


The Role of Carbon Nanotubes in Improving Drought Tolerance via Upregulation of the Physiological Processes of Peanut Plants Grown in Sandy Soils

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Water Irrigation Requirements
2.2. Chemical Analysis
2.2.1. In Peanut Shoots
2.2.2. In Peanut Seeds
2.3. ANN Modeling
2.4. Statistical Analysis
3. Results
3.1. Changes in Morphological Criteria
3.2. Changes in Photosynthetic Pigments
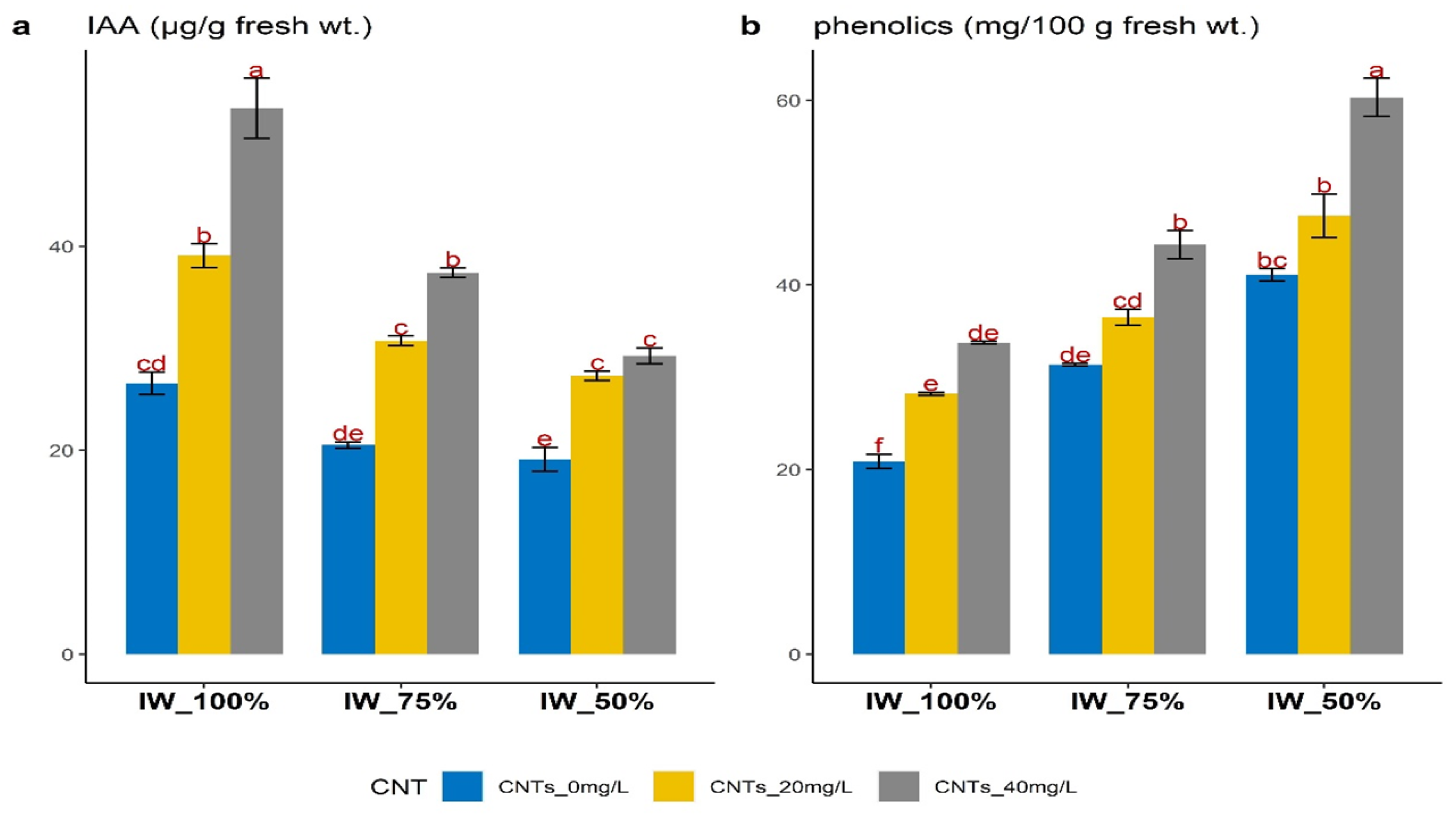
3.3. Changes in Endogenous IAA and Total Phenols
3.4. Changes in Osmolytes
3.5. Yield and Yield Components
3.6. Changes in Carbohydrates, Protein and Oil Contents
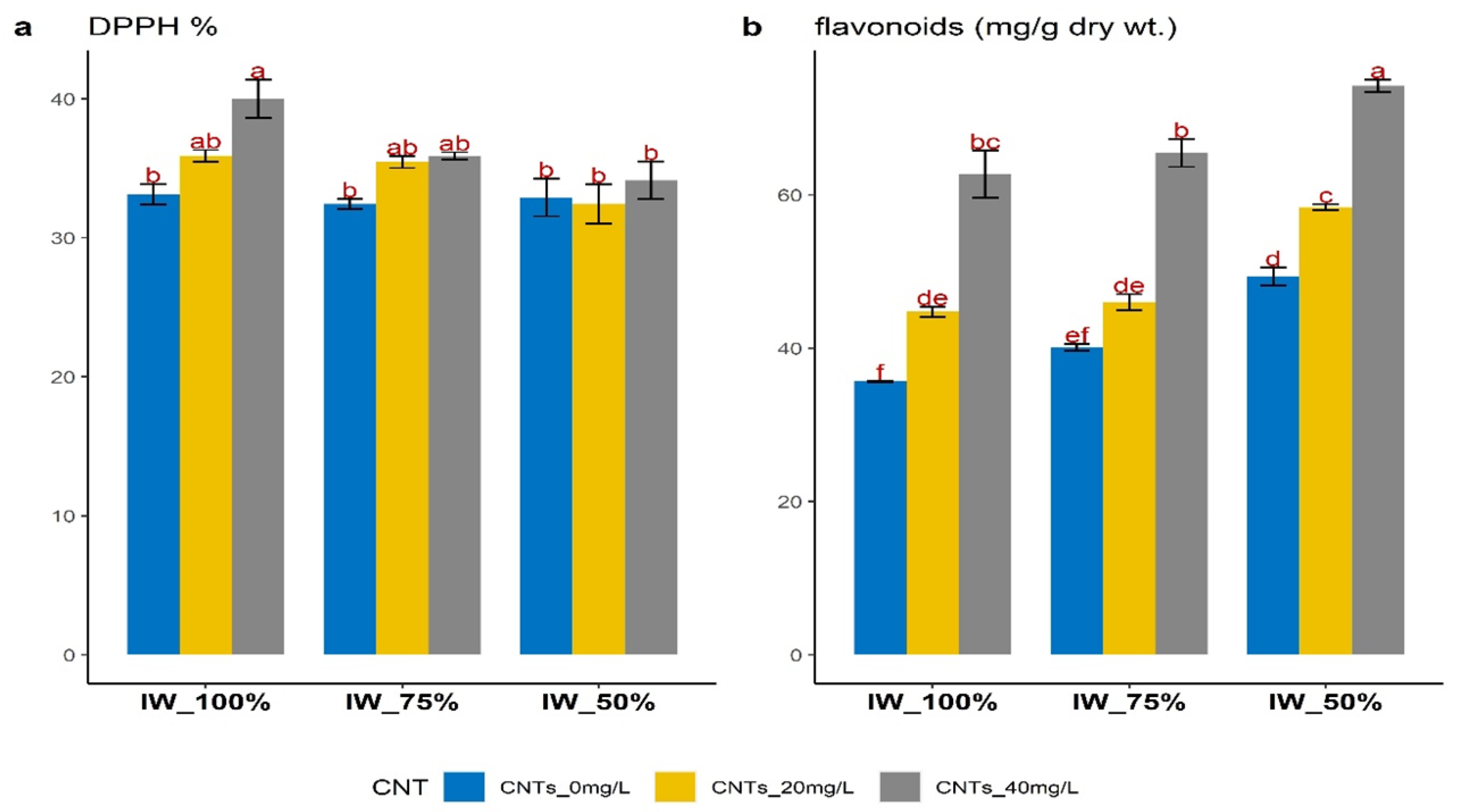
3.7. Changes in Antioxidant Activities (DPPH%) and Flavonoid Contents
3.8. Changes in Non-Photosynthetic Pigments
3.9. Artificial Neural Networks (ANNs)
3.10. Relative Importance of Agronomic Traits to Peanut Seed Yield
3.11. Yield Network Analysis
4. Discussion
4.1. Changes in Photosynthetic Pigments
4.2. Changes in Endogenous IAA and Total Phenols
4.3. Changes in Osmolytes
4.4. Changes in Morphological Criteria and Yield
4.5. Changes in Carbohydrates, Protein and Oil Contents
4.6. Changes in Antioxidant Activities (DPPH%), Flavonoid Contents and Non Photosynthetic Pigments
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vali, G.M.; Singh, S.; Sruthi, D.S.V.; Hinduja, N.; Talasila, V.; Tiwari, D. Effect of phosphorus and zinc on growth and yield of summer groundnut (Arachishypogaea L.). Bioscan 2020, 15, 535–540. [Google Scholar]
- Boote, K.J. Growth stages of peanut (Arachis hypogaea L.). Peanut Sci. 1982, 9, 35–40. [Google Scholar] [CrossRef]
- Bakry, A.B.; Sadak, M.S.; Abd El-Monem, A.A. Physiological aspects of tyrosine and salicylic acid on morphological, yield and biochemical constituents of peanut plants. Pak. J. Biol. Sci. 2020, 23, 375–384. [Google Scholar] [CrossRef]
- Velikova, V.; Petrova, N.; Kovács, L.; Petrova, A.; Koleva, D.; Tsonev, T.; Taneva, S.; Petrov, P.; Krumova, S. Single-Walled Carbon Nanotubes Modify Leaf Micromorphology, Chloroplast Ultrastructure and Photosynthetic Activity of Pea Plants. Int. J. Mol. Sci. 2021, 22, 4878. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Yan, F.; Zhu, Y.; Zhao, Y.; Wang, Y.; Li, J.; Wang, Q.; Liu, Y. De novo transcriptome sequencing and analysis of salt-, alkali-, and drought-responsive genes in Sophora alopecuroides. BMC Genom. 2020, 21, 423. [Google Scholar] [CrossRef] [PubMed]
