
Genome-Wide Exploration of the WD40 Gene Family in Eggplant (Solanum melongena L.) and Analysis of Its Function in Fruit Color Formation

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Physicochemical Property of SmWD40 Gene Family
2.2. Chromosomal Location and Phylogenetic Analysis of SmWD40
2.3. Conserved Motif Analysis and Cis-Acting Element Analysis of SmWD40
2.4. Collinearity Analysis of SmWD40
2.5. Plant Materials and Treatments
2.6. Virus-Induced Gene Silencing
2.7. Analysis of Anthocyanin Synthesis Structural Gene Expression Level
2.8. qRT-PCR and Viral Vector Construction
2.9. Interaction Network Analysis of SmWD40-56
3. Results
3.1. Identification of the WD40 Genes in Eggplant
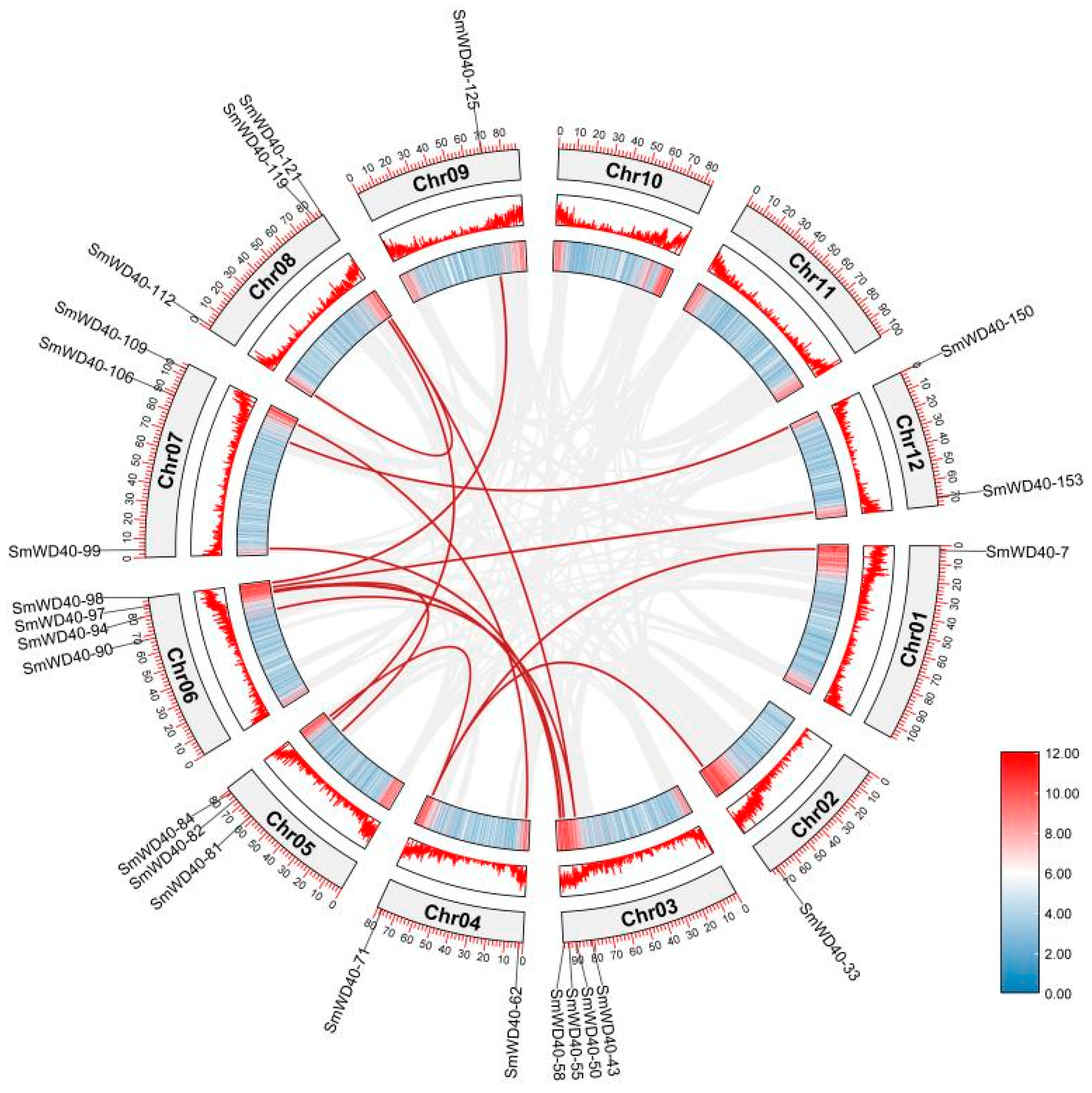
3.2. Chromosome Localization of SmWD40 Genes
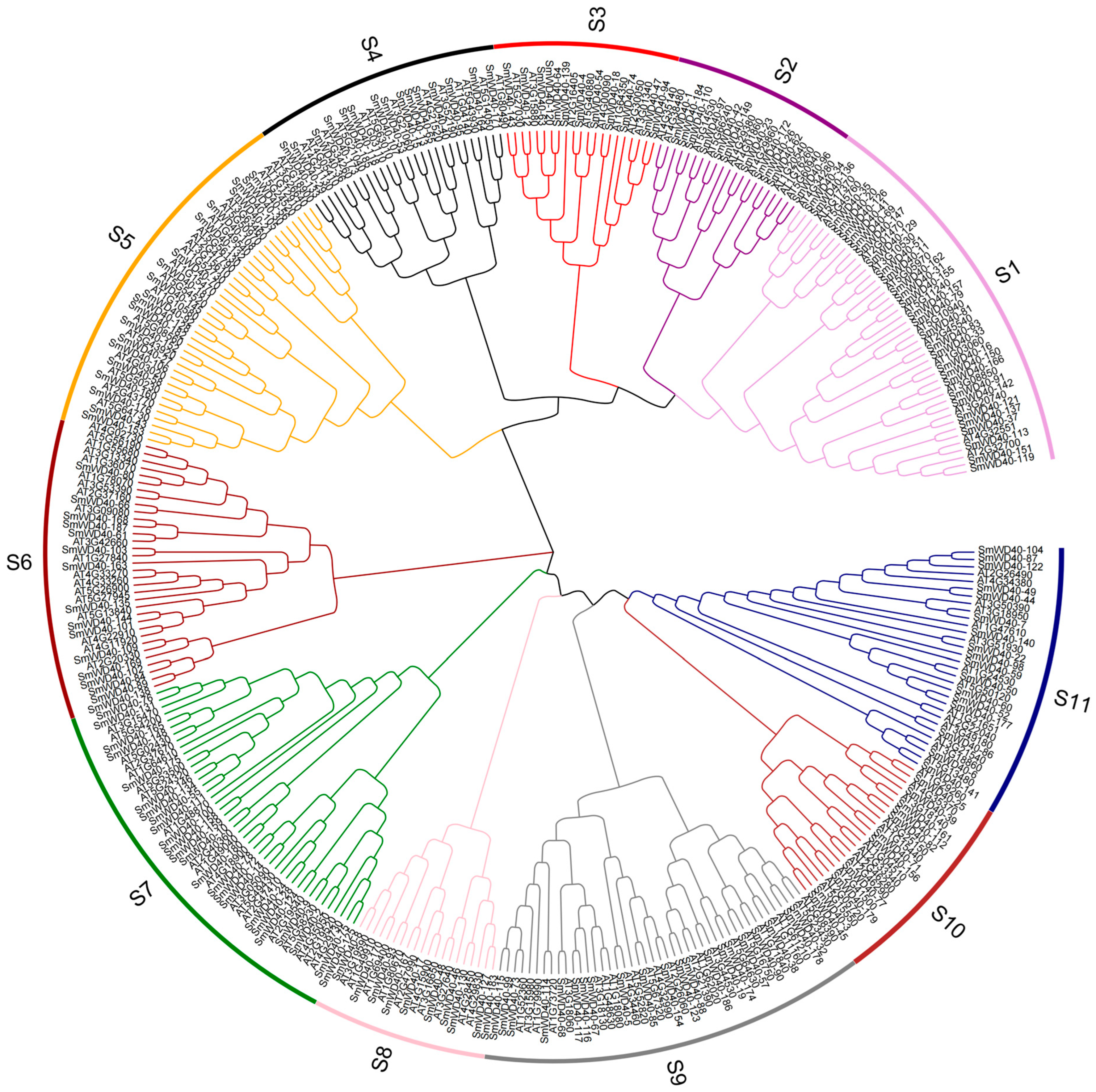
3.3. Phylogenetic Analysis of SmWD40 Proteins
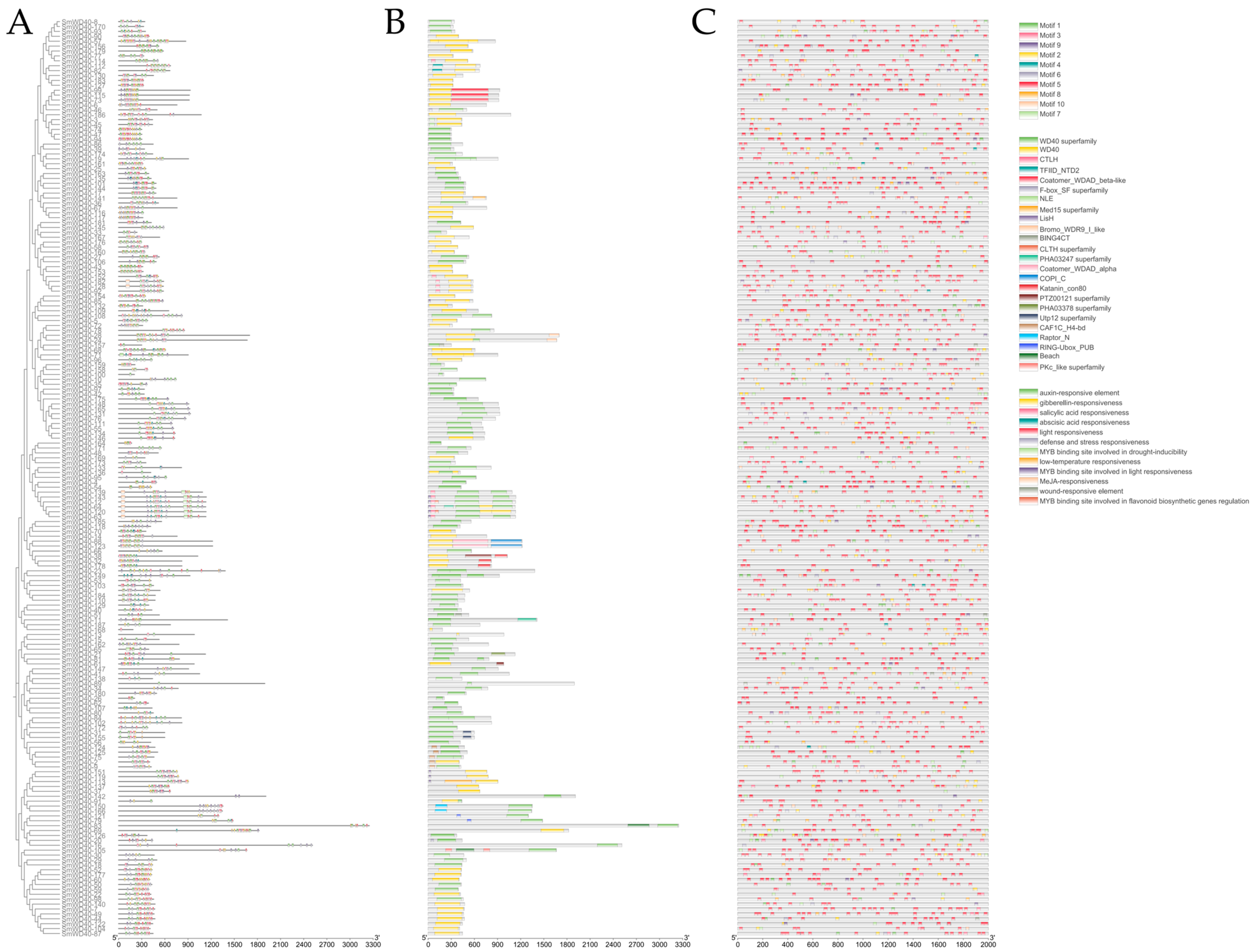
3.4. Conserved Domains and Cis-Acting Elements Prediction of SmWD40s
3.5. Collinearity Analysis of SmWD40 Genes
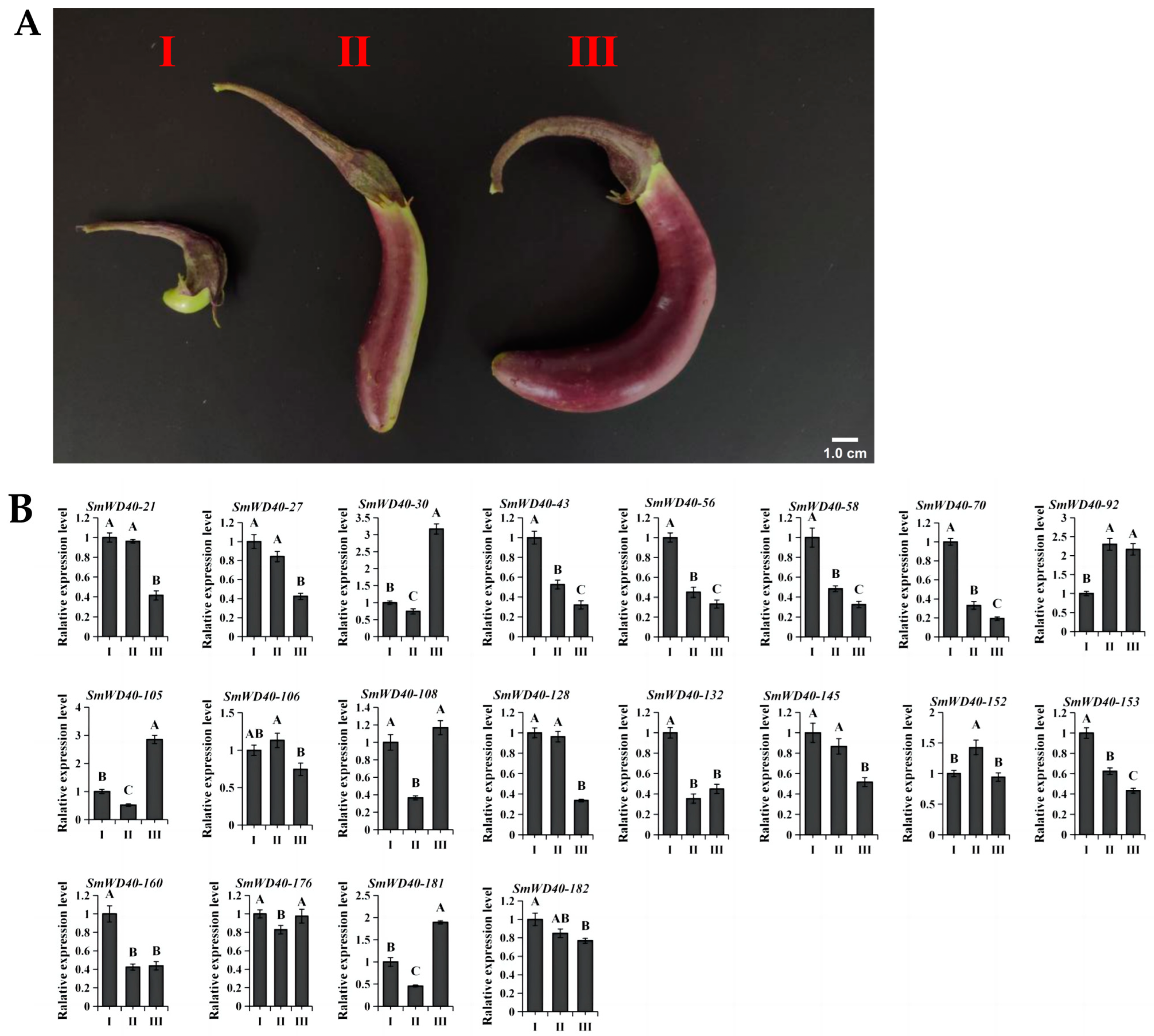
3.6. Expression Pattern Analysis of S5 Subgroup Genes in Fruit Color Development
3.7. Analysis of the Effects of Silencing SmWD40-56 on Fruit Color and the Expression Pattern of Structural Genes for Anthocyanin Synthesis after Silencing
3.8. Protein Interaction Network of SmWD40-56 in Eggplant
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene ID | Forward Primer (5′->3′) | Reverse Primer (5′->3′) | Function | Amplification Efficiency |
|---|---|---|---|---|---|
| SmWD40-21 | Smechr0102060.1 | GGATGGGGATGCAAGGACAA | CATGCATCAAGTTCTCGCCG | SmWD40-21 qRT-PCR | 0.9612 |
| SmWD40-27 | Smechr0102379.1 | TGGAATAGGTGTGGGGGACT | TGTGCATAACCACCAAACTGG | SmWD40-27 qRT-PCR | 0.9956 |
| SmWD40-30 | Smechr0200037.1 | AGAGGAATCTGGGGTTGGGA | AGCCAAACGGTAAGGGACAG | SmWD40-30 qRT-PCR | 0.9835 |
| SmWD40-43 | Smechr0202538.1 | ATGACTTCACCTACCGAGCC | AGAAGGGATCCATCGGAAACG | SmWD40-43 qRT-PCR | 0.9744 |
| SmWD40-56 | Smechr0302292.1 | TGAGTGCCGAAAATAACAATAAAAC | GGGCTATCAATTGGGTTTCGAC | SmWD40-56 qRT-PCR | 0.9725 |
| SmWD40-58 | Smechr0302644.1 | ACAGTCATGTTACCGCAGGTT | AGGTAGCCAGAGGTTGACCA | SmWD40-58 qRT-PCR | 0.9882 |
| SmWD40-70 | Smechr0303426.1 | GCTGGGACCTCTCGGTCA | ACCATCGGCATGTGTCTCTT | SmWD40-70 qRT-PCR | 0.9673 |
| SmWD40-92 | Smechr0500827.1 | AGCAGGTTCTAGCTGGGATG | AGGGCCTGTTCAACGAGATG | SmWD40-92 qRT-PCR | 0.9956 |
| SmWD40-105 | Smechr0502667.1 | TGATATTTCATCCCTGCACGA | AGGCCTTTTCCAGGTGAGTC | SmWD40-105 qRT-PCR | 1.0014 |
| SmWD40-106 | Smechr0600010.1 | CATCGGTCGACCATTCGAGA | TTTGGAGGGAGTGGTGCTTC | SmWD40-106 qRT-PCR | 0.9892 |
| SmWD40-108 | Smechr0600126.1 | TTTGCTCCTTGGCACAACAA | TGCAACTTTCTCGTCTGCCC | SmWD40-108 qRT-PCR | 0.9766 |
| SmWD40-128 | Smechr0701564.1 | TGCAAAAGATTGGTCTGGTGG | TGTTTATGTAAAGAGGCGCGA | SmWD40-128 qRT-PCR | 0.9883 |
| SmWD40-132 | Smechr0702707.1 | ATGTCGTGCTGCCATGAACA | GCTCTCAAAGCCAAATACACG | SmWD40-132 qRT-PCR | 0.9965 |
| SmWD40-145 | Smechr0802330.1 | AGAGTACGATCAACATCTCGGA | GGCGCAATAGATGGCATGG | SmWD40-145 qRT-PCR | 0.9746 |
| SmWD40-152 | Smechr0901596.1 | GTGCAGACCCTGAAAAAGGC | CACCACCTGTCCCCATGTTG | SmWD40-152 qRT-PCR | 1.0233 |
| SmWD40-153 | Smechr0901745.1 | CGGGGACATACTGGCTTTGT | TCACCGGCATAGAATGAGCC | SmWD40-153 qRT-PCR | 0.9516 |
