl radical |
| Os | oryza sativa |
| PMS | peroxymonosulfate |
| Pn | photosynthetic rate |
| POD | peroxidase |
| PPO | polyphenol oxidase |
| Pro | proline |
| PSII | photosystem II |
| ROS | reactive oxygen species |
| RUBP | ribulose-1,5-bisphosphate |
| SA | salicylic acid |
| SOD | superoxide dismutase |
| TDC | tryptophan decarboxylase |
| TGA | transcription factor |
| TOC | total organic carbon |
| VC | vitamin C |
| VE | vitamin E |
References
- Chung, S.W. How effective are common household preparations on removing pesticide residues from fruit and vegetables? A review. J. Sci. Food Agric. 2018, 98, 2857–2870. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#home (accessed on 16 December 2023).
- Rashid, S.; Rashid, W.; Tulcan, R.X.S.; Huang, H. Use, exposure, and environmental impacts of pesticides in Pakistan: A critical review. Environ. Sci. Pollut. Res. Int. 2022, 29, 43675–43689. [Google Scholar] [CrossRef]
- Li, C.; Zhu, H.; Li, C.; Qian, H.; Yao, W.; Guo, Y. The present situation of pesticide residues in China and their removal and transformation during food processing. Food Chem. 2021, 354, 129552. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Shao, C.; He, R.; Shi, R. Evolution and Efficiency Assessment of Pesticide and Fertiliser Inputs to Cultivated Land in China. Int. J. Environ. Res. Public Health 2021, 18, 3771. [Google Scholar] [CrossRef]
- The International Code of Conduct on Pesticide Management. Available online: http://www.fao.org/fileadmin/templates/agphome/documents/Pests_Pesticides/Code/CODE_2014Sep_ENG.pdf (accessed on 4 October 2019).
- Sharifzadeh, M.S.; Abdollahzadeh, G.; Damalas, C.A.; Rezaei, R.; Ahmadyousefi, M. Determinants of pesticide safety behavior among Iranian rice farmers. Sci. Total Environ. 2019, 651, 2953–2960. [Google Scholar] [CrossRef] [PubMed]
- Repetto, R.; Baliga, S.S. Pesticides and immunosuppression: The risks to public health. Health Policy Plan. 1997, 12, 97–106. [Google Scholar] [CrossRef]
- Andreotti, G.; Hou, L.; Beane Freeman, L.E.; Mahajan, R.; Koutros, S.; Coble, J.; Lubin, J.; Blair, A.; Hoppin, J.A.; Alavanja, M. Body mass index, agricultural pesticide use, and cancer incidence in the Agricultural Health Study cohort. Cancer Causes Control. 2010, 21, 1759–1775. [Google Scholar] [CrossRef] [PubMed]
- Fioresi, V.S.; de Cássia Ribeiro Vieira, B.; de Campos, J.M.S.; da Silva Souza, T. Cytogenotoxic activity of the pesticides imidacloprid and iprodione on Allium cepa root meristem. Environ. Sci. Pollut. Res. Int. 2020, 27, 28066–28076. [Google Scholar] [CrossRef]
- Li, Y.; Long, L.; Ge, J.; Li, H.; Zhang, M.; Wan, Q.; Yu, X. Effect of Imidacloprid Uptake from Contaminated Soils on Vegetable Growth. J. Agric. Food Chem. 2019, 67, 7232–7242. [Google Scholar] [CrossRef]
- Li, D.; Zhou, C.; Wang, S.; Hu, Z.; Xie, J.; Pan, C.; Sun, R. Imidacloprid-induced stress affects the growth of pepper plants by disrupting rhizosphere-plant microbial and metabolite composition. Sci. Total Environ. 2023, 898, 165395. [Google Scholar] [CrossRef]
- Liu, T.; Li, T.; Zhang, L.; Li, H.; Liu, S.; Yang, S.; An, Q.; Pan, C.; Zou, N. Exogenous salicylic acid alleviates the accumulation of pesticides and mitigates pesticide-induced oxidative stress in cucumber plants (Cucumis sativus L.). Ecotoxicol. Environ. Saf. 2021, 208, 111654. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, L.; Liu, L.; Cao, X.; Sun, C.; Lin, X. Metabolic disturbance in lettuce (Lactuca sativa) plants triggered by imidacloprid and fenvalerate. Sci. Total Environ. 2022, 802, 149764. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Zhang, M.; Li, Y.; Feng, F.; Yu, X.; Nie, J. Metabolomic Analysis Reveals the Effect of Insecticide Chlorpyrifos on Rice Plant Metabolism. Metabolites 2022, 12, 1289. [Google Scholar] [CrossRef]
- Wang, L.; Qin, Z.; Li, X.; Yang, J.; Xin, M. Persistence behavior of chlorpyrifos and biological toxicity mechanism to cucumbers under greenhouse conditions. Ecotoxicol. Environ. Saf. 2022, 242, 113894. [Google Scholar] [CrossRef] [PubMed]
- Sobiecka, E.; Mroczkowska, M.; Olejnik, T.P. The Enzymatic Antioxidants Activities Changes in Water Plants Tissues Exposed to Chlorpyrifos Stress. Antioxidants 2022, 11, 2104. [Google Scholar] [CrossRef]