- Dutta, T.; Neelapu, N.R.R.; Wani, S.H.; Challa, S. Compatible solute engineering of crop plants for improved tolerance toward abiotic stresses. In Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Wani, S.H., Ed.; Academic Press: London, UK, 2018; pp. 221–254. [Google Scholar]
- Nguyen, K.H.; Mostofa, M.G.; Watanabe, Y.; Tran, C.D.; Rahman, M.M.; Tran, L.S.P. Overexpression of GmNAC085 enhances drought tolerance in Arabidopsis by regulating glutathione biosynthesis, redox balance and glutathione-dependent detoxification of reactive oxygen species and methylglyoxal. Environ. Exp. Bot. 2019, 161, 242–254. [Google Scholar] [CrossRef]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Zakir, I.; Ali, M.A.; Ahmad, S. Oxidative stress and antioxidant defense in plants under drought conditions. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, The Netherlands, 2019; pp. 207–219. [Google Scholar]
- Bakry, B.A.; El-Nwehy, S.S.; Afify, R.R.; Ibrahim, O.M.; El-Aziz, M.A. Preparation, characterization and evaluation of nano Phosphorus foliar application on peanut production under sandy soil. Asian J. Plant Sci. 2022, 21, 423–431. [Google Scholar] [CrossRef]
- Singh, N.A. Nanotechnology, innovations, industrial applications and patents. Environ. Chem. Lett. 2017, 15, 185–191. [Google Scholar] [CrossRef]
- El-Bassiouny, H.M.S.; Abdallah, M.M.-S.; El-Enany, M.A.M.; Sadak, M.S. Nano-zinc oxide and Arbuscular mycorrhiza effects on physiological and biochemical aspects of wheat cultivars under saline conditions. Pak. J. Biol. Sci. 2020, 23, 478–490. [Google Scholar] [CrossRef]
- Abdalla, S.; Al-Marzouki, F.; Al-Ghamdi, A.A.; Abdel-Daiem, A. Different technical applications of carbon nanotubes. Nanoscale Res. Lett. 2015, 10, 358. [Google Scholar] [CrossRef]
- Abdallah, M.M.S.; Younis, A.E.S.M.; Bakry, B.A.; El-Bassiouny, H.M.S.; Ramadan, A.A.E.M. Potential impacts of carbon tube and silicon oxide nanoparticles on growth, yield and antioxidant system of soybean plant. Asian J. Plant Sci. 2020, 19, 495–507. [Google Scholar]
