
Efficacy and Persistence of Entomopathogenic Fungi against Rhynchophorus ferrugineus on Date Palm: Host to Host Transmission


 ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing RPW
2.2. Culturing of Fungal Isolates
2.3. Screening of Fungal Isolates against RPW Larvae and Adults
2.4. Concentration Response Bioassay against Larvae and Adults of RPW
2.5. Bioassay against Eggs
2.6. Horizontal Transmission of Fungal Isolates among Adults and Effectiveness of Sublethal Concentrations on Reproductive Stages of RPW
2.7. Semi-Field Trials for RPW Survival
2.8. Fungal Persistence over Time and Its Effectiveness against Larvae of RPW
2.9. Statistical Analysis
3. Results
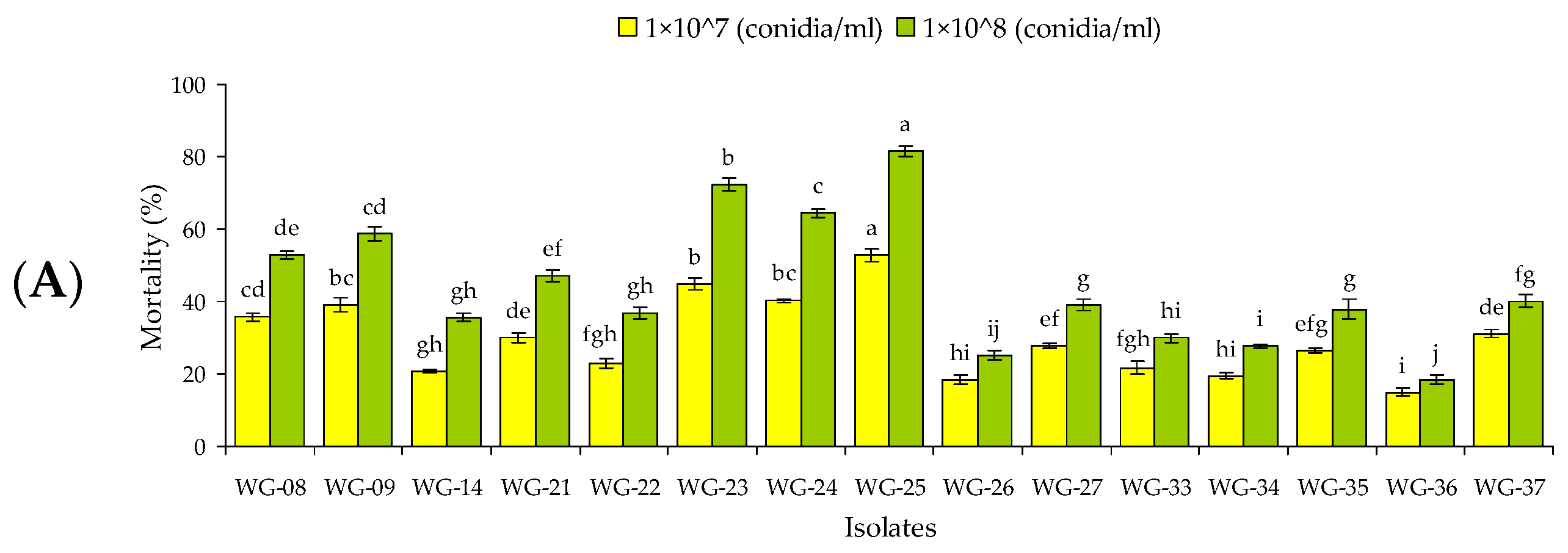
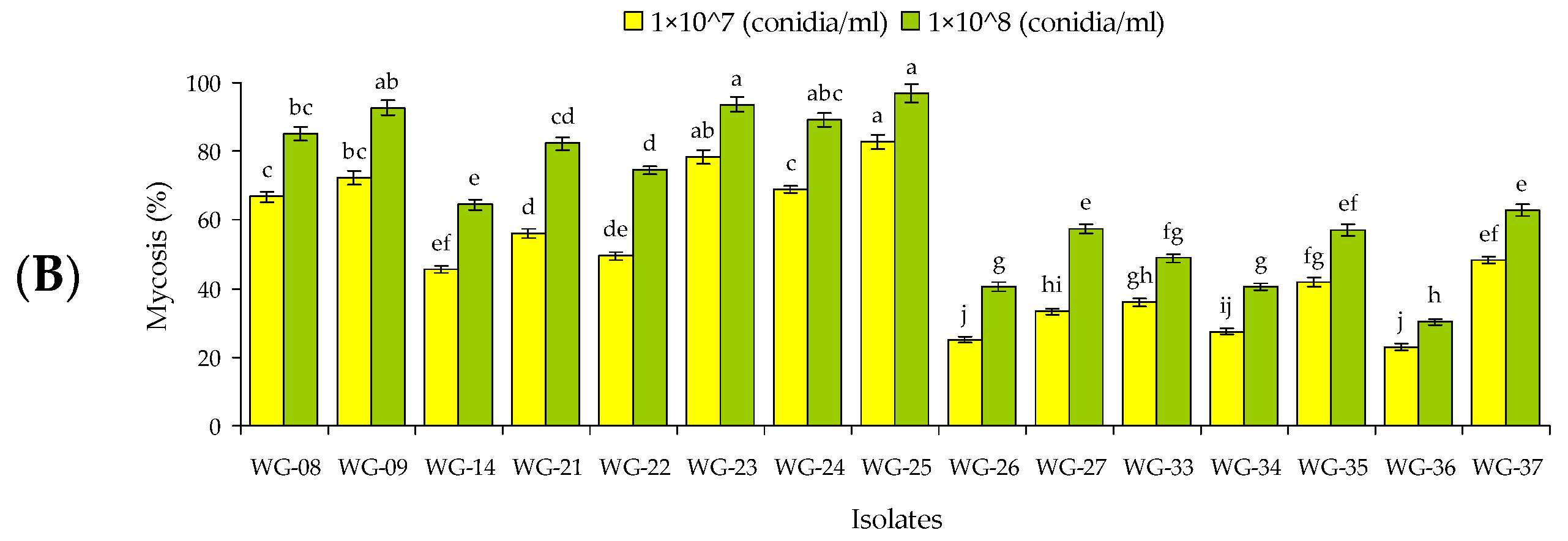
3.1. Screening of Fungal Isolates against Larvae and Adults
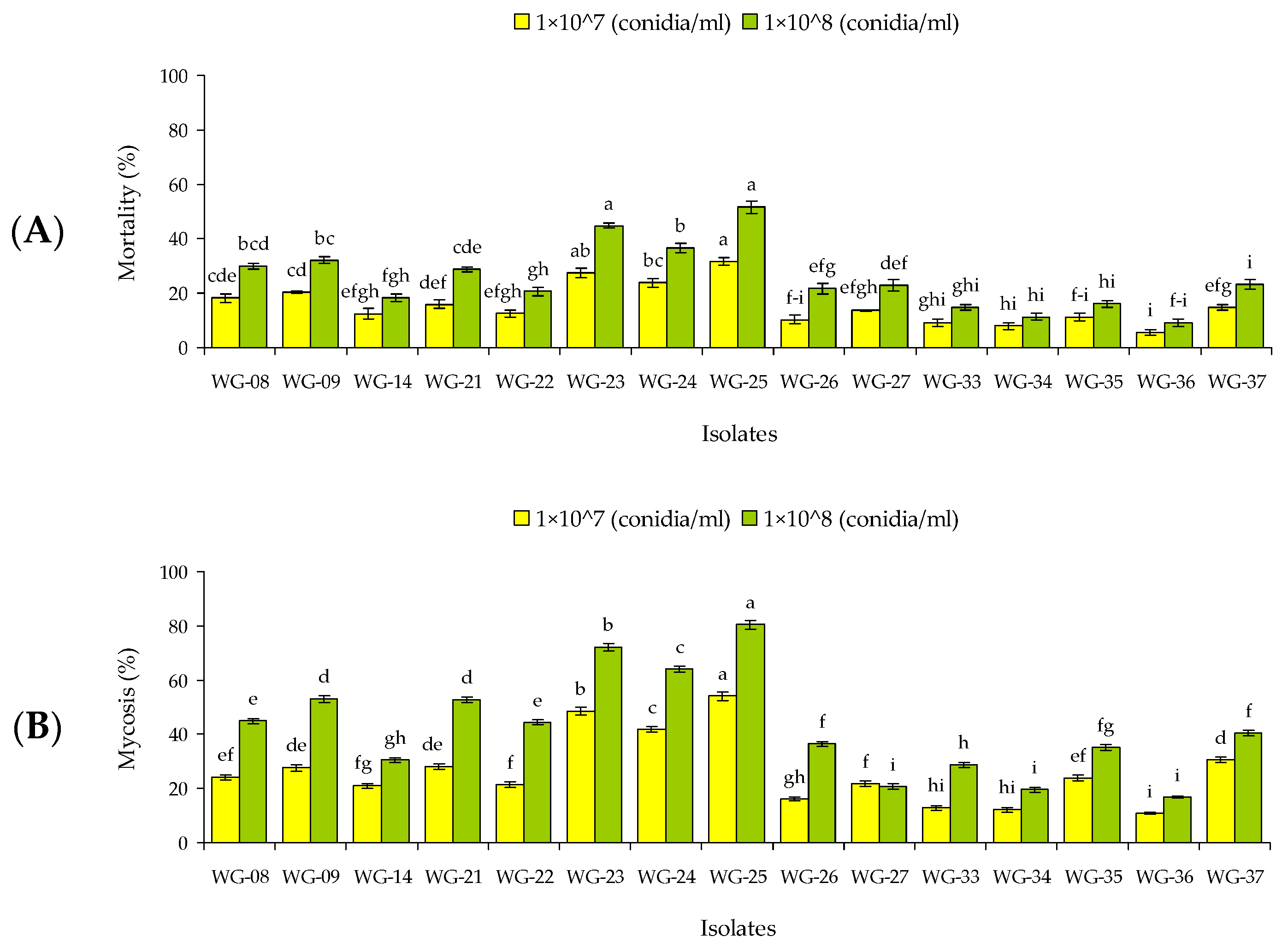
3.2. Concentration Response Bioassay against RPW Larvae and Adults
3.3. Lethal Concentration and Lethal Time
3.4. Bioassay against Eggs
3.5. Horizontal Transmission of Fungal Isolates among Adults and Effectiveness of Sublethal Concentrations on Reproductive Stages of RPW
3.6. Semi-Field Trials for RPW Survival