- Sobiecka, E.; Mroczkowska, M.; Olejnik, T.P. The Influence of Chlorpyrifos on the Nonenzymatic Antioxidants Content in Macrophytes Leaves. Antioxidants 2022, 11, 684. [Google Scholar] [CrossRef]
- Panfili, I.; Bartucca, M.L.; Marrollo, G.; Povero, G.; Del Buono, D. Application of a Plant Biostimulant to Improve Maize (Zea mays) Tolerance to Metolachlor. J. Agric. Food Chem. 2019, 67, 12164–12171. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, B.; Sørensen, P.B.; Bruus, M.; Bossi, R.; Dupont, Y.L.; Link, M.; Damgaard, C.F. Effects of glyphosate spray-drift on plant flowering. Environ. Pollut. 2021, 280, 116953. [Google Scholar] [CrossRef]
- Carpenter, D.J.; Mathiassen, S.K.; Boutin, C.; Strandberg, B.; Casey, C.S.; Damgaard, C. Effects of Herbicides on Flowering. Environ. Toxicol. Chem. 2020, 39, 1244–1256. [Google Scholar] [CrossRef]
- Sun, Z.; Dzakpasu, M.; Zhang, D.; Liu, G.; Wang, Z.; Qu, M.; Chen, R.; Wang, X.C.; Zheng, Y. Enantioselectivity and mechanisms of chiral herbicide biodegradation in hydroponic systems. Chemosphere 2022, 307, 135701. [Google Scholar] [CrossRef]
- Wang, X.D.; Zhang, C.Y.; Yuan, Y.; Hua, Y.F.; Asami, T.; Qin, Y.; Xiong, X.H.; Zhu, J.L.; Lu, Y.C. Molecular Responses and Degradation Mechanisms of the Herbicide Diuron in Rice Crops. J. Agric. Food Chem. 2022, 70, 14352–14366. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qin, Y.; Pu, J.; Hu, J.; Wen, Y. Phytotoxicity of the chiral herbicide dichlorprop: Cross-talk between nitric oxide, reactive oxygen species and phytohormones. Sci. Total Environ. 2021, 788, 147866. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Sun, S.; Zhao, N.; Yang, W.; Shi, Q.; Gong, B. COMT1 overexpression resulting in increased melatonin biosynthesis contributes to the alleviation of carbendazim phytotoxicity and residues in tomato plants. Environ. Pollut. 2019, 252, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Mo, X.; Wang, W.; Chen, X.; Lou, Y. The Commonly Used Bactericide Bismerthiazol Promotes Rice Defenses against Herbivores. Int. J. Mol. Sci. 2018, 19, 1271. [Google Scholar] [CrossRef] [PubMed]
- Jakl, M.; Kovač, I.; Ćavar Zeljković, S.; Jaklová Dytrtová, J. Triazole fungicides in soil affect the yield of fruit, green biomass, and phenolics production of Solanum lycopersicum L. Food Chem. 2021, 351, 129328. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, K.; De, S.; Banerjee, S.; Roy, S. Pesticide mediated oxidative stress induces genotoxicity and disrupts chromatin structure in fenugreek (Trigonella foenum-graecum L.) seedlings. J. Hazard. Mater. 2019, 369, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska-Trypuć, A.; Wiater, J. Protective effect of plant compounds in pesticides toxicity. J. Environ. Health Sci. Eng. 2022, 20, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Penconazole and calcium ameliorate drought stress in canola by upregulating the antioxidative enzymes. Funct. Plant Biol. 2020, 47, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Singh, R.; Bhardwaj, R.; Ahmad, P.; Kapoor, D. Plant growth regulators: A sustainable approach to combat pesticide toxicity. 3 Biotech 2020, 10, 466. [Google Scholar] [CrossRef] [PubMed]
- Sule, R.O.; Condon, L.; Gomes, A.V. A Common Feature of Pesticides: Oxidative Stress-The Role of Oxidative Stress in Pesticide-Induced Toxicity. Oxid. Med. Cell Longev. 2022, 2022, 5563759. [Google Scholar] [CrossRef]
- Ahuja, I.; de Vos, R.C.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of Stomatal Closure in Plants Exposed to Drought and Cold Stress. Adv. Exp. Med. Biol. 2018, 1081, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Thor, K.; Jiang, S.; Michard, E.; George, J.; Scherzer, S.; Huang, S.; Dindas, J.; Derbyshire, P.; Leitão, N.; DeFalco, T.A.; et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 2020, 585, 569–573. [Google Scholar] [CrossRef]
- McAinsh, M.R.; Gray, J.E.; Hetherington, A.M.; Leckie, C.P.; Ng, C. Ca2+ signalling in stomatal guard cells. Biochem. Soc. Trans. 2000, 28, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Song, S.; Zhang, H.; Li, Y.; Niu, L.; Zhang, J.; Wang, W. Signaling Transduction of ABA, ROS, and Ca2+ in Plant Stomatal Closure in Response to Drought. Int. J. Mol. Sci. 2022, 23, 14824. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Azeem, F.; Long, Y.; Boeglin, M.; Duby, G.; Mouline, K.; Hosy, E.; Vavasseur, A.; Chérel, I.; Simonneau, T.; et al. Non-autonomous stomatal control by pavement cell turgor via the K+ channel subunit AtKC1. Plant Cell 2022, 34, 2019–2037. [Google Scholar] [CrossRef]