| SmWD40-160 | Smechr1000926.1 | CCCCCTCTTATTGTCAGCGG | GAGGCGTCAGAGAAACAGACA | SmWD40-160 qRT-PCR | 0.9777 |
| SmWD40-176 | Smechr1100624.1 | ATGACCGCACAACTGCCAAA | TAGCCGGATGGTCCGATCTT | SmWD40-176 qRT-PCR | 0.1023 |
| SmWD40-181 | Smechr1102707.1 | AACAACCCTTTGGTCCCAGC | GTTAGAAGCAGCCGTTGTGC | SmWD40-181 qRT-PCR | 0.9761 |
| SmWD40-182 | Smechr1200022.1 | CAGGTCATAACGGCCCTGTG | TGGAGCCCATTCACACGAAA | SmWD40-182 qRT-PCR | 0.9552 |
| SmCHI | Smechr0500261.1 | CGATCGCTTAGCAGAAGCCG | GCGAACGATATCTTTGCACTACT | SmCHI qRT-PCR | 0.9765 |
| SmCHS | Smechr0500387.1 | GCTCGTCGTCTGTGCTGAG | ATCCCCGTTAGGCACGAATG | SmCHS qRT-PCR | 0.9838 |
| SmF3H | Smechr0101736.1 | AGCAATGGGATATACAAAGGCATC | CGCTAACGCAGGTCCAACT | SmF3H qRT-PCR | 0.9689 |
| SmDFR | Smechr0202337.1 | AGTCAGGGGAATGTTGAGCA | TCCAGCTAGTCTCGTCATAGA | SmDFR qRT-PCR | 1.0332 |
| SmFLS | Smechr0401078.1 | AAGGTCAGAGCACAACAGCC | GTCGATGACTGGCACCTCAA | SmFLS qRT-PCR | 0.9777 |
| SmPAL | Smechr0400589.1 | TGGAACTTTCGAGGGCGAAT | CGTCGGGCGATCGTACTTTT | SmPAL qRT-PCR | 0.9923 |
| SmActin | Smechr1001310.1 | GGTCGGAATGGGACAGAAGG | GGTGCCTCAGTCAGGAGAAC | Reference Gene | 0.9936 |
| SmWD40-56-VIGS | Smechr0302292.1 | ctccatggggatccgcCCAATGACATTCTTG | agacgcgtgagctcgATCAGCAGAAACGGAAG | SmWD40-56 amplification of silencing fragments | |
| TRV2 | none | TGGGAGATGATACGCTGTT | CCTAAAACTTCAGACACG | Detect infected plants |
| Gene Name | Gene ID | Number of Amino Acids | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity |
|---|---|---|---|---|---|---|---|
| SmWD40-1 | Smechr0100054.1 | 538 | 60030.64 | 4.34 | 34.27 | 70.46 | −0.589 |
| SmWD40-2 | Smechr0100084.1 | 403 | 45571.6 | 4.69 | 40.3 | 77.59 | −0.516 |
| SmWD40-3 | Smechr0100138.1 | 440 | 48400.93 | 5.45 | 36.58 | 87.09 | −0.189 |
| SmWD40-4 | Smechr0100203.1 | 493 | 55068.17 | 9.04 | 48.05 | 72.17 | −0.398 |
| SmWD40-5 | Smechr0100250.1 | 377 | 40950.02 | 7.19 | 29.56 | 76.47 | −0.233 |
| SmWD40-6 | Smechr0100269.1 | 757 | 82965.53 | 5.51 | 32.25 | 94.66 | −0.151 |
| SmWD40-7 | Smechr0100290.1 | 469 | 51874.12 | 8.72 | 41.8 | 76.46 | −0.486 |
| SmWD40-8 | Smechr0100456.1 | 340 | 37891.44 | 8.51 | 35.37 | 68.5 | −0.521 |
| SmWD40-9 | Smechr0100664.1 | 424 | 48291.91 | 4.7 | 46.24 | 78.4 | −0.548 |
| SmWD40-10 | Smechr0100732.1 | 747 | 84865.95 | 5.11 | 51.97 | 73.31 | −0.732 |
| SmWD40-11 | Smechr0100835.1 | 1412 | 150253.47 | 6.92 | 39.42 | 88.53 | −0.101 |
| SmWD40-12 | Smechr0100970.1 | 1132 | 124636.89 | 6.6 | 42.72 | 84.89 | −0.214 |
| SmWD40-13 | Smechr0101114.1 | 982 | 106620.95 | 5.42 | 52.75 | 75.51 | −0.339 |
| SmWD40-14 | Smechr0101146.1 | 759 | 83050.86 | 6.88 | 34.38 | 82.71 | −0.321 |
| SmWD40-15 | Smechr0101232.1 | 526 | 57618.08 | 4.84 | 35.11 | 87.05 | −0.282 |
| SmWD40-16 | Smechr0101322.1 | 2513 | 276904.98 | 6.3 | 42.12 | 86.42 | −0.061 |
| SmWD40-17 | Smechr0101516.1 | 484 | 53493.77 | 9.17 | 39.41 | 72.77 | −0.477 |
| SmWD40-18 | Smechr0101622.1 | 441 | 47647.5 | 8.92 | 32.22 | 86.73 | −0.199 |
| SmWD40-19 | Smechr0102034.1 | 907 | 101447.44 | 8.01 | 42.57 | 93.84 | −0.141 |
| SmWD40-20 | Smechr0102054.1 | 442 | 49465.81 | 9.04 | 37.15 | 82.62 | −0.238 |
| SmWD40-21 | Smechr0102060.1 | 1126 | 121602.13 | 5.23 | 49.64 | 78.53 | −0.288 |
| SmWD40-22 | Smechr0102098.1 | 419 | 46256.05 | 9.03 | 40.99 | 84.39 | −0.19 |