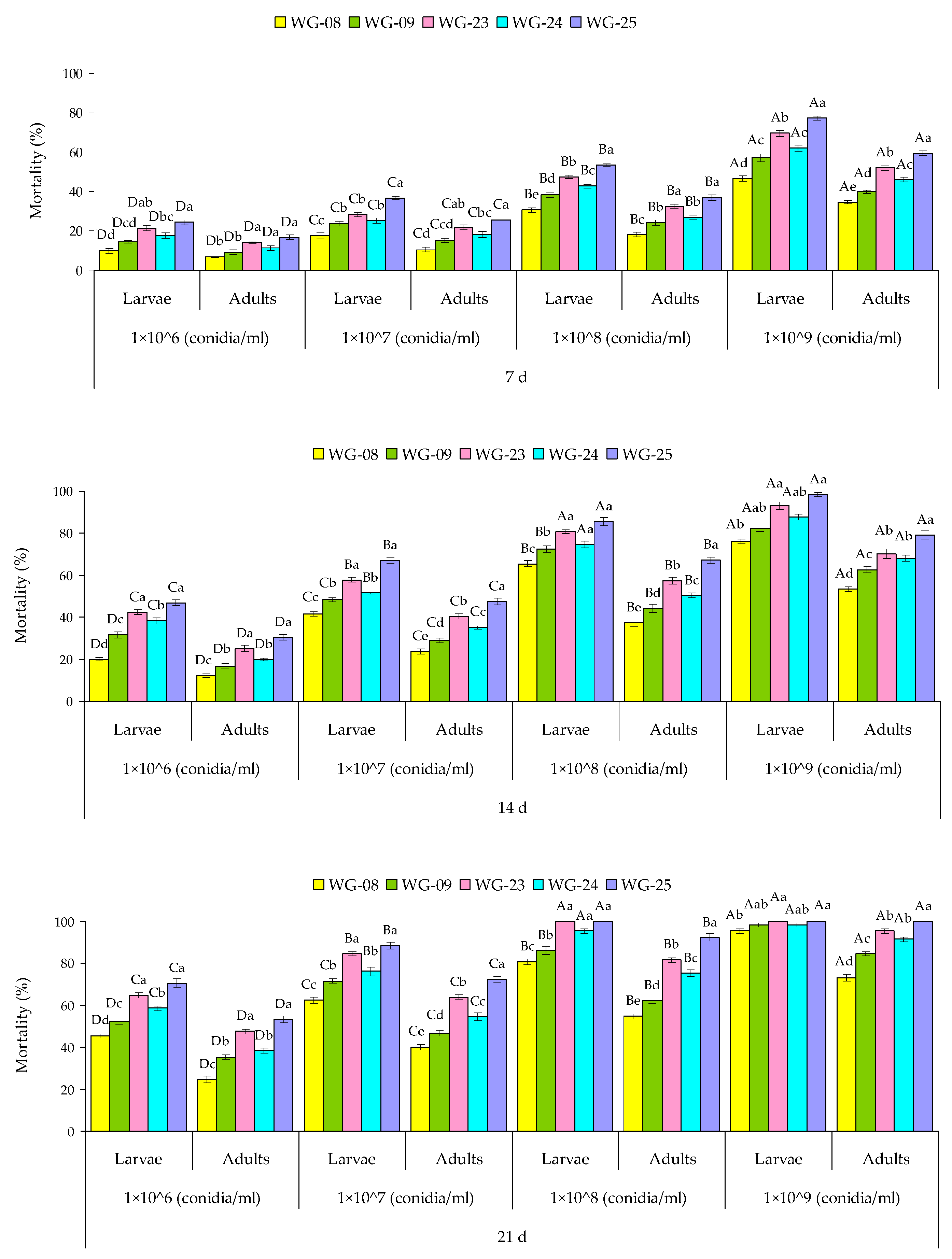
3.7. Fungal Persistence over Time and Its Effectiveness against RPW Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wakil, W.; Faleiro, J.R.; Miller, T.A.; Bedford, G.O.; Krueger, R.R. Date palm production and pest management challenges. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges, Sustainability in Plant and Crop Protection; Wakil, W., Faleiro, J.R., Miller, T.A., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–11. [Google Scholar]
- Tagliavia, M.; Messina, E.; Manachini, B.; Cappello, S.; Quatrini, P. The gut microbiota of larvae of Rhynchophorus ferrugineus Oliver (Coleoptera: Curculionidae). BMC Microbiol. 2014, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Fiaboe, K.K.M.; Peterson, A.T.; Kairo, M.T.K.; Roda, A.L. Predicting the potential worldwide distribution of the red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) using ecological niche modeling. Fla. Entomol. 2012, 95, 659–673. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). Rhynchophorus ferrugineus . EPPO Global Data Base. Available online: https://gd.eppo.int/taxon/RHYCFE/distribution (accessed on 25 February 2024).