- Liu, N.; Li, J.; Lv, J.; Yu, J.; Xie, J.; Wu, Y.; Tang, Z. Melatonin alleviates imidacloprid phytotoxicity to cucumber (Cucumis sativus L.) through modulating redox homeostasis in plants and promoting its metabolism by enhancing glutathione dependent detoxification. Ecotoxicol. Environ. Saf. 2021, 217, 112248. [Google Scholar] [CrossRef]
- Faize, M.; Burgos, L.; Faize, L.; Piqueras, A.; Nicolas, E.; Barba-Espin, G.; Clemente-Moreno, M.J.; Alcobendas, R.; Artlip, T.; Hernandez, J.A. Involvement of cytosolic ascorbate peroxidase and Cu/Zn-superoxide dismutase for improved tolerance against drought stress. J. Exp. Bot. 2011, 62, 2599–2613. [Google Scholar] [CrossRef]
- Ghorbani, A.; Ghasemi-Omran, V.O.; Chen, M. The Effect of Glycine Betaine on Nitrogen and Polyamine Metabolisms, Expression of Glycoside-Related Biosynthetic Enzymes, and K/Na Balance of Stevia under Salt Stress. Plants 2023, 12, 1628. [Google Scholar] [CrossRef]
- Sun, M.; Xu, W.; Zhang, W.; Guang, C.; Mu, W. Microbial elimination of carbamate pesticides: Specific strains and promising enzymes. Appl. Microbiol. Biotechnol. 2022, 106, 5973–5986. [Google Scholar] [CrossRef]
- Benina, M.; Ribeiro, D.M.; Gechev, T.S.; Mueller-Roeber, B.; Schippers, J.H. A cell type-specific view on the translation of mRNAs from ROS-responsive genes upon paraquat treatment of Arabidopsis thaliana leaves. Plant Cell Environ. 2015, 38, 349–363. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Mellidou, I.; Koukounaras, A.; Kostas, S.; Patelou, E.; Kanellis, A.K. Regulation of Vitamin C Accumulation for Improved Tomato Fruit Quality and Alleviation of Abiotic Stress. Genes 2021, 12, 694. [Google Scholar] [CrossRef]
- Broad, R.C.; Bonneau, J.P.; Hellens, R.P.; Johnson, A.A.T. Manipulation of Ascorbate Biosynthetic, Recycling, and Regulatory Pathways for Improved Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 1790. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, J.P. Does plant-Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, L.; Leng, R.; Zhang, J.; Zhou, Y.; Zhang, Y.; Yang, S.; He, K.; Huang, B. Mechanism study of the beneficial effect of sodium selenite on metabolic disorders in imidacloprid-treated garlic plants. Ecotoxicol. Environ. Saf. 2020, 200, 110736. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, W.; Li, J.; Pang, S.; Mishra, S.; Bhatt, P.; Zeng, D.; Chen, S. Emerging Technologies for Degradation of Dichlorvos: A Review. Int. J. Environ. Res. Public Health 2021, 18, 5789. [Google Scholar] [CrossRef]
- Fahey, J.E.; Nelson, P.E.; Ballee, D.L. Removal of gardona from fruit by commercial preparative methods. J. Agric. Food Chem. 1970, 18, 866–868. [Google Scholar] [CrossRef]
- Zhang, Y.; Hou, Y.; Chen, F.; Xiao, Z.; Zhang, J.; Hu, X. The degradation of chlorpyrifos and diazinon in aqueous solution by ultrasonic irradiation: Effect of parameters and degradation pathway. Chemosphere 2011, 82, 1109–1115. [Google Scholar] [CrossRef]
- Debabrata, P.; Sivakumar, M. Sonochemical degradation of endocrine-disrupting organochlorine pesticide Dicofol: Investigations on the transformation pathways of dechlorination and the influencing operating parameters. Chemosphere 2018, 204, 101–108. [Google Scholar] [CrossRef]
- Kovács, K.; He, S.; Mile, V.; Csay, T.; Takács, E.; Wojnárovits, L. Ionizing radiation induced degradation of diuron in dilute aqueous solution. Chem. Cent. J. 2015, 9, 21. [Google Scholar] [CrossRef]
- Tan, W.; Li, G.; Bai, M.; Yang, M.; Peng, J.; Barrow, C.J.; Yang, W.; Wang, H. The study of adsorption mechanism of mixed pesticides prometryne-acetochlor in the soil-water system. Int. Biodeterior. Biodegrad. 2015, 102, 281–285. [Google Scholar] [CrossRef]
- Yue, L.; Ge, C.; Feng, D.; Yu, H.; Deng, H.; Fu, B. Adsorption-desorption behavior of atrazine on agricultural soils in China. J. Environ. Sci. 2017, 57, 180–189. [Google Scholar] [CrossRef]