- Khodakovskaya, M.V.; Kim, B.S.; Kim, J.N.; Alimohammadi, M.; Dervishi, E.; Mustafa, T.; Cernigla, C.E. Carbon nanotubes as plant growth regulators: Effects on tomato growth, reproductive system and soil microbial community. Small 2012, 9, 115–123. [Google Scholar] [CrossRef]
- Chai, M.; Shi, F.; Li, R.; Liu, L.; Liu, Y.; Liu, F. Interactive effects of cadmium and carbon nanotubes on the growth and metal accumulation in a halophyte Spartinaalterniflora (Poaceae). Plant Growth Regul. 2013, 71, 171–179. [Google Scholar] [CrossRef]
- Mohamed, A.M.; Mahmoud, E.A.; Younes, N.A. Impact of foliar application of carbon nanotube and benzyladenine on broccoli growth and head yields. Arch. Agric. Sci. J. 2021, 4, 81–101. [Google Scholar] [CrossRef]
- Srivastava, A.; Rao, D.P. Enhancement of seed germination and plant growth of wheat, maize, peanut and garlic using multiwalled carbon nantubes. Eur. Chem. Bull. 2014, 3, 502–504. [Google Scholar]
- Rahimi, D.; Kartoolinejad, D.; Nourmohammadi, K.; Naghdi, R. Increasing drought resistance of Alnussub cordata C.A. Mey. seeds using a nano priming technique with multi-walled carbon nanotubes. J. For. Sci. 2016, 62, 269–278. [Google Scholar] [CrossRef]
- Karami, A.; Sepehri, A. Beneficial Role of MWCNTs and SNP on Growth, Physiological and Photosynthesis Performance of Barley under NaCl Stress. J. Soil. Sci. Plant Nutr. 2018, 18, 752–771. [Google Scholar] [CrossRef]
- Sadak, M.S.; Al Ashkar, N.M.; Bakry, B.A. Role of Carbon Nano Tubes for Improving Drought Tolerance via Upregulation of Physiological Attributes Processes of Peanut Plant Grown in Sandy Soils. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil Sampling and Methods of Analysis; CRC Press: Boca Raton, FL, USA, 2007; Available online: https://www.aweimagazine.com/ (accessed on 18 February 2024).
- Allen, M.E.J.; Wright, J.L.; Burman, R.D. Operational estimates of reference evapotranspiration. Agron. J. 1989, 81, 650–662. [Google Scholar] [CrossRef]
- Keller, J.; Karmeli, D. Trickle Irrigation Design; Rain Bird Sprinkler Manufacturing Corporation: Glendora, CA, USA, 1975; p. 133. [Google Scholar]