- Manzoor, M.; Yang, L.; Wu, S.; El-Shafie, H.; Haider, M.S.; Ahmad, J.N. Feeding preference of Rhynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae) on different date palm cultivars and host biochemical responses to its infestation. Bull. Entomol. Res. 2022, 112, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.T. Red palm weevil, Rhynchophorus ferrugineus, a significant threat to date palm tree, global invasions, consequences, and management techniques. J. Plant Dis. Prot. 2024, 131, 9–26. [Google Scholar] [CrossRef]
- Abdel-Banat, B.M.A.; El-Shafie, H.A.F. Management of the red palm weevil in date palm plantations in Al-Ahsa oasis of Saudi Arabia. In Plant Health Cases; CABI Digital Library: Wallingford, UK, 2023; Volume 23, pp. 1–11. [Google Scholar]
- Dembilio, Ó.; Jacas, J.A.; Llácer, E. Are the palms Washingtonia filifera and Chamaerops humilis suitable hosts for the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). J. Appl. Entomol. 2009, 133, 565–567. [Google Scholar] [CrossRef]
- Fetoh, B.E. Latent effects of gamma radiation on certain biological aspects of the red palm weevil (Rhynchophorus ferrugineus Olivier) as a new control technology. J. Agric. Technol. 2011, 7, 1169–1175. [Google Scholar]
- Pu, Y.C.; Xiang, H.J.; Liang, X.Y.; Wang, Y.; Hou, Y.M.; Fu, L.; Wang, R. External immune inhibitory efficiency of external secretions and their metabolic profiling in red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Front. Physiol. 2020, 10, 1624. [Google Scholar] [CrossRef]
- Pinhas, J.; Soroker, V.; Hetzroni, A.; Mizrach, A.; Teicher, M.; Goldberger, J. Automatic acoustic detection of the red palm weevil. Comput. Electron. Agric. 2008, 63, 131–139. [Google Scholar] [CrossRef]
- El-Shafie, H.A.F.; Faleiro, J.R. Red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae): Global invasion, current management options, challenges and future prospects. In Invasive Species-Introduction Pathways, Economic Impact, and Possible Management Options; El-Shafie, H.A.F., Ed.; IntechOpen: London, UK, 2020; pp. 1–30. [Google Scholar]
- Boulila, W.; Alzahem, A.; Koubaa, A.; Benjdira, B.; Ammar, A. Early detection of red palm weevil infestations using deep learning classification of acoustic signals. Comput. Electr. Agric. 2023, 212, 108154. [Google Scholar] [CrossRef]
- Faleiro, J.R. A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. Int. J. Trop. Insect Sci. 2006, 26, 135–154. [Google Scholar]
- Al-Ballaa, S.R.; Faleiro, J.R. Studies on curative treatment of red palm weevil, Rhynchophorus ferrugineus Olivier infested date palms based on an innovative fumigation technique. Arab J. Plant Prot. 2019, 37, 119–123. [Google Scholar] [CrossRef]
- Jalinas, J.; Güerri Agulló, B.; Dosunmu, O.G.; Haseeb, M.; Lopez Llorca, L.V.; Mankin, R.W. Acoustic signal applications in detection and management of Rhynchophorus spp. In fruit-crops and ornamental palms. Fla. Entomol. 2019, 102, 475–479. [Google Scholar] [CrossRef]
- Al-Ballaa, S.R. Fumigant action of commonly used insecticides as a curative treatment of red palm weevil Rhynchophorus ferrugineus (Olivier) in infested date palms. Arab J. Plant Prot. 2020, 38, 333–338. [Google Scholar]
- Rehman, G.; Mammon-ur-Rashid, M. Evaluation of entomopathogenic nematodes against red palm weevil, Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Insects 2022, 13, 733. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Latif, A.O.; Genbi, Y.M.O.; Adel, M.M. Enzyme inhibitory potency of nano formulation of some plant oils on the red palm weevil Rhynchophorus ferrugineus Olivier. Nat. Prod. Res. 2024. [Google Scholar] [CrossRef] [PubMed]
- Sabbahi, R.; Hock, V. Entomopathogenic fungi against the red palm weevil: Lab and field evidence. Crop. Prot. 2024, 177, 106566. [Google Scholar] [CrossRef]
- Zhu, H.; Qin, W.Q.; Huang, S.C.; Yan, W.; Sun, X.D. Isolation and identification of an entomopathogenic fungus strain of Rhynchophorus ferrugineus Oliver. Acta Phytophylacica Sin. 2010, 37, 336–340. [Google Scholar]
- Francardi, V.; Benvenuti, C.; Roversi, P.F.; Rumine, P.; Barzanti, G. Entomopathogenicity of Beauveria bassiana (Bals.) Vuill. and Metarhizium anisopliae (Metsch.) Sorokin isolated from different sources in the control of Rhynchophorus ferrugineus (Olivier) (Coleoptera Curculionidae). Redia 2012, 95, 49–55. [Google Scholar]
- Wakil, W.; Faleiro, J.R.; Miller, T.A. Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges; Springer International Publishing: Cham, Switzerland, 2015; p. 429. [Google Scholar]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol. Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Roy, H.E.; Vega, F.E.; Chandler, D.; Goettel, M.S.; Pell, J.K.; Wajnberg, E. The Ecology of Fungal Entomopathogens; Springer: Dordrecht, The Netherlands, 2010; p. 198. [Google Scholar]
- Tahir, M.; Wakil, W.; Ali, A.; Sahi, S.T. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae isolates against larvae of the polyphagous pest Helicoverpa armigera. Entomol. Gen. 2019, 38, 225–242. [Google Scholar] [CrossRef]
- Yasin, M.; Wakil, W.; Ghazanfar, M.U.; Qayyum, M.A.; Tahir, M.; Bedford, G.O. Virulence of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against red palm weevil, Rhynchophorus ferrugineus (Olivier). Entomol. Res. 2019, 49, 3–12. [Google Scholar] [CrossRef]
- Wakil, W.; Kavallieratos, N.G.; Ghazanfar, M.U.; Usman, M.; Habib, A.; El-Shafie, H.A.F. Efficacy of different entomopathogenic fungal isolates against four key stored-grain beetle species. J. Stored Prod. Res. 2021, 93, 101845. [Google Scholar] [CrossRef]
- Fan, Y.; Fang, W.; Guo, S.; Pei, X.; Zhang, Y.; Xiao, Y.; Li, D.; Jin, K.; Bidochka, M.J.; Pei, Y. Increased insect virulence in Beauveria bassiana strains overexpressing an engineered chitinase. Appl. Environ. Microbiol. 2007, 73, 295–302. [Google Scholar] [CrossRef]