- Bruckmann, F.S.; Schnorr, C.; Oviedo, L.R.; Knani, S.; Silva, L.F.O.; Silva, W.L.; Dotto, G.L.; Bohn Rhoden, C.R. Adsorption and Photocatalytic Degradation of Pesticides into Nanocomposites: A Review. Molecules 2022, 27, 6261. [Google Scholar] [CrossRef]
- Vitola, G.; Mazzei, R.; Giorno, L. Biohybrid membranes for organophosphate pesticides degradation: Hyperactivation of immobilized phosphotriesterase by surfactants. Environ. Technol. Innov. 2023, 30, 103053. [Google Scholar] [CrossRef]
- Lazarou, S.; Antonoglou, O.; Mourdikoudis, S.; Serra, M.; Sofer, Z.; Dendrinou-Samara, C. Magnetic Nanocomposites of Coated Ferrites/MOF as Pesticide Adsorbents. Molecules 2022, 28, 39. [Google Scholar] [CrossRef]
- Rani, M.; Shanker, U. Degradation of traditional and new emerging pesticides in water by nanomaterials: Recent trends and future recommendations. Int. J. Environ. Sci. Technol. 2017, 15, 1347–1380. [Google Scholar] [CrossRef]
- Dubocq, F.; L’Yvonnet, P.; Chatzidimitriou, E.; Mahouche-Chergui, S.; Sarda, X.; Vial, G.; Duboisset, A.; Carbonnier, B.; Parinet, J. Towards comprehensive identification of pesticide degradation products following thermal processing below and above 120 °C: A review. Food Chem. 2023, 402, 134267. [Google Scholar] [CrossRef]
- Syafrudin, M.; Kristanti, R.A.; Yuniarto, A.; Hadibarata, T.; Rhee, J.; Al-Onazi, W.A.; Algarni, T.S.; Almarri, A.H.; Al-Mohaimeed, A.M. Pesticides in Drinking Water-A Review. Int. J. Environ. Res. Public Health 2021, 18, 468. [Google Scholar] [CrossRef]
- He, J.; Evans, N.M.; Liu, H.; Zhu, Y.; Zhou, T.; Shao, S. UV treatment for degradation of chemical contaminants in food: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1857–1886. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, D.; Xu, C.; Zhong, J.; Chen, M.; Xu, S.; Cao, Y.; Zhao, Q.; Yang, M.; Ma, J. Synergistic oxidation—Filtration process analysis of catalytic CuFe2O4—Tailored ceramic membrane filtration via peroxymonosulfate activation for humic acid treatment. Water Res. 2020, 171, 115387. [Google Scholar] [CrossRef]
- Rezaei, S.S.; Dehghanifard, E.; Noorisepehr, M.; Ghadirinejad, K.; Kakavandi, B.; Esfahani, A.R. Efficient clean-up of waters contaminated with diazinon pesticide using photo-decomposition of peroxymonosulfate by ZnO decorated on a magnetic core/shell structure. J. Environ. Manag. 2019, 250, 109472. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Y. Study on the photocatalytic degradation of glyphosate by TiO(2) photocatalyst. Chemosphere 2007, 67, 1010–1017. [Google Scholar] [CrossRef]
- Marican, A.; Durán-Lara, E.F. A review on pesticide removal through different processes. Environ. Sci. Pollut. Res. Int. 2018, 25, 2051–2064. [Google Scholar] [CrossRef]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-Degrading Microorganisms and Their Potential for the Bioremediation of Contaminated Soils: A Review. Front. Microbiol. 2016, 7, 1463. [Google Scholar] [CrossRef]
- Whangchai, K.; Van Hung, T.; Al-Rashed, S.; Narayanan, M.; Kandasamy, S.; Pugazhendhi, A. Biodegradation competence of Streptomyces toxytricini D2 isolated from leaves surface of the hybrid cotton crop against β cypermethrin. Chemosphere 2021, 276, 130152. [Google Scholar] [CrossRef]
- Chen, C.; Yu, G.; Guo, Z.; Yang, Q.; Su, W.; Xie, Q.; Yang, G.; Ren, Y.; Li, H. Expression, Characterization, Fermentation, Immobilization, and Application of a Novel Esterase Est804 from Metagenomic Library in Pesticide Degradation. Front. Microbiol. 2022, 13, 922506. [Google Scholar] [CrossRef]
- Pang, S.; Lin, Z.; Zhang, Y.; Zhang, W.; Alansary, N.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the Toxicity and Degradation Mechanisms of Imidacloprid Via Physicochemical and Microbial Approaches. Toxics 2020, 8, 65. [Google Scholar] [CrossRef]
- Zheng, H.; Meng, K.; Liu, J.; Lin, Z.; Peng, Q.; Xie, G.; Wu, P.; Elsheery, N.I. Identification and expression of bifunctional acid urea-degrading enzyme/urethanase from Enterobacter sp. R-SYB082 and its application in degradation of ethyl carbamate in Chinese rice wine (Huangjiu). J. Sci. Food Agric. 2022, 102, 4599–4608. [Google Scholar] [CrossRef]
- Singh, T.; Singh, D.K. Rhizospheric Microbacterium sp. P27 Showing Potential of Lindane Degradation and Plant Growth Promoting Traits. Curr. Microbiol. 2019, 76, 888–895. [Google Scholar]