| SmWD40-23 | Smechr0102184.1 | 1382 | 154085.28 | 6.18 | 35.94 | 86.11 | −0.15 |
| SmWD40-24 | Smechr0102204.1 | 1667 | 186041.55 | 6.55 | 46.43 | 73.11 | −0.597 |
| SmWD40-25 | Smechr0102232.1 | 353 | 39736.77 | 5.72 | 35.53 | 75.38 | −0.292 |
| SmWD40-26 | Smechr0102378.1 | 208 | 23235.25 | 6.85 | 52.26 | 73.94 | −0.32 |
| SmWD40-27 | Smechr0102379.1 | 528 | 59948.58 | 9.06 | 50.64 | 73.86 | −0.476 |
| SmWD40-28 | Smechr0102778.1 | 1698 | 190580.35 | 6.39 | 50.3 | 67.02 | −0.738 |
| SmWD40-29 | Smechr0103230.1 | 496 | 55498.41 | 8.63 | 47.27 | 76.05 | −0.406 |
| SmWD40-30 | Smechr0200037.1 | 462 | 52158.93 | 8.17 | 30.21 | 90.65 | −0.177 |
| SmWD40-31 | Smechr0200130.1 | 598 | 65181.09 | 6.65 | 41.09 | 91.79 | −0.08 |
| SmWD40-32 | Smechr0200257.1 | 820 | 90126.36 | 8.35 | 38.2 | 87.24 | −0.286 |
| SmWD40-33 | Smechr0200514.1 | 239 | 26251.65 | 4.69 | 24.93 | 101.17 | −0.029 |
| SmWD40-34 | Smechr0200897.1 | 773 | 86581.58 | 7.06 | 51.25 | 89.5 | −0.232 |
| SmWD40-35 | Smechr0201236.1 | 369 | 40099.45 | 7.04 | 24.63 | 79.7 | −0.189 |
| SmWD40-36 | Smechr0201464.1 | 516 | 58044.79 | 6.6 | 50.01 | 69.38 | −0.766 |
| SmWD40-37 | Smechr0201698.1 | 671 | 73205.32 | 5.75 | 44.53 | 79.66 | −0.261 |
| SmWD40-38 | Smechr0201774.1 | 1027 | 113880.83 | 6.41 | 55.77 | 80.68 | −0.452 |
| SmWD40-39 | Smechr0201777.1 | 336 | 36529.3 | 6.86 | 39.42 | 81.25 | −0.208 |
| SmWD40-40 | Smechr0201803.1 | 432 | 48118.81 | 4.86 | 47.8 | 58.5 | −0.526 |
| SmWD40-41 | Smechr0202123.1 | 1051 | 115331.91 | 6.84 | 42.3 | 92.12 | −0.186 |
| SmWD40-42 | Smechr0202247.1 | 332 | 36903.65 | 6.09 | 37.48 | 78.07 | −0.228 |
| SmWD40-43 | Smechr0202538.1 | 314 | 34577.84 | 6.36 | 22.95 | 80.99 | −0.291 |
| SmWD40-44 | Smechr0202756.1 | 471 | 52145.57 | 8.9 | 42.35 | 71 | −0.524 |
| SmWD40-45 | Smechr0202868.1 | 440 | 48281.7 | 5.53 | 36.32 | 87.55 | −0.189 |
| SmWD40-46 | Smechr0202967.1 | 499 | 54952.24 | 7.31 | 41.3 | 82.65 | −0.244 |
| SmWD40-47 | Smechr0203005.1 | 302 | 32550.54 | 5.84 | 22.74 | 79.11 | −0.216 |
| SmWD40-48 | Smechr0203082.1 | 514 | 56664.04 | 8.77 | 50.81 | 74.36 | −0.388 |
| SmWD40-49 | Smechr0300525.1 | 460 | 51307.37 | 8.6 | 40.56 | 70.93 | −0.491 |
| SmWD40-50 | Smechr0301092.1 | 432 | 47227.71 | 9.15 | 41.93 | 87.59 | −0.168 |
| SmWD40-51 | Smechr0301403.1 | 528 | 59266.6 | 9.56 | 36.37 | 78.52 | −0.52 |
| SmWD40-52 | Smechr0301606.1 | 434 | 47495.08 | 9.01 | 37.03 | 83.53 | −0.235 |
| SmWD40-53 | Smechr0301684.1 | 421 | 47215.62 | 7.94 | 42.57 | 82.16 | −0.144 |
| SmWD40-54 | Smechr0301928.1 | 430 | 47321.27 | 8.13 | 32.35 | 71.56 | −0.381 |
| SmWD40-55 | Smechr0302236.1 | 714 | 78581.52 | 6.43 | 46.39 | 75.84 | −0.369 |
| SmWD40-56 | Smechr0302292.1 | 417 | 47092.84 | 4.89 | 49.66 | 79.28 | −0.433 |
| SmWD40-57 | Smechr0302364.1 | 871 | 96184.7 | 6.05 | 41.83 | 87.3 | −0.181 |
| SmWD40-58 | Smechr0302644.1 | 385 | 42188.43 | 6.49 | 32.66 | 83.32 | −0.242 |
| SmWD40-59 | Smechr0302647.1 | 392 | 44101.14 | 7.17 | 43.02 | 89.97 | −0.243 |
| SmWD40-60 | Smechr0302855.1 | 404 | 44930.37 | 8.86 | 35.58 | 84.41 | −0.215 |
| SmWD40-61 | Smechr0302946.1 | 980 | 108146.29 | 5.79 | 47.3 | 79.45 | −0.408 |
| SmWD40-62 | Smechr0302999.1 | 387 | 42791.76 | 4.97 | 32.53 | 76.02 | −0.42 |
| SmWD40-63 | Smechr0303123.1 | 1132 | 124471.51 | 6.57 | 39.07 | 82.47 | −0.249 |