- Vega, F.E.; Meyling, N.V.; Luangsa-Ard, J.J.; Blackwell, M. Fungal Entomopathogens’. In Insect Pathology; Vega, F.E., Kaya, H.K., Eds.; Elsevier: London, UK, 2012; pp. 171–220. [Google Scholar]
- Mazza, G.; Francardi, V.; Simoni, S.; Benvenuti, C.; Cervo, R.; Faleiro, J.R.; Llácer, E.; Longo, S.; Nannelli, R.; Tarasco, E.; et al. An overview on the natural enemies of Rhynchophorus palm weevils, with focus on R. ferrugineus. Biol. Control 2014, 77, 83–92. [Google Scholar] [CrossRef]
- Khun, K.K.; Wilson, B.A.; Stevens, M.M.; Huwer, R.K.; Ash, G.J. Integration of entomopathogenic fungi into IPM programs: Studies involving weevils (Coleoptera: Curculionoidea) affecting horticultural crops. Insects 2020, 11, 659. [Google Scholar] [CrossRef]
- Ment, D.; Levy, N.; Allouche, A.; Davidovitz, M.; Yaacobi, G. Efficacy of entomopathogenic fungi as prevention against early life stages of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in laboratory and greenhouse trials. Insects 2023, 14, 918. [Google Scholar] [CrossRef]
- Yasin, M.; Wakil, W.; El-Shafie, H.A.F.; Bedford, G.O.; Miller, T.A. Potential role of microbial pathogens in control of red palm weevil (Rhynchophorus ferrugineus)—A review. Entomol. Res. 2017, 47, 219–234. [Google Scholar] [CrossRef]
- Mkiga, A.M.; Mohamed, S.A.; Plessis, H.D.; Khamis, F.M.; Akutse, K.S.; Ekesi, S. Metarhizium anisopliae and Beauveria bassiana: Pathogenicity, horizontal transmission, and their effects on reproductive potential of Thaumatotibia leucotreta (Lepidoptera: Tortricidae). J. Econ. Entomol. 2020, 113, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Ekesi, S.; Dimbi, S.; Maniania, N.K. The role of entomopathogenic fungi in the integrated management of tephritid fruit flies (Diptera: Tephritidae) with emphasis on species occurring in Africa. In Use of Entomopathogenic Fungi in Biological Pest Management; Ekesi, S., Maniania, N.K., Eds.; Research SignPost: Kerala, India, 2007; pp. 239–274. [Google Scholar]
- Matveev, S.; Reingold, V.; Yossef, E.; Levy, N.; Kottakota, C.; Mechrez, G.; Ment, D. The dissemination of Metarhizium brunneum conidia by females of the red palm weevil, Rhynchophorus ferrugineus, suggests a new mechanism for prevention practices. J. Fungi 2023, 9, 458. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Martin-Carballo, I.; Garrido-Jurado, I.; Santiago-Álvarez, C. Horizontal transmission of Metarhizium anisopliae among laboratory populations of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Biol. Control 2008, 47, 115–124. [Google Scholar] [CrossRef]
- Brandl, M.A.; Schumann, M.; Przyklenk, M.; Patel, A.; Vidal, S. Wireworm damage reduction in potatoes with an attract-and-kill strategy using Metarhizium brunneum. J. Pest Sci. 2017, 90, 479–493. [Google Scholar] [CrossRef]
- Yiğit, A.U. Auto-dissemination of Cordyceps fumosorosea amongst adult females of the two-spotted spider mite. Exp. Appl. Acarol. 2023, 91, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Gardescu, S.; Hajek, A.E. Transmission of Metarhizium brunneum conidia between male and female Anoplophora glabripennis adults. BioControl 2011, 56, 771–780. [Google Scholar] [CrossRef]
- Wakil, W.; Yasin, M.; Shapiro-Ilan, D. Effects of single and combined applications of entomopathogenic fungi and nematodes against Rhynchophorus ferrugineus (Olivier). Sci. Rep. 2017, 7, 5971. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Evans, H.C.; Latge, J.P. Atlas of Entomopathogenic Fungi; Springer: Berlin\Heidelberg, Germany, 1988; p. 187. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; The American Phytopathological Society Press: St. Paul, MN, USA, 1998; p. 218. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi, 2nd ed.; IHW-Verlag: Eching, Germany, 2007; p. 672. [Google Scholar]
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Humber, R.A. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 151–187. [Google Scholar]
- Wakil, W.; Ghazanfar, M.U.; Riasat, T.; Kwon, Y.J.; Qayyum, M.A.; Yasin, M. Occurrence and diversity of entomopathogenic fungi in cultivated and uncultivated soils in Pak. Entomol. Res. 2013, 43, 70–78. [Google Scholar] [CrossRef]
- Wakil, W.; Ghazanfar, M.U.; Yasin, M. Naturally occurring entomopathogenic fungi infecting stored grain insect species in Punjab, Pakistan. J. Insect Sci. 2014, 14, 1–7. [Google Scholar] [CrossRef]
- Inglis, G.D.; Enkerli, J.; Goettel, M.S. Laboratory techniques used for entomopathogenic fungi: Hypocreales. In Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 189–253. [Google Scholar]
- Dembilio, Ó.; Quesada-Moraga, E.; Santiago-Álvarez, C.; Jacas, J.A. Potential of an indigenous strain of the entomopathogenic fungus Beauveria bassiana as a biological control agent against the red palm weevil, Rhynchophorus ferrugineus. J. Invertebr. Pathol. 2010, 104, 214–221. [Google Scholar] [CrossRef]
- Martín, M.M.; Cabello, T. Manejo de la cría del picudo rojo de la palmera, Rhynchophorus ferrugineus (Olivier, 1790) (Coleoptera, Dryophthoridae), en dieta artificial y efectos en su biometría y biología. Bol. Sanid. Veg. Plagas. 2006, 32, 631–641. [Google Scholar]
- Lo Verde, G.; Torta, L.; Mondello, V.; Caldarella, C.G. Pathogenicity bioassays of isolates of Beauveria bassiana on Rhynchophorus ferrugineus. Pest Manag. Sci. 2015, 71, 323–328. [Google Scholar] [CrossRef]
- Marannino, P.; Santiago-Álvarez, C.; de Lillo, E.; Quesada-Moraga, E. A new bioassay method reveals pathogenicity of Metarhizium anisopliae and Beauveria bassiana against early stages of Capnodis tenebrionis (Coleoptera: Buprestidae). J. Invertebr. Pathol. 2006, 93, 210–213. [Google Scholar] [CrossRef]