- Li, Y.; Chen, M. Novel chlorophylls and new directions in photosynthesis research. Funct. Plant Biol. 2015, 42, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Gusmiaty, M.; Restu, A.; Payangan, R.Y. Production of IAA (indole acetic acid) of the rhizosphere fungus in the Suren community forest stand. IOP Conf. Ser. Earth Environ. Sci. 2019, 343, 012058. [Google Scholar] [CrossRef]
- Siddiqui, N.; Rauf, A.; Latif, A.; Mahmood, Z. Spectrophotometric determination of the total phenolic content, spectral and fluorescence study of the herbal Unani drug Gul-e-Zoofa (Nepeta bracteate Benth). J. Taibah Uni. Med. Sci. 2017, 12, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Mecozzi, M. Estimation of total carbohydrate amount in environmental samples by the phenol-sulphuric acid method assisted by multivariate calibration. Chemom. Intell. Lab. Syst. 2005, 79, 84–90. [Google Scholar] [CrossRef]
- Vartanian, N.; Hervochon, P.; Marcotte, L.; Larher, F. Proline accumulation during drought rhizogenesis in Brassica napus var. oleifera. Plant Physiol. 1992, 140, 623–628. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline Metabolism and Its Implications for Plant-Environment Interaction. Arab. Book 2010, 8, e0140. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, P.; Pedrol, N. Chapter 22. Free Proline Quantification. In Handbook of Plant Ecophysiology Techniques; Reigosa, M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar] [CrossRef]
- Albalasmeh, A.A.; Berhe, A.A.; Ghezzehei, T.A. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Pedrol, N.; Tamayo, P.R. Protein content quantification by Bradford method. In Handbook of Plant Ecophysiology Techniques; Reigosa, M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 20th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Gyamfi, M.A.; Yonamine, M.; Aniya, Y. Free radical scavenging action of medicinal herbs from Ghana Thonningia sanguine on experimentally induced liver injuries. Gen Pharmacol. 2002, 32, 661–667. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogyo Gakkaish 1992, 39, 925–928. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of totalflavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Statistical Package for Social Sciences, Statistics for Windows, Version 17.0. Released 2008. SPSS Inc.: Chicago, USA. Available online: https://he02.tci-thaijo.org/index.php/ramajournal/statistical_software_references_format (accessed on 18 February 2024).