| SmWD40-64 | Smechr0303179.1 | 1130 | 124653.99 | 6.76 | 39.34 | 79.57 | −0.35 |
| SmWD40-65 | Smechr0303180.1 | 391 | 42538.27 | 5.22 | 52.86 | 89.05 | 0.017 |
| SmWD40-66 | Smechr0303262.1 | 564 | 63489.27 | 8.57 | 56 | 83.67 | −0.459 |
| SmWD40-67 | Smechr0303326.1 | 759 | 85606.12 | 6.63 | 39.59 | 94.18 | −0.162 |
| SmWD40-68 | Smechr0303331.1 | 609 | 66187.92 | 6.15 | 24.42 | 89.26 | −0.139 |
| SmWD40-69 | Smechr0303392.1 | 1821 | 202371.41 | 6.31 | 41.24 | 97.76 | −0.075 |
| SmWD40-70 | Smechr0303426.1 | 354 | 38347.79 | 5.4 | 38.02 | 74.92 | −0.329 |
| SmWD40-71 | Smechr0303432.1 | 903 | 101063.15 | 6.23 | 38.07 | 85.2 | −0.162 |
| SmWD40-72 | Smechr0303452.1 | 313 | 35957.79 | 5.13 | 42.84 | 85.59 | −0.286 |
| SmWD40-73 | Smechr0303453.1 | 916 | 103343.75 | 4.87 | 32.34 | 84.49 | −0.286 |
| SmWD40-74 | Smechr0303517.1 | 301 | 32399.3 | 5.09 | 32.56 | 81.96 | −0.158 |
| SmWD40-75 | Smechr0303588.1 | 459 | 50643.58 | 5.6 | 41.09 | 76.86 | −0.449 |
| SmWD40-76 | Smechr0400216.1 | 874 | 96741.3 | 5.58 | 47.68 | 70.55 | −0.581 |
| SmWD40-77 | Smechr0401256.1 | 326 | 35928.7 | 7.23 | 30.61 | 82.24 | −0.313 |
| SmWD40-78 | Smechr0401335.1 | 854 | 94768.78 | 5.31 | 39.39 | 94.67 | 0.025 |
| SmWD40-79 | Smechr0401384.1 | 1484 | 166473.85 | 5.68 | 51.06 | 89.97 | −0.285 |
| SmWD40-80 | Smechr0401851.1 | 450 | 50649.48 | 5 | 38.44 | 74.56 | −0.414 |
| SmWD40-81 | Smechr0401950.1 | 789 | 86855.86 | 6.27 | 51.99 | 79.15 | −0.397 |
| SmWD40-82 | Smechr0401952.1 | 665 | 73574.83 | 6.21 | 46.54 | 78.32 | −0.376 |
| SmWD40-83 | Smechr0401998.1 | 3249 | 364048.07 | 6.09 | 47.63 | 96 | −0.116 |
| SmWD40-84 | Smechr0402112.1 | 814 | 89327.63 | 6.31 | 39.76 | 84.9 | −0.191 |
| SmWD40-85 | Smechr0402166.1 | 580 | 65695.44 | 5.61 | 24.67 | 68.57 | −0.767 |
| SmWD40-86 | Smechr0402222.1 | 448 | 49231.7 | 6.03 | 48.3 | 89.44 | −0.212 |
| SmWD40-87 | Smechr0402308.1 | 447 | 49013.24 | 8.59 | 36.1 | 77.56 | −0.4 |
| SmWD40-88 | Smechr0402434.1 | 1218 | 137204.73 | 6.63 | 33.33 | 91.33 | −0.261 |
| SmWD40-89 | Smechr0500012.1 | 1895 | 211341.88 | 5.36 | 48.62 | 89.18 | −0.294 |
| SmWD40-90 | Smechr0500094.1 | 397 | 42640.62 | 4.3 | 47.4 | 75.42 | −0.271 |
| SmWD40-91 | Smechr0500782.1 | 438 | 49316.61 | 6.01 | 45 | 94.18 | −0.174 |
| SmWD40-92 | Smechr0500827.1 | 581 | 64645.73 | 5.75 | 42.19 | 93.87 | −0.199 |
| SmWD40-93 | Smechr0501136.1 | 348 | 38447.5 | 8.69 | 36.1 | 68.65 | −0.412 |
| SmWD40-94 | Smechr0501240.1 | 302 | 32668.9 | 5.9 | 29.18 | 83.54 | −0.135 |
| SmWD40-95 | Smechr0501281.1 | 623 | 69179.42 | 5.84 | 38.02 | 84.06 | −0.154 |
| SmWD40-96 | Smechr0501342.1 | 437 | 48304.69 | 8.37 | 35.31 | 82.59 | −0.288 |
| SmWD40-97 | Smechr0501500.1 | 334 | 36872.71 | 5.99 | 41.91 | 83.8 | −0.14 |
| SmWD40-98 | Smechr0501683.1 | 455 | 50098.93 | 8.06 | 44.26 | 82.68 | −0.168 |
| SmWD40-99 | Smechr0501744.1 | 929 | 104910.34 | 4.93 | 32.25 | 86.76 | −0.302 |
| SmWD40-100 | Smechr0501824.1 | 205 | 24016.19 | 8.46 | 49.64 | 62.73 | −1.172 |
| SmWD40-101 | Smechr0502137.1 | 486 | 53616.15 | 9.15 | 41.21 | 72.61 | −0.531 |
| SmWD40-102 | Smechr0502285.1 | 821 | 90061.98 | 7.33 | 38.18 | 85.7 | −0.162 |
| SmWD40-103 | Smechr0502290.1 | 453 | 49769.86 | 8.69 | 42.92 | 76.87 | −0.385 |
| SmWD40-104 | Smechr0502356.1 | 410 | 45405.3 | 8.34 | 30.5 | 74.85 | −0.383 |