- Ellegaard-Jensen, L.; Aamand, J.; Kragelund, B.B.; Johnsen, A.H.; Rosendahl, S. Strains of the soil fungus Mortierella show different degradation potentials for the phenylurea herbicide diuron. Biodegradation 2013, 24, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Zhu, D.; Zhang, S.S.; Guan, J.G. Construction and application of genetic engineering bacteria in contaminated environment bioremediation. Adv. Mat. Res. 2014, 955–959, 1935–1938. [Google Scholar] [CrossRef]
- Tiwari, N.; Santhiya, D.; Sharma, J.G. Microbial remediation of micro-nano plastics: Current knowledge and future trends. Environ. Pollut. 2020, 265, 115044. [Google Scholar] [CrossRef]
- Sarker, A.; Nandi, R.; Kim, J.E.; Islam, T. Remediation of chemical pesticides from contaminated sites through potential microorganisms and their functional enzymes: Prospects and challenges. Environ. Technol. Innov. 2021, 23, 101777. [Google Scholar] [CrossRef]
- Cherian, S.; Oliveira, M.M. Transgenic plants in phytoremediation: Recent advances and new possibilities. Environ. Sci. Technol. 2005, 39, 9377–9390. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Sun, Q.; Li, J.; Ahammed, G.J.; Yu, J.; Fang, H.; Xia, X. Glutaredoxin S25 and its interacting TGACG motif-binding factor TGA2 mediate brassinosteroid-induced chlorothalonil metabolism in tomato plants. Environ. Pollut. 2019, 255, 113256. [Google Scholar] [CrossRef]
- Mansoor, S.; Ali Wani, O.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive Oxygen Species in Plants: From Source to Sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S. Metabolism-based herbicide resistance and cross-resistance in crop weeds: A threat to herbicide sustainability and global crop production. Plant Physiol. 2014, 166, 1106–1118. [Google Scholar] [CrossRef]
- Chen, S.; Yan, Y.; Wang, Y.; Wu, M.; Mao, Q.; Chen, Y.; Ren, J.; Liu, A.; Lin, X.; Ahammed, G.J. Trichoderma asperellum reduces phoxim residue in roots by promoting plant detoxification potential in Solanum lycopersicum L. Environ. Pollut. 2020, 259, 113893. [Google Scholar] [CrossRef]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. BioEssays 2006, 28, 1091–1101. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef]
- Geu-Flores, F.; Møldrup, M.E.; Böttcher, C.; Olsen, C.E.; Scheel, D.; Halkier, B.A. Cytosolic γ-glutamyl peptidases process glutathione conjugates in the biosynthesis of glucosinolates and camalexin in Arabidopsis. Plant Cell 2011, 23, 2456–2469. [Google Scholar] [CrossRef]
- Su, X.N.; Zhang, J.J.; Liu, J.T.; Zhang, N.; Ma, L.Y.; Lu, F.F.; Chen, Z.J.; Shi, Z.; Si, W.J.; Liu, C.; et al. Biodegrading Two Pesticide Residues in Paddy Plants and the Environment by a Genetically Engineered Approach. J. Agric. Food Chem. 2019, 67, 4947–4957. [Google Scholar] [CrossRef] [PubMed]
- Dimaano, N.G.; Iwakami, S. Cytochrome P450-mediated herbicide metabolism in plants: Current understanding and prospects. Pest. Manag. Sci. 2021, 77, 22–32. [Google Scholar] [CrossRef]
- Pan, G.; Zhang, X.; Liu, K.; Zhang, J.; Wu, X.; Zhu, J.; Tu, J. Map-based cloning of a novel rice cytochrome P450 gene CYP81A6 that confers resistance to two different classes of herbicides. Plant Mol. Biol. 2006, 61, 933–943. [Google Scholar] [CrossRef]
- Li, J.; Yu, H.; Zhang, F.; Lin, C.; Gao, J.; Fang, J.; Ding, X.; Shen, Z.; Xu, X. A built-in strategy to mitigate transgene spreading from genetically modified corn. PLoS ONE 2013, 8, e81645. [Google Scholar] [CrossRef]
- Saika, H.; Horita, J.; Taguchi-Shiobara, F.; Nonaka, S.; Nishizawa-Yokoi, A.; Iwakami, S.; Hori, K.; Matsumoto, T.; Tanaka, T.; Itoh, T.; et al. A novel rice cytochrome P450 gene, CYP72A31, confers tolerance to acetolactate synthase-inhibiting herbicides in rice and Arabidopsis. Plant Physiol. 2014, 166, 1232–1240. [Google Scholar] [CrossRef]
- Iwakami, S.; Endo, M.; Saika, H.; Okuno, J.; Nakamura, N.; Yokoyama, M.; Watanabe, H.; Toki, S.; Uchino, A.; Inamura, T. Cytochrome P450 CYP81A12 and CYP81A21 Are Associated with Resistance to Two Acetolactate Synthase Inhibitors in Echinochloa phyllopogon. Plant Physiol. 2014, 165, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulou, E.; Georgakis, N.; Nianiou-Obeidat, I.; Madesis, P.; Perperopoulou, F.; Pouliou, F.; Vasilopoulou, E.; Ioannou, E.; Ataya, F.S.; Labrou, N.E. Plant glutathione transferases in abiotic stress response and herbicide resistance. In Glutathione in Plant Growth, Development, and Stress Tolerance; Springer: Cham, Switzerland, 2017; pp. 215–233. [Google Scholar] [CrossRef]