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics. In A Biometrical Approach, 3rd ed.; McGraw Hill Book Co. Inc.: New York, NY, USA, 1997; Available online: https://www.amazon.com/Principles-Procedures-Statistics-Biometrical-Approach/dp/0070610282 (accessed on 18 February 2024).
- Ibrahim, O.M.; El-Gamal, E.H.; Darwish, K.M.; Kianfar, N. Modeling main and interactional effects of some physiochemical properties of Egyptian soils on cation exchange capacity via artificial neural networks. Eurasian Soil Sci. 2022, 55, 1052–1063. [Google Scholar] [CrossRef]
- Elewa, T.A.; Sadak, M.S.; Saad, A.M. Proline treatment improves physiological responses in quinoa plants under drought stress. Biosci. Res. 2017, 14, 21–33. [Google Scholar]
- Sadak, M.S.; Bakry, B.A. Zinc-oxide and nano ZnO oxide effects on growth, some biochemical aspects, yield quantity, and quality of flax (Linum uitatissimum L.) in absence and presence of compost under sandy soil. Bull. Natl. Res. Cent. 2020, 44, 98. [Google Scholar] [CrossRef]
- Nazarbeygi, E.; Yazdi, H.L.; Naseri, R.; Soleimani, R. The effects of different levels of salinity on proline and a, b chlorophylls in canola. Am. Eurasian J. Agric. Environ. Sci. 2011, 10, 70–74. [Google Scholar]
- Fang, Z.; Bouwkamp, J.C.; Solomos, T. Chlorophyllaseactivities and chlorophyll degradation during leaf senescence innon-yellowing mutant and wild-type of Phaseolus vulgaris L. J. Exp. Bot. 1998, 49, 503–510. [Google Scholar]
- El Hariri, D.M.; Sadak, M.S.; El-Bassiouny, H.M.S. Response of flax cultivars to ascorbic acid and αtocopherol under salinity stress conditions. Inter. J. Acad. Res. 2010, 2, 101–109. [Google Scholar]
- Chen, Q.; Qi, W.B.; Reiter, R.J.; Wei, W.; Wang, B.M. Exogenously applied melatonin stimulates root growthand raises endogenous indole acetic acid in roots of etiolated seedlings of Brassica juncea. J. Plant Physiol. 2009, 166, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Liu, Y.F.; Lu, G.Y.; Zhang, X.K.; Xie, L.L.; Yuan, C.F.; Xu, B.B. Graphene oxide modulates root growth of Brassica napus L. and regulates ABA and IAA concentration. J. Plant Physiol. 2016, 193, 57–63. [Google Scholar] [CrossRef]
- Naghizadeh, M.; Kabiri, R.; Hatami, A.; Oloumi, H.; Nasibi, F.; Tahmasei, Z. Exogenous application of melatonin mitigates the adverse effects of drought stress on morpho-physiological traits and secondary metabolites in Moldavian balm (Dracocephalum moldavica). Physiol. Mol. Biol. Plants 2019, 25, 881–894. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Kang, S.C. Free radical scavenging activity from different extracts of leaves of Bauhinia vahlii Wight &Arn. Saudi J. Biol. Sci. 2013, 20, 319–325. [Google Scholar]
- González-García, Y.; López-Vargas, E.R.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; González-Morales, S.; Robledo-Olivo, A.; Alpuche-Solís, G.; Juárez-Maldonado, A. Impact of carbon nanomaterials on the antioxidants system of tomato seedlings. Int. J. Mol. Sci. 2019, 20, 5858. [Google Scholar] [CrossRef]
- Ashraf, M.F.M.R.; Foolad, M.R. Role of glycinebetaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Hasanloo, T.; Mohammadi, S. Physiological characteristics, antioxidant enzyme activities, and gene expression in 2 spring canola (Brassica napus L.) cultivars under drought stress conditions. Turk. J. Agric. For. 2014, 38, 413–420. [Google Scholar] [CrossRef]