| SmWD40-105 | Smechr0502667.1 | 1663 | 184193.33 | 6.19 | 51.02 | 89.99 | −0.146 |
| SmWD40-106 | Smechr0600010.1 | 487 | 55330.28 | 7.69 | 48.69 | 76.86 | −0.441 |
| SmWD40-107 | Smechr0600111.1 | 434 | 47259.87 | 5.42 | 41 | 91.68 | −0.058 |
| SmWD40-108 | Smechr0600126.1 | 826 | 91024.23 | 6.08 | 41.81 | 87.19 | −0.123 |
| SmWD40-109 | Smechr0600145.1 | 650 | 71617.27 | 5.45 | 47.01 | 70.52 | −0.661 |
| SmWD40-110 | Smechr0600493.1 | 473 | 53158.71 | 6.35 | 52.38 | 78.9 | −0.465 |
| SmWD40-111 | Smechr0601395.1 | 695 | 77092.81 | 5.98 | 44.39 | 83.41 | −0.31 |
| SmWD40-112 | Smechr0601940.1 | 673 | 74624.11 | 6.28 | 48.35 | 78.66 | −0.39 |
| SmWD40-113 | Smechr0601947.1 | 905 | 99310.05 | 6.42 | 54.43 | 65.31 | −0.713 |
| SmWD40-114 | Smechr0602039.1 | 514 | 58190.36 | 6.53 | 48.13 | 82.47 | −0.361 |
| SmWD40-115 | Smechr0602212.1 | 917 | 103580.79 | 4.89 | 31.65 | 86.65 | −0.284 |
| SmWD40-116 | Smechr0602338.1 | 323 | 35657.39 | 7.03 | 30.15 | 87.77 | −0.16 |
| SmWD40-117 | Smechr0602339.1 | 315 | 34997.6 | 6.65 | 33.71 | 86.6 | −0.194 |
| SmWD40-118 | Smechr0602689.1 | 416 | 47296.75 | 6.12 | 31.03 | 83.92 | −0.143 |
| SmWD40-119 | Smechr0603088.1 | 778 | 84819.8 | 6.43 | 49.78 | 67.98 | −0.539 |
| SmWD40-120 | Smechr0700242.1 | 1134 | 125041.54 | 6.46 | 37.69 | 75.56 | −0.343 |
| SmWD40-121 | Smechr0700811.1 | 1300 | 146022.8 | 5.8 | 43.64 | 99 | −0.109 |
| SmWD40-122 | Smechr0700862.1 | 443 | 48787.03 | 8.85 | 30.77 | 78.28 | −0.373 |
| SmWD40-123 | Smechr0700863.1 | 1219 | 136975.43 | 6.55 | 31.74 | 91.23 | −0.244 |
| SmWD40-124 | Smechr0700946.1 | 472 | 51755.65 | 5.08 | 48.5 | 75.4 | −0.434 |
| SmWD40-125 | Smechr0701007.1 | 507 | 55962.59 | 5.45 | 43.86 | 78.42 | −0.403 |
| SmWD40-126 | Smechr0701091.1 | 369 | 40180.43 | 5.37 | 40.88 | 83.79 | −0.132 |
| SmWD40-127 | Smechr0701168.1 | 323 | 34828.15 | 5.6 | 37.42 | 86.04 | −0.074 |
| SmWD40-128 | Smechr0701564.1 | 580 | 65174.62 | 6.28 | 39.93 | 95.5 | −0.206 |
| SmWD40-129 | Smechr0701747.1 | 391 | 42415.3 | 5.34 | 36.39 | 95.73 | −0.109 |
| SmWD40-130 | Smechr0702099.1 | 452 | 51980.04 | 9.59 | 43.59 | 69.87 | −0.662 |
| SmWD40-131 | Smechr0702444.1 | 931 | 102681.23 | 6.26 | 43.32 | 70 | −0.628 |
| SmWD40-132 | Smechr0702707.1 | 313 | 34969.47 | 6.5 | 24.56 | 78.21 | −0.377 |
| SmWD40-133 | Smechr0702744.1 | 816 | 89440.77 | 6.49 | 44.01 | 87.76 | −0.112 |
| SmWD40-134 | Smechr0800148.1 | 734 | 81426.56 | 5.72 | 39.17 | 73.27 | −0.401 |
| SmWD40-135 | Smechr0800182.1 | 422 | 47143.08 | 9.1 | 45.03 | 77.65 | −0.405 |
| SmWD40-136 | Smechr0800245.1 | 417 | 46766.29 | 8.84 | 46.53 | 81.77 | −0.252 |
| SmWD40-137 | Smechr0800403.1 | 656 | 72721.72 | 6.1 | 49.6 | 74.94 | −0.387 |
| SmWD40-138 | Smechr0800720.1 | 440 | 48200.45 | 5.83 | 40.79 | 85.11 | −0.081 |
| SmWD40-139 | Smechr0800881.1 | 1088 | 121328.9 | 5.85 | 43.4 | 89.59 | −0.173 |
| SmWD40-140 | Smechr0801440.1 | 473 | 53597.8 | 8.73 | 50.14 | 75.39 | −0.374 |
| SmWD40-141 | Smechr0801468.1 | 756 | 83339.82 | 8.48 | 55.39 | 54.81 | −0.728 |
| SmWD40-142 | Smechr0801696.1 | 1909 | 207813.61 | 5.27 | 48.43 | 84.89 | −0.365 |
| SmWD40-143 | Smechr0801889.1 | 1137 | 124958.9 | 7 | 44.38 | 77.12 | −0.319 |
| SmWD40-144 | Smechr0802284.1 | 484 | 53175.71 | 9.25 | 51.85 | 75.72 | −0.408 |