- Hanusz, Z.; Tarasińska, J. Normalization of the Kolmogorov–Smirnov and Shapiro–Wilk tests of normality. Biom. Lett. 2015, 52, 85–93. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry; Freeman: New York, NY, USA, 1995. [Google Scholar]
- Minitab, LLC. Getting Started with Minitab 18; Minitab Inc.: State College, PA, USA, 2017; p. 73. [Google Scholar]
- Bidochka, M.J.; Kasperski, J.E.; Wild, G.A.M. Occurrence of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana in soils from temperate and near-northern habitats. Can. J. Bot. 1998, 76, 1198–1204. [Google Scholar]
- Qayyum, M.A.; Bilal, H.; Ali, H.; Raza, H.; Wajid, M. Factors affecting the epizootics of entomopathogenic fungi—A review. J. Bioresour. Manag. 2021, 8, 78–85. [Google Scholar] [CrossRef]
- Sun, X.; Yan, W.; Qin, W.; Zhang, J.; Niu, X.; Ma, G.; Li, F. Screening of tropical isolates of Metarhizium anisopliae for virulence to the red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae). SpringerPlus 2016, 5, 1100. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-H.; Wu, L.-H.; Liao, C.-T.; Li, D.; Shin, T.Y.; Kim, J.S.; Nai, Y.-S. Entomopathogenic fungi-mediated biological control of the red palm weevil Rhynchophorus ferrugineus. J. Asia-Pac. Entomol. 2023, 26, 102037. [Google Scholar] [CrossRef]
- Serna-Domínguez, M.G.; Andrade-Michel, G.Y.; Rosas-Valdez, R.; Castro-Félix, P.; Arredondo-Bernal, H.C.; Gallou, A. High genetic diversity of the entomopathogenic fungus Beauveria bassiana in Colima, Mexico. J. Invertebr. Pathol. 2019, 163, 67–74. [Google Scholar] [CrossRef]
- Qayyum, M.A.; Saeed, S.; Wakil, W.; Nawaz, A.; Iqbal, N.; Yasin, M.; Alamri, S. Diversity and correlation of entomopathogenic and associated fungi with soil factors. J. King Saud Univ.-Sci. 2021, 33, 101520. [Google Scholar] [CrossRef]
- Ullah, S.; Raza, M.; Alkafafy, M.; Sayed, S.; Hamid, M.I.; Majeed, M.Z.; Riaz, M.A.; Gaber, N.M.; Asim, M. Isolation, identification and virulence of indigenous entomopathogenic fungal strains against the peach-potato aphid, Myzus persicae Sulzer (Hemiptera: Aphididae), and the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). Egypt. J. Biol. Pest Control. 2022, 32, 1–11. [Google Scholar] [CrossRef]
- Soares, G.G.; Marchal, M.; Ferron, P. Susceptibility of Otiorhynchus sulcatus (Coleoptera: Curculionidae) larvae to Metarhizium anisopliae and Metarhizium flavoviridae (Deuteromycotina: Hyphomycetes) at two different temperatures. Environ. Entomol. 1983, 12, 1886–1890. [Google Scholar] [CrossRef]
- Poprawski, T.J.; Marchal, M.; Robert, P.-H. Comparative susceptibility of Otiorhynchus sulcatus and Sitona lineatus (Coleoptera: Curculionidae) early stages to five entomopathogenic hyphomycetes. Environ. Èntomol. 1985, 14, 247–253. [Google Scholar] [CrossRef]
- Gindin, G.; Levski, S.; Glazer, I.; Soroker, V. Evaluation of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana against the red palm weevil Rhynchophorus ferrugineus. Phytoparasitica. 2006, 34, 370–379. [Google Scholar] [CrossRef]
- Güerri-Agulló, B.; López-Follana, R.; Asensio, L.; Barranco, P.; Lopez-Llorca, L.V. Use of a solid formulation of Beauveria bassiana for biocontrol of the red palm weevil (Rhynchophorus ferrugineus) (Coleoptera: Dryophthoridae) under field conditions in SE Spain. Fla. Entomol. 2011, 94, 737–747. [Google Scholar] [CrossRef]
- Francardi, V.; Benvenuti, C.; Barzanti, G.P.; Roversi, P.F. Autocontamination trap with entomopathogenic fungi: A possible strategy in the control of Rhynchophorus ferrugineus (Olivier, 1790) (Coleoptera, Curculionidae). Redia 2013, 96, 57–67. [Google Scholar]
- Liu, H.; Skinner, M.; Brownbridge, M.; Parker, B. Characterization of Beauveria bassiana and Metarhizium anisopliae isolates for management of tarnished plant bug, Lygus lineolaris (Hemiptera: Miridae). J. Invertebr. Pathol. 2003, 82, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, M.A.; Saleem, M.A.; Saeed, S.; Wakil, W.; Ishtiaq, M.; Ashraf, W.; Ahmed, N.; Ali, M.; Ikram, R.M.; Yasin, M.; et al. Integration of entomopathogenic fungi and eco-friendly insecticides for management of red palm weevil, Rhynchophorus ferrugineus (Olivier). Saudi J. Biol. Sci. 2020, 27, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Riasat, T.; Wakil, W.; Ashfaq, M.; Sahi, S.T. Effect of Beauveria bassiana mixed with diatomaceous earth on mortality, mycosis and sporulation of Rhyzopertha dominica on stored wheat. Phytoparasitica 2011, 39, 325–331. [Google Scholar] [CrossRef]
- Hajek, A.E.; Everest, T.A.; Clifton, E.H. Accumulation of fungal pathogens infecting the invasive spotted lanternfly, Lycorma delicatula. Insects 2023, 14, 912. [Google Scholar] [CrossRef]
- Goettel, M.S.; Eilenberg, J.; Glare, T. Entomopathogenic fungi and their role in regulation of insect populations. In Insect Control: Biological and Synthetic Agents; Gilbert, L.I., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 361–405. [Google Scholar]
- Güerri-Agulló, B.; Gómez-Vidal, S.; Asensio, L.; Barranco, P.P.; Lopez-Llorca, L.V. Infection of the red palm weevil (Rhynchophorus ferrugineus) by the entomopathogenic fungus Beauveria bassiana: An SEM study. Microsc. Res. Techn. 2010, 73, 714–725. [Google Scholar] [CrossRef]
- Ansari, M.A.; Butt, T.M. Susceptibility of different developmental stages of large pine weevil Hylobius abietis (Coleoptera: Ccurculionidae) to entomopathogenic fungi and effect of fungal infection to adult weevils by formulation and application methods. J. Invertebr. Pathol. 2012, 111, 33–40. [Google Scholar] [CrossRef]