- Debnath, M. Responses of Bacopa monnieri to salinity and drought stress in vitro. J. Med. Plants Res. 2008, 11, 347–351. [Google Scholar]
- Hatami, M.; Hadian, J.; Ghorbanpour, M. Mechanisms underlying toxicity and stimulatory role of single-walled carbon nanotubes in Hyoscyamus niger during drought stress simulated by polyethylene glycol. J. Hazard. Mater. 2017, 324, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Sadak, M.S.; El-Enany MA, M.; Bakry, B.A.; Shater Abdallah, M.M.; El-Bassiouny, H.M.S. Signal molecules improving growth, yield and biochemical aspects of wheat cultivars under water stress. Asian J. Plant Sci. 2020, 19, 35–53. [Google Scholar] [CrossRef]
- Amanullah, K. Phosphorus and compost management influence maize (Zea mays) productivity under semiarid condition with and without phosphate solubilizing bacteria. Front. Plant Sci. 2015, 6, 1083. [Google Scholar]
- Jabeen, M.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Thiamin stimulates growth and secondary metabolites in turnip (Brassica rapa L.) leaf and root under drought stress. Physiol. Plant. 2020, 172, 1399–1411. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahbaz, M.; Ali, Q. Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pak. J. Bot. 2013, 45, 93–98. [Google Scholar]
- Ali, A.; Alqurainy, F. Activities of antioxidants in plants under environmental stress. In The Lutein-Prevention and Treatment for Diseases; Motohashi, N., Ed.; Trans-World Research Network: Trivandrum, India, 2006; pp. 187–256. [Google Scholar]
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 2019, 667, 485–499. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Hadian, J. Multi-walled carbon nanotubes stimulate callus induction, secondary metabolites biosynthesis and antioxidant capacity in medicinal plant Satureja khuzestanica in vitro. Carbon 2015, 94, 749–759. [Google Scholar] [CrossRef]
- Joshi, A.; Kaur, S.; Dharamvir, K.; Nayyar, H.; Verma, G. Multi-walled carbon nanotubes applied through seed-proiming influence early germination, root hair, growth and yield of bread wheat (Triticum aestivum L.). J. Sci. Food Agric. 2018, 98, 3148–3160. [Google Scholar] [CrossRef]
- Ramadan, E.; Freeg, H.A.; Shalaby, N.; Rizk, M.S.; Ma, J.; Du, W.; Ibrahim, O.M.; Alwutayd, K.M.; AbdElgawad, H.; Jo, I.; et al. Response of nine triticale genotypes to different salt concentrations at the germination and early seedling stages. PeerJ 2023, 11, e16256. [Google Scholar] [CrossRef]
- Patel, A.; Tiwari, S.; Parihar, P.; Singh, R.; Prasad, S.M. Carbon nanotubes as plant growth regulators: Impacts on growth reproductive system and soil microbial community. In Nanomaterials in Plants, Algae and Microorganisms; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2020; Volume 2, p. 2342. [Google Scholar]
- Singh, S.; Sinha, S. Accumulation of metals and its effects in Brassica juncea (L.) Czern. (cv. Rohini) grown on various amendments of tannery waste. Ecotoxicol. Environ. Saf. 2005, 62, 118–127. [Google Scholar] [CrossRef]
- Baranska, M.; Schütze, W.; Schulz, H. Determination of lycopene and B-carotene content in tomato fruits and related products: Comparison of FT-Raman, ATR-IR, and NIR Spectroscopy. Anal. Chem. 2006, 78, 8456–8461. [Google Scholar] [CrossRef]