| SmWD40-145 | Smechr0802330.1 | 589 | 67263.63 | 6.31 | 45.83 | 62.43 | −0.787 |
| SmWD40-146 | Smechr0802536.1 | 728 | 81007.96 | 6.91 | 38.18 | 71.57 | −0.49 |
| SmWD40-147 | Smechr0900060.1 | 908 | 100784.21 | 8.76 | 50.2 | 72.28 | −0.565 |
| SmWD40-148 | Smechr0900415.1 | 911 | 102220.86 | 8.76 | 49.83 | 71.68 | −0.678 |
| SmWD40-149 | Smechr0900611.1 | 925 | 99984.42 | 8.06 | 38.82 | 88.01 | −0.001 |
| SmWD40-150 | Smechr0900982.1 | 1353 | 148539.39 | 5.95 | 46.24 | 90.78 | −0.088 |
| SmWD40-151 | Smechr0901524.1 | 761 | 83468.49 | 6.57 | 44.54 | 67.95 | −0.554 |
| SmWD40-152 | Smechr0901596.1 | 516 | 58319.57 | 5.52 | 32.1 | 92 | −0.233 |
| SmWD40-153 | Smechr0901745.1 | 318 | 34965.64 | 8.83 | 26.12 | 82.67 | −0.191 |
| SmWD40-154 | Smechr0902013.1 | 349 | 38922.34 | 5.03 | 25.92 | 72.84 | −0.378 |
| SmWD40-155 | Smechr0902163.1 | 599 | 64872.51 | 6.42 | 40.41 | 91.65 | −0.08 |
| SmWD40-156 | Smechr0902418.1 | 521 | 56569.94 | 6.18 | 29.8 | 84.09 | −0.254 |
| SmWD40-157 | Smechr1000020.1 | 300 | 33350.69 | 9.75 | 36.8 | 72.17 | −0.586 |
| SmWD40-158 | Smechr1000478.1 | 377 | 42581.03 | 9.12 | 41.57 | 78.7 | −0.528 |
| SmWD40-159 | Smechr1000759.1 | 212 | 24088.84 | 8.75 | 28.49 | 93.3 | −0.193 |
| SmWD40-160 | Smechr1000926.1 | 341 | 37563.78 | 7.11 | 36.75 | 72.02 | −0.358 |
| SmWD40-161 | Smechr1001004.1 | 315 | 35276.69 | 5.99 | 36.49 | 78 | −0.321 |
| SmWD40-162 | Smechr1001354.1 | 783 | 84904.31 | 6.5 | 47.52 | 76.87 | −0.368 |
| SmWD40-163 | Smechr1001468.1 | 391 | 43489.13 | 8.63 | 50.52 | 81.3 | −0.401 |
| SmWD40-164 | Smechr1001592.1 | 169 | 18088.63 | 9.1 | 42.47 | 92.9 | −0.034 |
| SmWD40-165 | Smechr1001624.1 | 924 | 102745.94 | 6.71 | 53.42 | 64.89 | −0.716 |
| SmWD40-166 | Smechr1001754.1 | 1345 | 148416.1 | 6.06 | 44.42 | 89.62 | −0.097 |
| SmWD40-167 | Smechr1001849.1 | 533 | 59967.06 | 9.58 | 42.66 | 83.32 | −0.38 |
| SmWD40-168 | Smechr1001886.1 | 187 | 20631.09 | 8.57 | 49.91 | 103.1 | 0.107 |
| SmWD40-169 | Smechr1001952.1 | 343 | 38023.91 | 5.38 | 45.56 | 86.91 | −0.145 |
| SmWD40-170 | Smechr1001992.1 | 325 | 35950.37 | 5.77 | 37.24 | 80.37 | −0.229 |
| SmWD40-171 | Smechr1002755.1 | 555 | 61974.65 | 6.07 | 40.04 | 80.22 | −0.611 |
| SmWD40-172 | Smechr1100180.1 | 380 | 41235.45 | 6.23 | 37.84 | 79.34 | −0.309 |
| SmWD40-173 | Smechr1100321.1 | 354 | 38754.14 | 4.72 | 33.59 | 88.95 | −0.342 |
| SmWD40-174 | Smechr1100584.1 | 443 | 48823.45 | 6.5 | 41.29 | 83.43 | −0.227 |
| SmWD40-175 | Smechr1100594.1 | 650 | 72901.54 | 6.33 | 44.64 | 66.37 | −0.547 |
| SmWD40-176 | Smechr1100624.1 | 298 | 33074.38 | 6.85 | 38.76 | 84.97 | −0.233 |
| SmWD40-177 | Smechr1101663.1 | 430 | 47675.99 | 9.16 | 42.56 | 80 | −0.29 |
| SmWD40-178 | Smechr1102321.1 | 821 | 89984.91 | 6.49 | 39.31 | 88.33 | −0.292 |
| SmWD40-179 | Smechr1102445.1 | 580 | 63841.26 | 5.43 | 49.8 | 82.79 | −0.37 |
| SmWD40-180 | Smechr1102608.1 | 493 | 53785.19 | 4.52 | 40.31 | 84.12 | −0.326 |
| SmWD40-181 | Smechr1102707.1 | 424 | 46942.13 | 8.38 | 41.49 | 84.03 | −0.262 |
| SmWD40-182 | Smechr1200022.1 | 584 | 65408.14 | 6.12 | 39.28 | 99.61 | −0.099 |
| SmWD40-183 | Smechr1200183.1 | 323 | 34904.16 | 5.64 | 38.71 | 83.62 | −0.071 |
| SmWD40-184 | Smechr1201542.1 | 476 | 53542.49 | 6.47 | 44.76 | 76.16 | −0.501 |
| SmWD40-185 | Smechr1201594.1 | 557 | 63080.15 | 8.55 | 38.78 | 86.62 | −0.32 |
