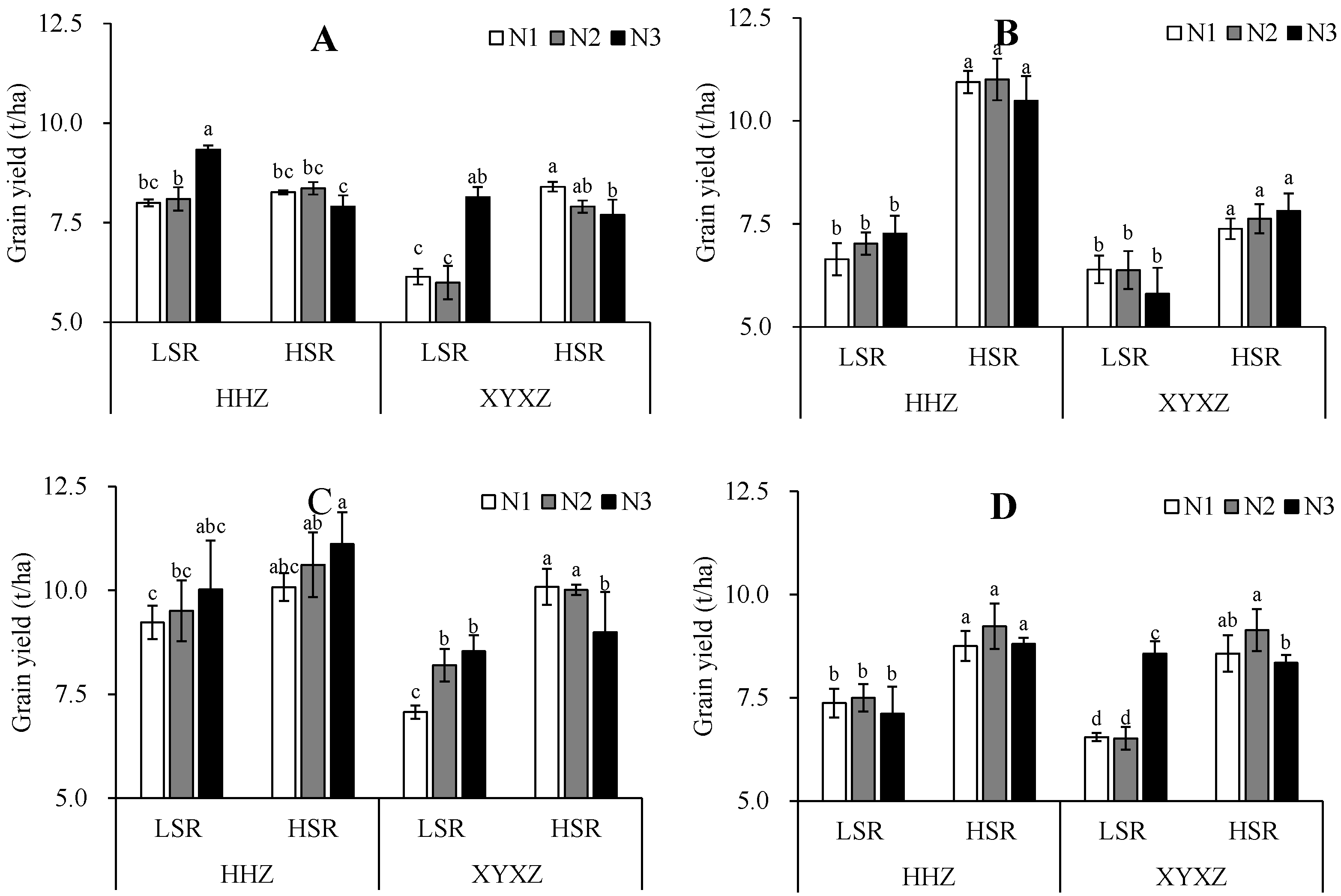
3.1. Grain Yield
Grain yield was influenced by SR and N fertilization rate (
Figure 2,
Table 1). In the late seasons of 2020 and 2021 and the early season in 2021, high seeding rate (HSR) resulted in yield that were 55.0%, 10.6%, and 22.0% higher than those of the low seeding rate (LSR) for HHZ, and these differences were statistically significant. A similar pattern was observed for XYXZ, where grain yields were 18.3%, 19.1%, 22.3%, and 28.2% higher for HSR than for LSR in both early and late seasons in 2020 and 2021. These differences were also statistically significant. Overall, increasing SR was associated with significant increases in yield for both cultivars in both years, though no significant differences were observed in the early season of 2020 for HHZ. Likewise, increasing N fertilization intensity increased grain yield for both cultivars in both years, though these differences were not statistically significant.