- Britton, G. UV/visible spectroscopy. In Carotenoids: Spectroscopy; Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 1995; Volume 1B, pp. 13–62. [Google Scholar]
- Astorg, P.; Gradelet, S.; Bergès, R.; Suschetet, M. Dietary lycopene decreases the initiation of liver preneoplasticfoci by diethylnitrosamine in the rat. Nutr. Cancer 1997, 29, 60–68. [Google Scholar] [CrossRef]
- Mascio, P.D.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid single oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Dorais, M.; Turcotie, G.; Papadopoulos, A.P.; Hao, X.; Gosselin. Control of tomato fruit quality and flavor by EC and water management. Agric. Agri-Food Can. Rep. 2000, 18–21. [Google Scholar]
- Ali, H.E.M.; Ismail, G.S.M. Tomato fruit quality as influenced by salinity and nitric oxide. Turk. J. Bot. 2014, 38, 122–129. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Klapaki, G. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Hortic. 2000, 86, 247–260. [Google Scholar] [CrossRef]
- Dumas, Y.; Dadomo, M.; Lucca, G.D.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidant content of tomatoes. J. Sci. Food Agr. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Ann, B.M.; Devesh, S.; Gothandam, K.M. Effect of salt stress on expression of carotenoid pathway genes in tomato. J. Stress Physiol. Biochem. 2011, 7, 87–94. [Google Scholar]
- Dehkourdi, E.H.; Mosavi, M. Effect of anatase nano-particles (TiO2) on parsley seed germination (Petroselinum crispum) in vitro. Biol. Trace Elem. Res. 2013, 155, 283–286. [Google Scholar] [CrossRef]
- Patel, A.; Tiwari, S.; Parihar, P.; Singh, R.; Prasad, S.M. Carbon nanotubes as plant growth regulators. In Carbon Nanotubes as Plant Growth Regulators; Patel, A.S., Tiwarl, P., Parlhar, R., Singh, S., Parsad, M., Eds.; Elsevier: London, UK, 2019; pp. 23–42. [Google Scholar]
- Naudts, K.; Van Den Berge, J.; Farfan, E.; Rose, P.; Abdelgawad, H.; Ceulemans, R.; Janssens, I.A.; Asard, H.; Nijs, I. Future climate alleviates stress impact on grassland productivity through altered antioxidant capacity. Environ. Exp. Bot. 2014, 99, 150–158. [Google Scholar] [CrossRef]
- Saleh, A.M.; Abdel-Mawgoud, M.; Hassan, A.R.; Habeeb, T.H.; Yehia, R.S.; AbdElgawad, H. Global metabolic changes induced by arbuscular mycorrhizal fungi in oregano plants grown under ambient and elevated levels of atmospheric CO2. Plant Physiol. Biochem. 2020, 151, 255–263. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WIR | CNTs (mg/L) | Shoot Length (cm) | Branch no./Plant | Leaf no./Plant | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Length (cm) | Root Fresh Weight (g) |
|---|---|---|---|---|---|---|---|---|
| 100% | 0 | 22.7 d | 9.56 d | 34.44 cd | 56.42 d | 14.79 e | 10.05 c | 0.78 e |
| 20 | 37.76 b | 12.19 b | 38.24 c | 83.98 b | 32.63 b | 11.13 bc | 1.25 de | |
| 40 | 42.2 a | 20.35 a | 76.36 a | 131.33 a | 51.53 a | 11.09 bc | 1.58 d | |
| 75% | 0 | 22.15 de | 7.04 e | 21.49 f | 32.51 e | 13.43 e | 12.08 ab | 1.29 de |
| 20 | 32.43 c | 11.36 bc | 36.53 c | 78.93 b | 30.26 b | 13.33 a | 3.32 c | |
| 40 | 35 bc | 10.18 cd | 64.7 b | 64.72 c | 32.72 b | 12.73 a | 3.63 c | |
| 50% | 0 | 18.53 e | 6.21 e | 15.05 g | 24.55 f | 13.07 e | 12.74 a | 3.36 c |
| 20 | 21.77 de | 6.58 e | 24.07 ef | 51.22 d | 19.26 d | 13 a | 6.34 b | |