- Iwakami, S.; Kamidate, Y.; Yamaguchi, T.; Ishizaka, M.; Endo, M.; Suda, H.; Nagai, K.; Sunohara, Y.; Toki, S.; Uchino, A.; et al. CYP81A P450s are involved in concomitant cross-resistance to acetolactate synthase and acetyl-CoA carboxylase herbicides in Echinochloa phyllopogon. New Phytol. 2019, 221, 2112–2122. [Google Scholar] [CrossRef]
- Garnik, E.Y.; Belkov, V.I.; Tarasenko, V.I.; Korzun, M.A.; Konstantinov, Y.M. Glutathione Reductase Gene Expression Depends on Chloroplast Signals in Arabidopsis thaliana. Biochemistry 2016, 81, 364–372. [Google Scholar] [CrossRef]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef]
- Berriri, S.; Garcia, A.V.; Frei dit Frey, N.; Rozhon, W.; Pateyron, S.; Leonhardt, N.; Montillet, J.L.; Leung, J.; Hirt, H.; Colcombet, J. Constitutively active mitogen-activated protein kinase versions reveal functions of Arabidopsis MPK4 in pathogen defense signaling. Plant Cell 2012, 24, 4281–4293. [Google Scholar] [CrossRef] [PubMed]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-transferase: A versatile protein family. 3 Biotech 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Yan, J.; Wu, X.; Li, T.; Fan, W.; Abbas, M.; Qin, M.; Li, R.; Liu, Z.; Liu, P. Effect and mechanism of nano-materials on plant resistance to cadmium toxicity: A review. Ecotoxicol. Environ. Saf. 2023, 266, 115576. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Hirosawa, Y.; Tada, A.; Matsuura, T.; Mori, I.C.; Ogura, Y.; Hayashi, T.; Uehara, S.; Ito-Inaba, Y.; Inaba, T. Salicylic Acid Acts Antagonistically to Plastid Retrograde Signaling by Promoting the Accumulation of Photosynthesis-associated Proteins in Arabidopsis. Plant Cell Physiol. 2021, 62, 1728–1744. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.Y.; Zhai, X.Y.; Qiao, Y.X.; Zhang, A.P.; Zhang, N.; Liu, J.; Yang, H. Identification of a novel function of a component in the jasmonate signaling pathway for intensive pesticide degradation in rice and environment through an epigenetic mechanism. Environ. Pollut. 2021, 268, 115802. [Google Scholar] [CrossRef]
- Lu, F.F.; Xu, J.Y.; Ma, L.Y.; Su, X.N.; Wang, X.Q.; Yang, H. Isoproturon-Induced Salicylic Acid Confers Arabidopsis Resistance to Isoproturon Phytotoxicity and Degradation in Plants. J. Agric. Food Chem. 2018, 66, 13073–13083. [Google Scholar] [CrossRef]
- Yu, G.B.; Chen, R.N.; Chen, Q.S.; Chen, F.Q.; Liu, H.L.; Ren, C.Y.; Zhang, Y.X.; Yang, F.J.; Wei, J.P. Jasmonic acid promotes glutathione assisted degradation of chlorothalonil during tomato growth. Ecotoxicol. Environ. Saf. 2022, 233, 113296. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.Y.; Zhang, S.H.; Zhang, J.J.; Zhang, A.P.; Li, N.; Wang, X.Q.; Yu, Q.Q.; Yang, H. Jasmonic Acids Facilitate the Degradation and Detoxification of Herbicide Isoproturon Residues in Wheat Crops (Triticum aestivum). Chem. Res. Toxicol. 2018, 31, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Ma, L.Y.; Chen, Z.J.; Wang, Y.; Gu, Y.; Yang, H. OsBR6ox, a member in the brassinosteroid synthetic pathway facilitates degradation of pesticides in rice through a specific DNA demethylation mechanism. Sci. Total Environ. 2022, 838, 156503. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wang, N.; Sun, S.; Geng, L.; Guo, N.; Liu, A.; Chen, S.; Ahammed, G.J. Reactive oxygen species signaling is involved in melatonin-induced reduction of chlorothalonil residue in tomato leaves. J. Hazard. Mater. 2023, 443, 130212. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Gao, T.; Liang, B.; Zhao, Q.; Ma, F.; Li, C. Effects of Exogenous Melatonin on Methyl Viologen-Mediated Oxidative Stress in Apple Leaf. Int. J. Mol. Sci. 2018, 19, 316. [Google Scholar] [CrossRef] [PubMed]