| SmWD40-186 | Smechr1201735.1 | 1071 | 116791.17 | 6.24 | 38.87 | 73.52 | −0.419 |
| SmWD40-187 | Smechr1201743.1 | 672 | 76211.31 | 6.84 | 41.77 | 83.79 | −0.294 |
References
- Harborne, J.B.; Williams, C.A. Anthocyanins and other flavonoids. Nat. Prod. Rep. 2001, 18, 310–333. [Google Scholar] [CrossRef]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The Effects of Plant Flavonoids on Mammalian Cells: Implications for Inflammation, Heart Disease, and Cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Khan, R.A. Anthocyanins: Traditional Uses, Structural and Functional Variations, Approaches to Increase Yields and Products’ Quality, Hepatoprotection, Liver Longevity, and Commercial Products. Int. J. Mol. Sci. 2022, 23, 2149. [Google Scholar] [CrossRef]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentrations of Anthocyanins in Common Foods in the United States and Estimation of Normal Consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef]
- An, J.; Zhang, X.; Bi, S.; You, C.; Wang, X.; Hao, Y. The ERF transcription factor MdERF38 promotes drought stress-induced anthocyanin biosynthesis in apple. Plant J. 2020, 101, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, S.; Cheng, Y.; Peng, Z.; Han, J. Transcriptome profiling of anthocyanin-related genes reveals effects of light intensity on anthocyanin biosynthesis in red leaf lettuce. PeerJ 2018, 6, e4607. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Lim, S.; Lee, J.G.; Lee, E.J. VcBBX, VcMYB21, and VcR2R3MYB Transcription Factors Are Involved in UV–B-Induced Anthocyanin Biosynthesis in the Peel of Harvested Blueberry Fruit. J. Agric. Food Chem. 2017, 65, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB–bHLH–WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.J.; Zhang, F.J.; Zhang, G.Z.; Jiang, X.Y.; Yu, H.M.; Hou, B.K. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Zhao, M.; Morohashi, K.; Hatlestad, G.; Grotewold, E.; Lloyd, A. The TTG1-bHLH-MYB complex controls trichome cell fate and patterning through direct targeting of regulatory loci. Development 2008, 135, 1991–1999. [Google Scholar] [CrossRef]
- Wei, Q.; Zhang, F.; Sun, F.; Luo, Q.; Wang, R.; Hu, R.; Chen, M.; Chang, J.; Yang, G.; He, G. A wheat MYB transcriptional repressor TaMyb1D regulates phenylpropanoid metabolism and enhances tolerance to drought and oxidative stresses in transgenic tobacco plants. Plant Sci. 2017, 265, 112–123. [Google Scholar] [CrossRef]
- Gil-Muñoz, F.; Sánchez-Navarro, J.A.; Besada, C.; Salvador, A.; Badenes, M.L.; Naval, M.d.M.; Ríos, G. MBW complexes impinge on anthocyanidin reductase gene regulation for proanthocyanidin biosynthesis in persimmon fruit. Sci. Rep. 2020, 10, 3543. [Google Scholar] [CrossRef]
- Xu, W.; Grain, D.; Bobet, S.; Le Gourrierec, J.; Thévenin, J.; Kelemen, Z.; Lepiniec, L.; Dubos, C. Complexity and robustness of the flavonoid transcriptional regulatory network revealed by comprehensive analyses of MYB–b HLH–WDR complexes and their targets in Arabidopsis seed. New Phytol. 2014, 202, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/bHLH/MYB Complexes to Regulate Jasmonate-Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.F.; Gaitatzes, C.; Saxena, K.; Neer, E.J. The WD repeat: A common architecture for diverse functions. Trends Biochem. Sci. 1999, 24, 181–185. [Google Scholar] [CrossRef]