- Hajek, A.E.; Leger, R.J.S. Interactions between fungal pathogens and insect hosts. Annu. Rev. Entomol. 1994, 39, 293–322. [Google Scholar] [CrossRef]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Use of hyphomycete fungi for managing insect pests. In Fungi as Biocontrol Agents. Progress, Problems and Potential; Butt, T.M., Jackson, C.W., Magan, N., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 23–69. [Google Scholar]
- El Kichaoui, A.Y.; Asaker, B.A.A.; El-Hindi, M.W. Isolation, molecular identification and under lab evaluation of the entomopathogenic fungi M. anisopliae and B. bassiana against the red palm weevil R. ferrugineus in Gaza strip. Adv. Microbiol. 2017, 7, 109–124. [Google Scholar] [CrossRef][Green Version]
- Cherry, A.J.; Abalob, P.; Hell, K. A laboratory assessment of the potential of different strains of the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) to control Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in stored cowpea. J. Stored Prod. Res. 2005, 41, 295–309. [Google Scholar] [CrossRef]
- Dembilio, Ó.; Tapia, G.V.; Téllez, M.M.; Jacas, J.A. Lower temperature thresholds for oviposition and egg hatching of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in a Mediterranean climate. Bull. Entomol. Res. 2012, 102, 97–102. [Google Scholar] [CrossRef]
- Hou, F.J.; Addis, K.; Azmi, W.A. Virulence evaluation of entomopathogenic fungi against the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Dryopthoridae). Malays. Appl. Biol. 2018, 47, 25–30. [Google Scholar]
- Saleem, M.A.; Qayyum, M.A.; Ali, M.; Amin, M. Effect of sub-lethal doses of Beauveria bassiana and Nitenpyram on the development of red palm weevil, Rhynchophorus ferrugineus (Olivier). Pak. J. Zool. 2018, 51, 559–565. [Google Scholar] [CrossRef]
- Al-Manie, M.; Alkanhal, I. Acoustic detection of the red date palm weevil. Trans. Eng. Comput. Technol. 2004, 2, 209–212. [Google Scholar]
- Toledo, J.; Campos, S.E.; Flores, S.; Liedo, P.; Barrera, J.F.; Villaseñor, A.; Montoya, P. Horizontal transmission of Beauveria bassiana in Anastrepha ludens (Diptera: Tephritidae) under laboratory and field cage conditions. J. Econ. Entomol. 2007, 100, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Furlong, M.J.; Pell, J.K. Horizontal transmission of entomopathogenic fungi by the diamondback moth. Biol. Control 2001, 22, 288–299. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Santos-Quiros, R.; Valverde-Garcia, P.; Santiago-Álvarez, C. Virulence, horizontal transmission, and sublethal reproductive effects of Metarhizium anisopliae (anamorphic fungi) on the German cockroach (Blattodea: Blattellidae). J. Invertebr. Pathol. 2004, 87, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Scholte, E.J.; Knols, L.; Takken, W. Autodissemination of the entomopathogenic fungus Metarhizium anisopliae amongst adult of the malaria vector Anopheles gambiae. Malar. J. 2004, 3, 45. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Maranhão, E.A.; Valverde-Garcia, P.; Santiago-Álvarez, C. Selection of Beauveria bassiana isolates for control of the whiteflies Bemisia tabaci and Trialeurodes vaporarium on the basis of their virulence, thermal requirement and toxicogenic activity. Biol. Control 2006, 36, 274–287. [Google Scholar] [CrossRef]
- Wai, Y.K.; Bakar, A.A.; Azmi, W.A. Fecundity, fertility and survival of red palm weevil (Rhynchophorus ferrugineus) larvae reared on Sago palm. Sains Malays. 2015, 44, 1371–1375. [Google Scholar]
- Kaaya, G.P.; Okech, M.A. Horizontal transmission of mycotic infection in adult tsetse, Glossina morsitans morsitans. Entomophaga 1990, 35, 589–600. [Google Scholar] [CrossRef]
- Mulock, B.S.; Chandler, L.D. Effect of Beauveria bassiana on the fecundity of Western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae). Biol. Control. 2001, 22, 16–21. [Google Scholar] [CrossRef][Green Version]
- Castillo, M.-A.; Moya, P.; Hernández, E.; Primo-Yúfera, E. Susceptibility of Ceratitis capitata Wiedemann (Diptera: Tephritidae) to entomopathogenic fungi and their extracts. Biol. Control. 2000, 19, 274–282. [Google Scholar] [CrossRef]
- Meadow, R.; Vandenberg, J.D.; Shelton, A.M. Exchange of inoculum of Beauveria bassiana (Bals.) Vuill. (Hyphomycetes) between adult flies of the cabbage maggot Delia radicum L. (Diptera: Anthomyiidae). Biocontrol Sci. Technol. 2000, 10, 479–485. [Google Scholar] [CrossRef]
- Fragues, J.; Delmas, J.C.; Augé, J.; Lebrun, R.A. Fecundity and egg fertility in the adult Colorado beetle (Leptinostarsa decimilineata) surviving larval infection by the fungus Beauveria bassiana. Entomol. Exp. Appl. 1991, 61, 45–51. [Google Scholar] [CrossRef]
- Sikura, A.I.; Sikura, L.V.; Trebesava, R.M. Influence of white muscardine fungus (Beauveria bassiana Balsamo Vuillemin) on the reproductive system of the Colorado potato beetle. Zashch. Rast. Kichinev 1972, 2, 89–97. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Stage | Isolate Code | LC50 (conidia/mL) | 95% Fiducial Limits | Slope | X2 (df = 2) | p |
|---|---|---|---|---|---|---|
| Larvae | WG-08 | 1.72 × 106 | 7.67 × 105–3.14 × 106 | 0.24 ± 0.02 | 1.42 | 0.73 |
| WG-09 | 8.25 × 105 | 3.30 × 105–1.58 × 106 | 0.25 ± 0.02 | 3.05 | 0.81 | |
| WG-23 | 4.30 × 105 | 1.81 × 105–7.57 × 105 | 0.39 ± 0.05 | 7.19 | 0.63 | |
| WG-24 | 4.96 × 105 | 1.88 × 105–9.70 × 105 | 0.28 ± 0.03 | 2.19 | 0.47 | |
| WG-25 | 2.61 × 105 | 8.29 × 104–5.18 × 105 | 0.38 ± 0.05 | 5.04 | 0.68 | |
| Adults | WG-08 | 3.15 × 107 | 1.66 × 107–5.97 × 107 | 0.17 ± 0.02 | 0.27 | 0.56 |
| WG-09 | 8.20 × 106 | 3.92 × 106–1.50 × 107 | 0.18 ± 0.02 | 3.29 | 0.91 | |
| WG-23 | 1.36 × 106 | 5.63 × 105–2.60 × 106 | 0.23 ± 0.02 | 1.94 | 0.61 | |
| WG-24 | 3.63 × 106 | 1.80 × 106–6.33 × 106 | 0.23 ± 0.02 | 1.40 | 0.83 | |
| WG-25 | 8.97 × 105 | 4.28 × 105–1.54 × 106 | 0.31 ± 0.03 | 5.75 | 0.24 |
| Isolate Code | Concentration | LT50 (d) | 95% Fiducial Limits | Slope | X2 (df = 2) | p |
|---|---|---|---|---|---|---|