- Yüzbaşıoğlu, E.; Dalyan, E. Salicylic acid alleviates thiram toxicity by modulating antioxidant enzyme capacity and pesticide detoxification systems in the tomato (Solanum lycopersicum Mill.). Plant Physiol. Biochem. 2019, 135, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Mehrian, S.K.; Karimi, N.; Rahmani, F. 24-Epibrassinolide alleviates diazinon oxidative damage by escalating activities of antioxidant defense systems in maize plants. Sci. Rep. 2023, 13, 19631. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.Q.; Lu, F.F.; Ma, L.Y.; Yang, H.; Song, N.H. Residues of Reduced Herbicides Terbuthylazine, Ametryn, and Atrazine and Toxicology to Maize and the Environment through Salicylic Acid. ACS Omega 2021, 6, 27396–27404. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Li, D.; Zhou, C.; Wu, Y.; Miao, P.; Dong, Q.; Zhu, S.; Pan, C. Application of insecticides on peppermint (Mentha × piperita L.) induces lignin accumulation in leaves by consuming phenolic acids and thus potentially deteriorates quality. J. Plant Physiol. 2022, 279, 153836. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Baek, K.H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef]
- Dehghanian, Z.; Asgari Lajayer, B.; Biglari Quchan Atigh, Z.; Nayeri, S.; Ahmadabadi, M.; Taghipour, L.; Senapathi, V.; Astatkie, T.; Price, G.W. Micro (nano) plastics uptake, toxicity and detoxification in plants: Challenges and prospects. Ecotoxicol. Environ. Saf. 2023, 268, 115676. [Google Scholar] [CrossRef] [PubMed]
- Gaafar, R.M.; Osman, M.E.H.; Abo-Shady, A.M.; Almohisen, I.A.A.; Badawy, G.A.; El-Nagar, M.M.F.; Ismail, G.A. Role of Antioxidant Enzymes and Glutathione S-Transferase in Bromoxynil Herbicide Stress Tolerance in Wheat Plants. Plants 2022, 11, 2679. [Google Scholar] [CrossRef]
- Tian, M.; Zhang, Q.; Hua, X.; Tang, B.; Gao, B.; Wang, M. Systemic stereoselectivity study of flufiprole: Stereoselective bioactivity, acute toxicity and environmental fate. J. Hazard. Mater. 2016, 320, 487–494. [Google Scholar] [CrossRef]
- Li, R.; Pan, X.; Wang, Q.; Tao, Y.; Chen, Z.; Jiang, D.; Wu, C.; Dong, F.; Xu, J.; Liu, X.; et al. Development of S-Fluxametamide for Bioactivity Improvement and Risk Reduction: Systemic Evaluation of the Novel Insecticide Fluxametamide at the Enantiomeric Level. Environ. Sci. Technol. 2019, 53, 13657–13665. [Google Scholar] [CrossRef]
- Gao, B.; Zhang, Z.; Li, L.; Kaziem, A.E.; He, Z.; Yang, Q.; Qing, P.; Zhang, Q.; Wang, M. Stereoselective environmental behavior and biological effect of the chiral organophosphorus insecticide isofenphos-methyl. Sci. Total Environ. 2019, 648, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hansen, H.C.B.; Strobel, B.W.; Cedergreen, N. What is the aquatic toxicity of saponin-rich plant extracts used as biopesticides? Environ. Pollut. 2018, 236, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Diogo, B.S.; Antunes, S.C.; Rodrigues, S. Are biopesticides safe for the environment? Effects of pyrethrum extract on the non-target species Daphnia magna. Environ. Toxicol. Pharmacol. 2023, 99, 104114. [Google Scholar] [CrossRef] [PubMed]
- Cappa, F.; Baracchi, D.; Cervo, R. Biopesticides and insect pollinators: Detrimental effects, outdated guidelines, and future directions. Sci. Total Environ. 2022, 837, 155714. [Google Scholar] [CrossRef]
- Huang, Y.; Li, Z.; Luo, X.; Liu, D. Biopesticides extension and rice farmers’ adoption behavior: A survey from Rural Hubei Province, China. Environ. Sci. Pollut. Res. Int. 2022, 29, 51744–51757. [Google Scholar] [CrossRef]
- Awad, M.; Ibrahim, E.S.; Osman, E.I.; Elmenofy, W.H.; Mahmoud, A.W.M.; Atia, M.A.M.; Moustafa, M.A.M. Nano-insecticides against the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae): Toxicity, development, enzyme activity, and DNA mutagenicity. PLoS ONE 2022, 17, e0254285. [Google Scholar] [CrossRef]
- Gao, F.; Cui, B.; Wang, C.; Li, X.; Li, B.; Zhan, S.; Shen, Y.; Zhao, X.; Sun, C.; Wang, C.; et al. Nano-EMB-SP improves the solubility, foliar affinity, photostability and bioactivity of emamectin benzoate. Pest. Manag. Sci. 2022, 78, 3717–3724. [Google Scholar] [CrossRef] [PubMed]
- Laudani, F.; Campolo, O.; Caridi, R.; Latella, I.; Modafferi, A.; Palmeri, V.; Sorgonà, A.; Zoccali, P.; Giunti, G. Aphicidal Activity and Phytotoxicity of Citrus sinensis Essential-Oil-Based Nano-Insecticide. Insects 2022, 13, 1150. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Pesticides Types | Concentration | Main Results | Reference |
|---|---|---|---|---|
| Onion (Allium cepa L.) | Imidacloprid | 1.75 μg/mL, 17.5 μg/mL, 175 μg/mL, 1750 μg/mL | Root length↓ | [10] |