With the increase in nitrogen application at the LSR, the rice grain yield of HHZ showed an overall increase, although a fluctuating trend was observed in late season for 2021. In the case of XYXZ, there was a general increase in yield, with a minor decrease observed in late season for 2020 that was not statistically significant. With the increase in the nitrogen rate at the HSR, differences were observed between the two cultivars. The rice grain yield of HHZ showed an initial increase followed by a decrease except for an increase in early season of 2021. On the other hand, the rice grain yield of XYXZ displayed a decreasing trend in early season, an increasing trend in late season of 2020 and an initial increase followed by a decrease in late season of 2021.
The impact of the source of variation on grain yield was analyzed, with significant effects observed for SR and N (
Table 1). SR showed a significant effect on grain yield for both early and late season rice across two years. Additionally, there was a significant interaction effect between SR and N on grain yield, particularly in the early season of 2020 and late season of 2021.
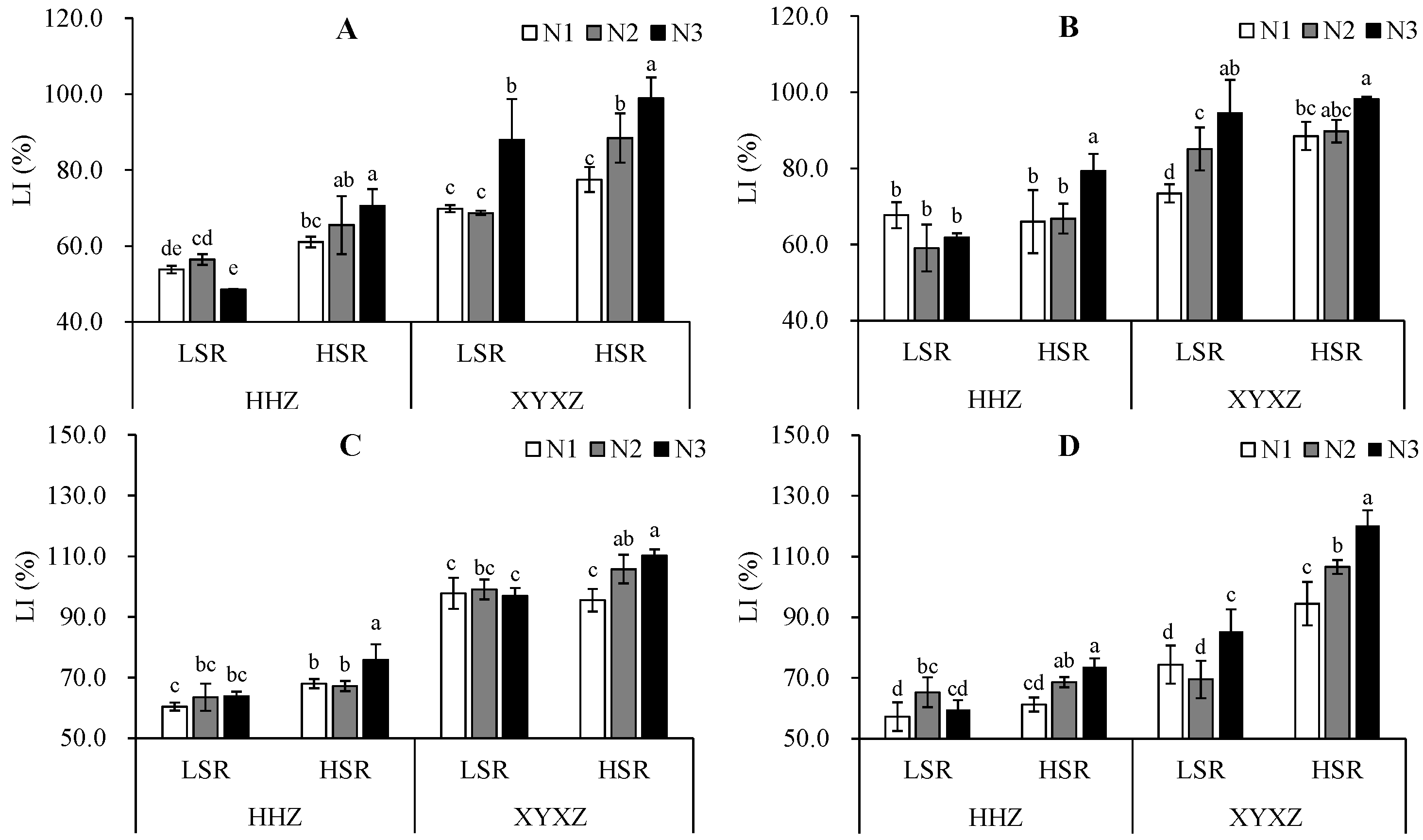
3.2. Lodging Index
Seeding rates had a significant impact on the LI, as shown in
Table 2 and
Table 3. In 2020, for early season, late season, and in 2021 for both early and late season, the HSR resulted in LI values that were 24.1%, 12.5%, 12.4% and 11.8% higher than those obtained with the LSR for HHZ. A similar pattern was observed in XYXZ, with the LI values under the HSR being 16.9%, 6.0% and 40.2% higher compared to those for LSR for early season of 2020, early season of 2021, and late season of 2021, respectively, and these differences were statistically significant. However, no significant difference was observed in late season of 2020.