| WG-08 | 106 | 22.28 | 20.00–26.07 | 0.07 ± 0.01 | 0.83 | 0.36 |
| 107 | 16.52 | 15.00–18.37 | 0.08 ± 0.01 | 0.40 | 0.71 | |
| 108 | 10.93 | 9.26–12.30 | 0.09 ± 0.01 | 2.23 | 0.53 | |
| 109 | 7.54 | 5.68–8.90 | 0.12 ± 0.01 | 0.29 | 0.47 | |
| WG-09 | 106 | 19.58 | 17.63–22.55 | 0.07 ± 0.01 | 0.31 | 0.76 |
| 107 | 14.12 | 12.64–15.61 | 0.08 ± 0.01 | 0.75 | 0.35 | |
| 108 | 8.98 | 7.00–10.45 | 0.09 ± 0.01 | 1.80 | 0.92 | |
| 109 | 5.59 | 3.28–7.15 | 0.12 ± 0.01 | 1.21 | 0.80 | |
| WG-23 | 106 | 14.28 | 12.57–15.84 | 0.08 ± 0.01 | 0.49 | 0.44 |
| 107 | 11.71 | 10.36–12.91 | 0.07 ± 0.01 | 0.19 | 0.28 | |
| 108 | 7.52 | 6.13–8.58 | 0.16 ± 0.01 | 5.68 | 0.68 | |
| 109 | 3.64 | 0.66 -5.39 | 0.15 ± 0.02 | 1.35 | 0.74 | |
| WG-24 | 106 | 17.49 | 15.81–19.75 | 0.08 ± 0.01 | 0.71 | 0.37 |
| 107 | 13.21 | 11.78–14.57 | 0.09 ± 0.01 | 0.69 | 0.56 | |
| 108 | 8.24 | 6.63–9.48 | 0.12 ± 0.01 | 0.30 | 0.29 | |
| 109 | 4.49 | 1.75–6.24 | 0.12 ± 0.01 | 0.11 | 0.83 | |
| WG-25 | 106 | 14.28 | 12.75–15.84 | 0.08 ± 0.01 | 0.82 | 0.49 |
| 107 | 9.67 | 8.00–10.99 | 0.10 ± 0.01 | 0.22 | 0.75 | |
| 108 | 6.50 | 4.86–7.68 | 0.16 ± 0.01 | 3.71 | 0.19 | |
| 109 | 3.24 | 0.308–4.91 | 0.20 ± 0.04 | 0.14 | 0.38 |
| Isolate Code | Concentration | LT50 (d) | 95% Fiducial Limits | Slope | X2 (df = 2) | p |
|---|---|---|---|---|---|---|
| WG-08 | 106 | 31.65 | 26.03–46.38 | 0.05 ± 0.01 | 0.28 | 0.45 |
| 107 | 23.63 | 20.84–28.80 | 0.06 ± 0.01 | 0.08 | 0.16 | |
| 108 | 18.46 | 16.59–21.19 | 0.07 ± 0.01 | 0.36 | 0.37 | |
| 109 | 12.18 | 10.10–13.92 | 0.07 ± 0.01 | 0.61 | 0.62 | |
| WG-09 | 106 | 25.81 | 22.48–32.43 | 0.06 ± 0.01 | 0.28 | 0.43 |
| 107 | 21.35 | 18.96–25.50 | 0.06 ± 0.01 | 0.47 | 0.51 | |
| 108 | 15.91 | 14.17–18.00 | 0.07 ± 0.01 | 0.17 | 0.73 | |
| 109 | 9.76 | 7.73–11.28 | 0.09 ± 0.01 | 0.15 | 0.28 | |
| WG-23 | 106 | 21.40 | 19.17–25.09 | 0.07 ± 0.01 | 0.26 | 0.56 |
| 107 | 16.25 | 14.67–18.17 | 0.08 ± 0.01 | 0.52 | 0.67 | |
| 108 | 11.56 | 9.97–12.92 | 0.09 ± 0.01 | 0.85 | 0.25 | |
| 109 | 7.07 | 4.81–8.64 | 0.10 ± 0.01 | 3.77 | 0.92 | |
| WG-24 | 106 | 24.85 | 21.65–31.16 | 0.06 ± 0.01 | 0.25 | 0.58 |
| 107 | 18.73 | 16.84–21.52 | 0.07 ± 0.01 | 0.63 | 0.24 | |
| 108 | 13.34 | 11.86–14.75 | 0.09 ± 0.01 | 0.32 | 0.69 | |
| 109 | 8.24 | 6.16–9.75 | 0.10 ± 0.01 | 1.22 | 0.47 | |
| WG-25 | 106 | 19.58 | 17.63–22.57 | 0.07 ± 0.01 | 0.12 | 0.84 |
| 107 | 14.11 | 12.63–15.60 | 0.08 ± 0.01 | 0.44 | 0.99 | |
| 108 | 9.78 | 8.35–10.96 | 0.12 ± 0.01 | 0.61 | 0.68 | |
| 109 | 5.74 | 3.49–7.25 | 0.13 ± 0.01 | 10.57 | 0.71 |
| Isolate Code | Hatching (%) | Average Survival Time (AST) | ||
|---|---|---|---|---|
| 1 × 106 conidia/mL | 1 × 108 conidia/mL | 1 × 106 conidia/mL | 1 × 108 conidia/mL | |
| WG-08 | 75.55 ± 1.92 b | 46.67 ± 1.92 b | 3.02 ± 0.06 a | 1.51 ± 0.05 a |
| WG-09 | 68.14 ± 1.85 b | 42.96 ± 1.17 b | 2.55 ± 0.07 b | 1.20 ± 0.09 ab |
| WG-23 | 54.07 ± 1.73 c | 27.40 ± 1.33 d | 1.93 ± 0.08 cd | 0.73 ± 0.08 cd |
| WG-24 | 61.48 ± 1.48 b | 35.55 ± 1.11 c | 2.11 ± 0.07 c | 1.02 ± 0.08 bc |
| WG-25 | 49.63 ± 2.25 c | 18.51 ± 0.97 e | 1.71 ± 0.06 d | 0.55 ± 0.06 d |
| Control | 78.51 ± 0.97 a | 72.52 ± 0.74 a | - | - |
| df | 5, 53 | 5, 53 | 4, 44 | 4, 44 |
| F | 43.8 | 219 | 52.7 | 22.6 |
| p | <0.01 | <0.01 | <0.01 | <0.01 |
| Isolate Code | Treatment | Mortality | ||
|---|---|---|---|---|
| Female | Male | Female | Male | |
| WG-08 | Non-infected | Infected | 11.11 ± 3.51 a | 27.22 ± 3.23 a |
| Infected | Non-infected | 15.55 ± 2.93 a | 13.33 ± 3.33 b | |
| Infected | Infected | 20.00 ± 3.33 a | 36.11 ± 2.60 a | |
| F | 29.46 | 21.63 | ||
| p | <0.01 | <0.01 | ||
| WG-09 | Non-infected | Infected | 23.33 ± 2.20 b | 43.88 ± 3.70 a |
| Infected | Non-infected | 29.44 ± 4.74 ab | 25.00 ± 2.88 b | |
| Infected | Infected | 36.66 ± 2.20 a | 47.77 ± 3.23 a | |
| F | 33.17 | 24.35 | ||
| p | <0.01 | <0.01 | ||
| WG-23 | Non-infected | Infected | 36.66 ± 3.99 b | 54.44 ± 2.93 b |
| Infected | Non-infected | 48.33 ± 4.24 ab | 41.11 ± 3.51 c | |
| Infected | Infected | 53.33 ± 2.88 a | 67.77 ± 4.00 a | |
| F | 27.36 | 21.53 | ||
| p | <0.01 | <0.01 | ||
| WG-24 | Non-infected | Infected | 33.88 ± 3.09 a | 47.77 ± 3.23 a |
| Infected | Non-infected | 36.11 ± 4.23 a | 33.88 ± 3.09 b | |
| Infected | Infected | 40.55 ± 5.55 a | 56.66 ± 4.08 a | |
| F | 25.93 | 32.69 | ||
| p | <0.01 | <0.01 | ||
| WG-25 | Non-infected | Infected | 45.55 ± 2.93 b | 64.44 ± 4.12 b |
| Infected | Non-infected | 54.44 ± 2.93 ab | 46.66 ± 2.88 c | |
| Infected | Infected | 61.66 ± 1.66 a | 78.88 ± 3.41 a | |
| F | 35.78 | 22.53 | ||
| p | <0.01 | <0.01 | ||
| Isolate Code | Treatment of Pairs | Oviposition Rate | Fecundity | Egg Hatching (%) | Larval Survival | |
|---|---|---|---|---|---|---|
| Female | Male | |||||
| WG-08 | Control | 3.28 ± 0.10 a | 61.62 ± 1.68 a | 70.37 ± 1.32 a | 80.38 ± 0.86 a | |
| Non-infected | Infected | 3.04 ± 0.08 ab | 46.22 ± 1.32 b | 67.72 ± 1.73 ab | 78.85 ± 1.21 ab | |
| Infected | Non-infected | 2.86 ± 0.10 b | 41.62 ± 1.47 b | 64.02 ± 1.39 b | 75.27 ± 0.87 bc | |
| Infected | Infected | 2.28 ± 0.11 c | 32.26 ± 1.67 c | 55.55 ± 1.12 c | 71.49 ± 1.06 c | |
| F | 14.52 | 23.18 | 27.59 | 33.61 | ||
| p | <0.01 | <0.01 | <0.01 | <0.01 | ||
| WG-09 | Control | 2.93 ± 0.09 a | 54.11 ± 1.26 a | 68.78 ± 1.15 a | 86.27 ± 1.45 a | |
| Non-infected | Infected | 2.51 ± 0.12 b | 38.57 ± 1.24 b | 57.67 ± 1.24 b | 65.99 ± 0.86 b | |
| Infected | Non-infected | 2.08 ± 0.10 c | 33.57 ± 0.78 c | 53.43 ± 1.05 b | 63.31 ± 0.90 bc | |
| Infected | Infected | 1.75 ± 0.09 c | 27.02 ± 1.13 d | 46.03 ± 1.12 c | 58.67 ± 2.82 c | |
| F | 18.73 | 26.31 | 24.15 | 29.37 | ||
| p | <0.01 | <0.01 | <0.01 | <0.01 | ||
| WG-23 | Control | 3.20 ± 0.11 a | 56.02 ± 0.87 a | 73.01 ± 0.79 a | 84.12 ± 1.01 a | |