| Pok choy (Brassica rapa L.) | Imidacloprid | 10 mg/L | Proline↑, Sucrose↑, Raffinose↑, Disaccharides↑, Glutathione oxidized↑ | [11] |
| Wheat (Triticum aestivum L.) | Imidacloprid | 100 mg/kg, 200 mg/kg | Jasmonic acid in root and leaf↓, Indole acetic acid in root and leaf↓, Abscisic acid in root and leaf↑, Ferulic acid↓ | [12] |
| Cucumber (Cucumis sativus L.) | Imidacloprid | 2.75 mM | Fv/Fm↓, Ascorbic acid↓, Glutathione↓, Glutathione oxidized↑ | [13] |
| Lettuce (Lactuca sativa L.) | Imidacloprid, Fenvalerate | 10mg/L | Under IMI treatment, Iron↓, Arginine↓, Cysteine↓, Homoserine↓, 4-hydroxyisoleucine↓, Proline↓, Amino acid↓, Under FEN treatment, Iron↑, Flavonoid↓, Vitamin C↓ | [14] |
| Rice (Oryza sativa L.) | Chlorpyrifos | 5.0 mg/L | Peroxidase↑, Superoxide dismutase↑, Malondialdehyde↑, Protein↓, Chlorophyll↑ | [15] |
| Cucumber (Cucumis sativus L.) | Chlorpyrifos | 0.6 kg/ha | H+ efflux↑, K+ efflux↑, H2O2↑, O2−↑, Superoxide dismutase↑, Chlorophyll a↓, Chlorophyll b↓, Carotene↓ | [16] |
| Canadian waterweed (Elodea canadensis Michx.), needle spikerush (Eleocharis acicularis L.), water mint (Mentha aquatica L.) | Chlorpyrifos | 50 μg/dm3, 100 μg/dm3, 150 μg/dm3 | Glutathione peroxidase↑, Glutathione s-transferase↓, Chlorophyll a↓, Chlorophyll b↓, Carotene↓ | [17,18] |
| Plants | Pesticides Types | Concentration | Main Results | Reference |
|---|---|---|---|---|
| Maize (Zea mays L.) | Metolachlor | 0.5 mg/L, 1.0 mg/L, 2.0 mg/L, 4.0 mg/L, 8.0 mg/L | Malondialdehyde↑, Ascorbic acid peroxidase↑, Glutathione peroxidase↑, Catalase↑, Germination↓, Biomass production↓, Vigor index↓, Ethyl carbamate50↓ | [19] |
| Trifolium pratense L. Lotus corniculatus L. Trifolium repens L. Cichorium intybus L. | Glyphosate | 1440 g a.i./ha | cumulative number of Trifolium pratense L. flowers↓, cumulative number of Lotus corniculatus L. flowers↓, cumulative number of Trifolium repens L. flowers↑, | [20] |
| Cency (Centaurea cyanus L.), Silno (Silene noctiflora L.), Vioar (Viola arvensis Murray), Cerar (Cerastium arvense L.), Cirar (Cirsium arvense L.), Epimo (Epilobium montanum L.), Knaar (Knautia arvensis L.), Tarof (Taraxacum officinale F.H. Wigg.), Trfpr (Trifolium pratense L.) | bromoxynil, ioxynil Bromoxynil, Metsulfuron-Methyl, Clopyralid, Glyphosate | 280 g/ha 240 g a.i./ha 6 g a.i./ha 80 g/ha 1440 g/ha | Cumulative number of flowers↓, Flowering time↓ | [21] |
| Reed (Phragmites australis) | Metolachlor | 245 μg/L | Malondialdehyde↑, Reactive oxygen species↑, Chlorophyll↓then↑, Superoxide dismutase↑, Peroxidase↑, Catalase↑ | [22] |
| Rice (Oryza sativa L.) | Diuron | 0.125 mg/L, 0.25 mg/L, 0.5 mg/L, 1.0 mg/L, 2.0 mg/L | Elongation↓, Biomass↓, Chlorophyll↓, Malondialdehyde↑, Superoxide dismutase↑, Peroxidase↑, Glutathione reductase↑, Polyphenol oxidase↑, Ascorbic acid peroxidase↑, Catalase↑, Glutathione↑then↓, Jasmonic acid↑ | [23] |
| Arabidopsis (Arabidopsis thaliana) | Dichlorprop | 0.1 μM, 0.2 μM, 0.3 μM | Plant growth↓, H2O2↑, Jasmonic acid↑, Salicylic acid↑, Abscisic acid↓then↑ | [24] |
| Tomato (Solanum lycopersicum L.) | Carbendazim | 1 mM | Malondialdehyde↑, Fv/Fm↓, Superoxide dismutase↑, Catalase↑, Ascorbic acid peroxidase↑, Glutathione peroxidase↓, Ascorbic acid↓ | [25] |
| Rice (Oryza sativa L.) | Bismerthiazol | 10 mg/L, 20 mg/L, 50 mg/L | Jasmonic acid↑, OsAOS1↑, OsAOS2↑, H2O2↑, Volatile chemicals↑, | [26] |
| Tomato (Solanum lycopersicum L.) | Triazoles | 3.52 × 10−6 mol | Weight of thick stems↓, Weight of thin stems↓, Weight of leaves↓, Fruit weight↓, Quercetin↓, Naringin↓, Salicylic acid↓, Chlorogenic acid↑, Hesperidin↑ | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Lv, F.; Yang, J. Pesticides Toxicity, Removal and Detoxification in Plants: A Review. Agronomy 2024, 14, 1260. https://doi.org/10.3390/agronomy14061260
Zhang B, Lv F, Yang J. Pesticides Toxicity, Removal and Detoxification in Plants: A Review. Agronomy. 2024; 14(6):1260. https://doi.org/10.3390/agronomy14061260
Chicago/Turabian StyleZhang, Boyu, Fang Lv, and Jing Yang. 2024. "Pesticides Toxicity, Removal and Detoxification in Plants: A Review" Agronomy 14, no. 6: 1260. https://doi.org/10.3390/agronomy14061260
APA StyleZhang, B., Lv, F., & Yang, J. (2024). Pesticides Toxicity, Removal and Detoxification in Plants: A Review. Agronomy, 14(6), 1260. https://doi.org/10.3390/agronomy14061260