Nitrogen fertilization increased the LI of both cultivars overall. The LI values were ranked as follows: N3 > N2 > N1, except for HHZ which showed N2 > N3 > N1 in early season of 2020. A slight decrease in the LI was noticed under N2 compared to N1 for HHZ in late season of 2020, with the ranking being N3 > N1 > N2. Additionally, the LI of HHZ with N2 and N3 in the late season of 2021, as well as the LI of XYXZ with N3 in both the early and late season of 2020, showed a significant increase compared to N1.
The different treatments had an impact on the LI. As shown in
Figure 3, it is evident that for HHZ, at the LSR, the LI initially increased and then decreased with the rising nitrogen application in the early season of 2020 and late season of 2021; conversely, an increasing trend was observed in the early season of 2021, while a reverse decreasing trend was observed in the late season of 2020. For XYXZ, at the LSR, the LI of N2 decreased compared to N1, but increased with N3 in the early season of 2020 and late season of 2021; an increasing trend was observed in the late season of 2020, while a pattern of an initial increasing and then decreasing trend was observed in early season of 2021. At the HSR, a trend of a consistent increasing trend in the LI was observed for both cultivars over the two years with an increase in nitrogen application.
SR, N and SR × N had effects on the LI, as shown in
Table 2 and
Table 3. The LI was significantly affected by the SR in both the early season and late season over two years, and the LI was significantly affected by N in the late season of 2021. Additionally, there were interaction effects of SR × N on the LI in the early season of 2020 and late season of 2021.
3.3. Lodging-Related Characteristics
The lodging-related characteristics, including SL, FW, WP, F and BM, were associated with the LI (
Table 2 and
Table 3). In both cultivars over two years, the HSR exhibited a higher SL compared to the LSR, with the differences in SL not being significant between the LSR and HSR of HHZ. However, for XYXZ, the SL was consistently higher than that of LSR by 2.6%, 6.6%, 1.0% and 7.8% with significant differences except for the early season in both years. Increased nitrogen application led to an increase in SL compared to the N1 treatment for both cultivars over two years except for HHZ in the late season of 2021. Furthermore, the SL of N3 was significantly higher than that of N1 for XYXZ in both the early and late seasons of 2020, and a significant increasing trend was observed for HHZ in the early season of 2021. Variance analysis revealed that SR and N had no significant effect on SL, and there were no significant interaction effects.
The HSR resulted in higher FW compared to the LSR for both cultivars over two years, except for the early season of XYXZ. For HHZ, FW was approximately 2.4%, 6.6%, 6.5% and 12.7% higher under the HSR than under the LSR in early season of 2020, late season of 2020, early season of 2021, and late season of 2021, respectively, with statistical significance except in 2020 early season. A similar increasing trend for XYXZ was only observed in the late season with a significant difference, while a decreasing trend was observed in the early season, with a significant decrease.
With the increase in nitrogen application, the FW of N2 and N3 showed an upward trend compared to N1 in both cultivars and years. In the case of HHZ, compared to N1, the FW of N2 and N3 saw a significant increase of 12.2%, 11.7%, 8.1% and 10.2% during the early season of 2020 and 2021, respectively. Moreover, a notable 8.8% difference was observed in the FW of N3 compared to N1 during the late season of 2020. Conversely, for XYXZ, the FW only showed a significant increase of 9.6% under N2 compared to N1 in the early season of 2020. Upon analyzing the sources of variation, it was found that FW was significantly influenced by N in all seasons except for the late season of 2020. Furthermore, in the early season of 2021, there was an interaction effect between SR and N on FW.
The HSR exhibited a higher WP compared to the LSR for HHZ in both years. However, there were variations in XYXZ, with WP decreasing in the early season but increasing in the late season. Furthermore, there was a significant decrease in WP in the early season of 2020. The differences in WP were particularly notable in the late season of 2021, with an increase of 14.0% and 31.6% for HHZ and XYXZ, respectively.