| 40 | 21.37 de | 9.92 d | 28.83 de | 56.33 d | 22.36 c | 13.38 a | 7.15 a |
| WIR | CNTs (mg/L) | TSS (mg/100 g Dry wt) | Proline (mg/100 g Dry wt) | FAA (mg/100 g Dry wt) |
|---|---|---|---|---|
| 100% | 0 | 1323.28 b | 35.41 g | 230.51 e |
| 20 | 1464.76 ab | 42.45 fg | 241.05 de | |
| 40 | 1517.5 a | 47.99 ef | 251.53 de | |
| 75% | 0 | 1554.11 a | 54.12 de | 268.48 cde |
| 20 | 1492.3 a | 59.26 cd | 279.86 bcd | |
| 40 | 1576.56 a | 64.4 bc | 298.61 bc | |
| 50% | 0 | 1453.75 ab | 64.87 bc | 302.06 bc |
| 20 | 1466.15 ab | 69.12 b | 318.86 ab | |
| 40 | 1491.66 a | 78.33 a | 357.89 a |
| WIR | CNTs (mg/L) | Plant Height (cm) | Branch no./Plant | Pod no./Plant | Plant Fresh wt. (g) | Pod wt./Plant (g) | Seed wt./Plant (g) |
|---|---|---|---|---|---|---|---|
| 100% | 0 | 42.09 d | 7.94 cd | 15.14 e | 105.91 e | 51.45 f | 22.26 e |
| 20 | 70.33 a | 11.41 b | 34.29 d | 310 c | 123.75 c | 61.88 b | |
| 40 | 70.02 a | 16.16 a | 42.13 a | 511.54 a | 179.6 a | 91.58 a | |
| 75% | 0 | 35.31 e | 6.87 d | 14.19 e | 77.29 f | 27.23 h | 18.41 e |
| 20 | 51.1 c | 7.49 d | 38.7 bc | 170.32 d | 84.01 e | 42.14 d | |
| 40 | 73.61 a | 12.68 b | 41.19 ab | 331.32 b | 134.42 b | 63.51 b | |
| 50% | 0 | 36.31 e | 6.7 d | 13.17 e | 62.26 f | 25.58 h | 13.75 f |
| 20 | 46.25 cd | 7.51 d | 15.34 e | 76.26 f | 37.74 g | 20.67 e | |
| 40 | 58.89 b | 9.28 c | 36.83 cd | 303.47 c | 97.53 d | 49.72 c |
| WIR | CNTs (mg/L) | Pod Yield (kg/Fed) | Seed Yield (kg/Fed) | Oil Yield (kg/Fed) | Protein Yield (kg/Fed) |
|---|---|---|---|---|---|
| 100% | 0 | 1064.52 d | 477.15 d | 209.09 ef | 86.79 de |
| 20 | 2088.54 a | 1158.5 b | 553.52 b | 231.91 b | |
| 40 | 2211.67 a | 2006.35 a | 952.25 a | 396.64 a | |
| 75% | 0 | 881.7 d | 397.52 d | 173.41 fg | 65.27 e |
| 20 | 1441.68 c | 940.08 bc | 435.48 d | 160.5 c | |
| 40 | 1782.64 b | 1139.53 b | 496.79 c | 213.84 b | |
| 50% | 0 | 496.41 e | 368.62 d | 152.71 g | 60.61 e |
| 20 | 979.24 d | 566.22 d | 256.49 e | 97.43 d | |
| 40 | 1373.24 c | 886.38 c | 433.25 d | 160.71 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakry, B.A.; Sadak, M.S.; Al Ashkar, N.M.; Ibrahim, O.M.; Okla, M.K.; El-Tahan, A.M. The Role of Carbon Nanotubes in Improving Drought Tolerance via Upregulation of the Physiological Processes of Peanut Plants Grown in Sandy Soils. Agronomy 2024, 14, 611. https://doi.org/10.3390/agronomy14030611
Bakry BA, Sadak MS, Al Ashkar NM, Ibrahim OM, Okla MK, El-Tahan AM. The Role of Carbon Nanotubes in Improving Drought Tolerance via Upregulation of the Physiological Processes of Peanut Plants Grown in Sandy Soils. Agronomy. 2024; 14(3):611. https://doi.org/10.3390/agronomy14030611
Chicago/Turabian StyleBakry, Bakry A., Mervat Sh. Sadak, Nagla M. Al Ashkar, Omar M. Ibrahim, Mohammad K. Okla, and Amira M. El-Tahan. 2024. "The Role of Carbon Nanotubes in Improving Drought Tolerance via Upregulation of the Physiological Processes of Peanut Plants Grown in Sandy Soils" Agronomy 14, no. 3: 611. https://doi.org/10.3390/agronomy14030611
APA StyleBakry, B. A., Sadak, M. S., Al Ashkar, N. M., Ibrahim, O. M., Okla, M. K., & El-Tahan, A. M. (2024). The Role of Carbon Nanotubes in Improving Drought Tolerance via Upregulation of the Physiological Processes of Peanut Plants Grown in Sandy Soils. Agronomy, 14(3), 611. https://doi.org/10.3390/agronomy14030611