| Non-infected | Infected | 1.24 ± 0.11 b | 26.08 ± 1.74 b | 35.97 ± 1.15 b | 47.00 ± 1.68 b | |
| Infected | Non-infected | 1.01 ± 0.06 bc | 20.60 ± 1.06 c | 30.15 ± 1.37 c | 43.59 ± 2.32 b | |
| Infected | Infected | 0.75 ± 0.05 c | 13.00 ± 0.37 d | 23.28 ± 2.16 d | 41.75 ± 2.99 b | |
| F | 21.66 | 19.27 | 34.12 | 38.80 | ||
| p | <0.01 | <0.01 | <0.01 | <0.01 | ||
| WG-24 | Control | 2.86 ± 0.03 a | 57.28 ± 1.20 a | 64.55 ± 0.83 a | 77.83 ± 0.28 a | |
| Non-infected | Infected | 2.17 ± 0.10 b | 33.06 ± 0.62 b | 46.03 ± 1.37 b | 55.08 ± 1.50 b | |
| Infected | Non-infected | 1.73 ± 0.08 c | 25.44 ± 0.56 c | 44.44 ± 1.12 b | 47.77 ± 1.96 c | |
| Infected | Infected | 1.20 ± 0.10 d | 18.35 ± 0.48 d | 39.68 ± 1.37 c | 42.69 ± 1.67 c | |
| F | 25.84 | 31.36 | 39.04 | 44.59 | ||
| p | <0.01 | <0.01 | <0.01 | <0.01 | ||
| WG-25 | Control | 3.07 ± 0.07 a | 53.55 ± 1.16 a | 65.07 ± 0.79 a | 81.31 ± 1.23 a | |
| Non-infected | Infected | 1.00 ± 0.07 b | 17.13 ± 0.54 b | 21.69 ± 0.83 b | 33.33 ± 2.63 b | |
| Infected | Non-infected | 0.86 ± 0.06 b | 15.04 ± 0.69 b | 16.40 ± 0.83 c | 29.63 ± 1.46 b | |
| Infected | Infected | 0.57 ± 0.04 c | 11.75 ± 0.74 c | 11.64 ± 0.83 d | 25.92 ± 6.86 b | |
| F | 28.01 | 37.96 | 46.18 | 51.06 | ||
| p | <0.01 | <0.01 | <0.01 | <0.01 | ||
| Treatments | Palm Infested | Number of Larvae | Efficacy (%) |
|---|---|---|---|
| WG-08 | 80.00 ± 0.00 ab | 36.05 ± 1.75 a | 16.91 ± 7.47 d |
| WG-09 | 65.00 ± 9.57 bc | 27.15 ± 1.94 b | 37.99 ± 3.86 c |
| WG-23 | 40.00 ± 8.16 cd | 13.30 ± 2.08 c | 69.38 ± 5.13 b |
| WG-24 | 55.00 ± 5.00 bcd | 19.45 ± 1.32 bc | 55.28 ± 3.93 bc |
| WG-25 | 35.00 ± 9.57 d | 4.15 ± 0.95 d | 90.50 ± 2.14 a |
| Control | 100.00 ± 0.00 a | 43.85 ± 1.90 a | - |
| df | 5, 119 | 5, 119 | 4, 99 |
| F | 17.78 | 70.57 | 43.33 |
| p | <0.01 | <0.01 | <0.01 |
| Interval (d) | Treatments | Palm Infest | Number of Larvae | Efficacy (%) |
|---|---|---|---|---|
| 1 | WG-08 | 100.00 ± 0.00 a | 37.15 ± 2.01 b | 27.62 ± 4.38 d |
| WG-09 | 80.00 ± 8.16 ab | 32.05 ± 2.61 bc | 37.92 ± 3.66 cd | |
| WG-23 | 45.00 ± 15.00 bc | 17.75 ± 2.20 d | 65.69 ± 3.62 ab | |
| WG-24 | 60.00 ± 8.16 bc | 25.00 ± 1.87 cd | 51.06 ± 4.69 bc | |
| WG-25 | 30.00 ± 10.00 c | 9.10 ± 0.69 e | 82.21 ± 1.64 a | |
| Control | 100.00 ± 0.00 a | 51.45 ± 1.57 a | - | |
| df | 5, 119 | 5, 119 | 4, 99 | |
| F | 12.30 | 65.40 | 32.71 | |
| p | <0.01 | <0.01 | <0.01 | |
| 15 | WG-08 | 100.00 ± 0.00 a | 31.70 ± 1.53 b | 34.10 ± 5.45 d |
| WG-09 | 85.00 ± 9.57 ab | 26.65 ± 2.29 bc | 44.89 ± 5.06 cd | |
| WG-23 | 30.00 ± 5.77 c | 12.75 ± 2.35 de | 72.84 ± 6.15 ab | |
| WG-24 | 50.00 ± 12.91 bc | 19.50 ± 1.29 cd | 59.78 ± 2.55 bc | |
| WG-25 | 20.00 ± 8.16 c | 6.90 ± 0.38 e | 85.77 ± 0.76 a | |
| Control | 100.00 ± 0.00 a | 48.75 ± 2.98 a | - | |
| df | 5, 119 | 5, 119 | 4, 99 | |
| F | 18.43 | 55.17 | 37.52 | |
| p | <0.01 | <0.01 | <0.01 | |
| 30 | WG-08 | 100.00 ± 0.00 a | 38.50 ± 2.72 ab | 22.16 ± 8.27 c |
| WG-09 | 85.00 ± 9.57 a | 29.25 ± 2.80 bc | 41.52 ± 5.08 bc | |
| WG-23 | 30.00 ± 12.91 b | 20.00 ± 3.17 cd | 59.03 ± 8.05 ab | |
| WG-24 | 80.00 ± 8.16 a | 25.50 ± 3.09 cd | 49.54 ± 4.14 ab | |
| WG-25 | 40.00 ± 8.16 b | 13.85 ± 1.15 d | 72.27 ± 2.28 a | |
| Control | 100.00 ± 0.00 a | 50.05 ± 2.12 a | - | |
| df | 5, 119 | 5, 119 | 4, 99 | |
| F | 12.95 | 23.57 | 10.81 | |
| p | <0.01 | <0.01 | <0.01 | |
| 45 | WG-08 | 100.00 ± 0.00 a | 42.30 ± 3.18 ab | 18.68 ± 8.55 c |
| WG-09 | 100.00 ± 0.00 a | 36.10 ± 3.51 bc | 30.18 ± 9.85 bc | |
| WG-23 | 80.00 ± 8.16 ab | 24.00 ± 2.79 cd | 53.55 ± 7.45 ab | |
| WG-24 | 100.00 ± 0.00 a | 34.45 ± 2.25 bc | 34.02 ± 5.44 bc | |
| WG-25 | 60.00 ± 14.14 b | 18.50 ± 3.05 d | 63.88 ± 7.54 a | |
| Control | 100.00 ± 0.00 a | 52.70 ± 2.77 a | - | |
| df | 5, 119 | 5, 119 | 4, 99 | |
| F | 6.63 | 15.51 | 11.16 | |
| p | <0.01 | <0.01 | <0.01 | |
| 60 | WG-08 | 100.00 ± 0.00 a | 45.50 ± 3.61 ab | 14.65 ± 7.23 c |
| WG-09 | 100.00 ± 0.00 a | 43.40 ± 3.60 ab | 18.34 ± 7.96 c | |
| WG-23 | 80.00 ± 8.16 b | 29.65 ± 2.69 cd | 43.90 ± 7.08 ab | |
| WG-24 | 100.00 ± 0.00 a | 37.80 ± 3.68 bc | 29.27 ± 7.04 bc | |
| WG-25 | 60.00 ± 8.16 c | 22.45 ± 2.31 d | 57.65 ± 5.47 a | |
| Control | 100.00 ± 0.00 a | 53.55 ± 1.99 a | - | |
| 5, 119 | 5, 119 | 4, 99 | df | |
| F | 15.75 | 14.44 | 10.47 | |
| p | <0.01 | <0.01 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alwaneen, W.S.; Wakil, W.; Kavallieratos, N.G.; Qayyum, M.A.; Tahir, M.; Rasool, K.G.; Husain, M.; Aldawood, A.S.; Shapiro-Ilan, D. Efficacy and Persistence of Entomopathogenic Fungi against Rhynchophorus ferrugineus on Date Palm: Host to Host Transmission. Agronomy 2024, 14, 642. https://doi.org/10.3390/agronomy14040642
Alwaneen WS, Wakil W, Kavallieratos NG, Qayyum MA, Tahir M, Rasool KG, Husain M, Aldawood AS, Shapiro-Ilan D. Efficacy and Persistence of Entomopathogenic Fungi against Rhynchophorus ferrugineus on Date Palm: Host to Host Transmission. Agronomy. 2024; 14(4):642. https://doi.org/10.3390/agronomy14040642
Chicago/Turabian StyleAlwaneen, Waleed S., Waqas Wakil, Nickolas G. Kavallieratos, Mirza Abdul Qayyum, Muhammad Tahir, Khawaja G. Rasool, Mureed Husain, Abdulrahman S. Aldawood, and David Shapiro-Ilan. 2024. "Efficacy and Persistence of Entomopathogenic Fungi against Rhynchophorus ferrugineus on Date Palm: Host to Host Transmission" Agronomy 14, no. 4: 642. https://doi.org/10.3390/agronomy14040642
APA StyleAlwaneen, W. S., Wakil, W., Kavallieratos, N. G., Qayyum, M. A., Tahir, M., Rasool, K. G., Husain, M., Aldawood, A. S., & Shapiro-Ilan, D. (2024). Efficacy and Persistence of Entomopathogenic Fungi against Rhynchophorus ferrugineus on Date Palm: Host to Host Transmission. Agronomy, 14(4), 642. https://doi.org/10.3390/agronomy14040642