The WP was increasing with nitrogen application. In the case of HHZ, the WP of N2 and N3 significantly increased compared to N1, with N3 showing a significant increasing trend in the late season of 2020 and early season of 2021. On the other hand, for XYXZ, the WP of N2 and N3 only showed a significant increase in the early season of 2020 when compared to N1. An analysis of the sources of variation revealed that both SR and N had a significant effect on WP across both cultivars and years, except for SR in 2020. Additionally, there was a significant interaction between SR and N on WP, except in the late season of 2021.
Overall, F was lower for the HSR compared to the LSR across two cultivars and years. Specifically, for the HHZ cultivar, the F was lower under the HSR than under the LSR in both early and late season of 2020, and early season of 2021. However, a slight increase was observed in the late season of 2021, although it was not statistically significant.
There was a significant decrease in XYXZ during the early season of 2020. When compared with N1, the application of nitrogen at levels of N2 and N3 showed no significant difference in two cultivars over the two years. Analyzing the source of variation, it was found that SR had a significant effect on F during the early season of 2020.
In both two cultivars and years, the HSR resulted in a lower BM compared to the LSR. Specifically, for the HHZ cultivar, the BM under the HSR was consistently lower than that under the LSR, except for the late season in 2021. Furthermore, the BM under the HSR was significantly lower than that in the early season of 2020. On the other hand, for the XYXZ cultivar, the BM under the HSR was lower than that under the LSR only in the early season of 2020, with no significant differences observed in other seasons.
As demonstrated in
Table 2 and
Table 3, nitrogen application has an impact on BM. In the case of HHZ, an increasing in nitrogen application led to a corresponding increasing in BM in both seasons of 2020, although the difference was not statistically significant. Furthermore, a trend of an initial increasing followed by a decreasing trend was observed in both seasons of 2021, without significant differences. Conversely, for XYXZ, there was no significant effect on BM with the increment in nitrogen. In an analysis of the sources of variation, it was found that SR had a significant effect on BM in the early season of 2020.
3.4. Culm Characteristics
The PH and GCH were increased under the HSR compared to the LSR, with no significant differences found in the PH and GCH of HHZ. However, there were significant differences in XYXZ in the late season of both years, and the PH of XYXZ was also significantly increased in the early season of 2020.
The PH and GCH were positively influenced by higher nitrogen rates. In the early season of 2021, there was a significant increment of PH for HHZ with N2 and N3 compared to N1, whereas significant differences were only seen in the early season of 2020 for XYXZ. Additionally, the GCH of N3 showed a significant increased only in the early season of 2020 for HHZ compared to N1, while significant differences were observed in the late season of 2020 and early season of 2021 for XYXZ.
The CD and CWT exhibited a decreasing trend under the HSR compared to the LSR, as shown in
Table 4 and
Table 5. While CD was significantly reduced only in the 2020 early season of HHZ, the sole significant difference was observed in the 2020 late season of XYXZ. No significant differences were observed for CWT.
In the late season of 2020, the CD of HHZ significantly decreased with the increasing of nitrogen application from N1 to N3. For XYXZ, the CD increased in the early season and then showed an initial increasing followed by a decreasing trend in the late season for both years. No significant differences were observed in the CD of both cultivars over the two years.
With the nitrogen application increased, no significant differences were observed in the CWT of HHZ between the two cultivars and years. In the case of XYXZ, the CWT decreased in the early season of 2020 and the late season of 2021, while it initially increased and then decreased in the late season of 2020 and the early season of 2021. However, a significant reduction was only observed with N3 compared to N1 in the late season of 2020.
The length of the basal internodes, such as the first (1st) basal internode and the second (2nd) basal internode, were higher under the HSR compared to the LSR for two cultivars and years. There were no significant differences were observed for HHZ. Significant difference was only observed in the 2021 late season of XYXZ. Overall, with the nitrogen application increased, the length of the first and second internodes were increased. In comparison to N1, the first internode of N3 was significantly higher for HHZ, except in the early season of 2020, and no significant difference was found for the second internode.
An analysis of the sources of variation in culm characteristics revealed that there was no significant difference observed in the PH. The GCH was significantly affected by SR in both seasons of 2020 and in the late season of 2021, as well as by N in both seasons of 2020. No interaction effects were observed for GCH. CD was significantly affected by SR and SR × N in the early season of 2020. Similarly, CWT was significantly affected by SR and SR × N in the early season of 2020 and by N in the late season of 2020. The basal first internode was significantly affected by SR in the early season of 2020 and by N in the late season of 2020. The basal second internode was significantly affected by SR in the early season of 2020 and in the late season of 2021.



{kind=link}
{kind=link}
{kind=link}