- Stirnimann, C.U.; Petsalaki, E.; Russell, R.B.; Müller, C.W. WD40 proteins propel cellular networks. Trends Biochem. Sci. 2010, 35, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Fong, H.; Hurley, J.B.; Hopkins, R.S.; Miake-Lye, R.; Johnson, M.S.; Doolittle, R.F.; Simon, M.I. Repetitive segmental structure of the transducin beta subunit: Homology with the CDC4 gene and identification of related mRNAs. Proc. Natl. Acad. Sci. USA 1986, 83, 2162–2166. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.A.; Coleman, D.E.; Lee, E.; Iñiguez-Lluhi, J.A.; Posner, B.A.; Gilman, A.G.; Sprang, S.R. The structure of the G protein heterotrimer Giα1β1γ2. Cell 1995, 83, 1047–1058. [Google Scholar] [CrossRef]
- Sondek, J.; Bohm, A.; Lambright, D.G.; Hamm, H.E.; Sigler, P.B. Crystal structure of a GA protein βγdimer at 2.1 Å resolution. Nature 1996, 379, 369–374. [Google Scholar] [CrossRef]
- Miller, J.C.; Chezem, W.R.; Clay, N.K. Ternary WD40 Repeat-Containing Protein Complexes: Evolution, Composition and Roles in Plant Immunity. Front. Plant Sci. 2016, 6, 1108. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 Locus, Which Regulates Trichome Differentiation and Anthocyanin Biosynthesis in Arabidopsis, Encodes a WD40 Repeat Protein. Plant Cell 1999, 11, 1337–1349. [Google Scholar] [CrossRef]
- Schaart, J.G.; Dubos, C.; Romero De La Fuente, I.; van Houwelingen, A.M.; de Vos, R.C.; Jonker, H.H.; Xu, W.; Routaboul, J.M.; Lepiniec, L.; Bovy, A.G. Identification and characterization of MYB-b HLH-WD40 regulatory complexes controlling proanthocyanidin biosynthesis in strawberry (Fragaria × ananassa) fruits. New Phytol. 2013, 197, 454–467. [Google Scholar] [CrossRef]
- Baudry, A.; Caboche, M.; Lepiniec, L. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. 2006, 46, 768–779. [Google Scholar] [CrossRef]
- de Vetten, N.; Quattrocchio, F.; Mol, J.; Koes, R. The an11 locus controlling flower pigmentation in petunia encodes a novel WD-repeat protein conserved in yeast, plants, and animals. Genes Dev. 1997, 11, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.A.; Walker, A.R.; Timmis, J.N.; Orford, S.J. Two WD-repeat genes from cotton are functional homologues of the Arabidopsis thaliana TRANSPARENT TESTA GLABRA1 (TTG1) gene. Plant Mol. Biol. 2005, 57, 67–81. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Tian, Y.; Chen, K.; Wang, X.; Hao, Y. The apple WD40 protein MdTTG1 interacts with bHLH but not MYB proteins to regulate anthocyanin accumulation. J. Plant Physiol. 2012, 169, 710–717. [Google Scholar] [CrossRef]
- Mat Sulaiman, N.N.; Rafii, M.Y.; Duangjit, J.; Ramlee, S.I.; Phumichai, C.; Oladosu, Y.; Datta, D.R.; Musa, I. Genetic Variability of Eggplant Germplasm Evaluated under Open Field and Glasshouse Cropping Conditions. Agronomy 2020, 10, 436. [Google Scholar] [CrossRef]
- Chapman, M.A. Introduction: The Importance of Eggplant; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–10. [Google Scholar] [CrossRef]
- Gisbert, C.; Dumm, J.M.; Prohens, J.; Vilanova, S.; Stommel, J.R. A Spontaneous Eggplant (Solanum melongena L.) Color Mutant Conditions Anthocyanin-free Fruit Pigmentation. HortScience 2016, 51, 793–798. [Google Scholar] [CrossRef]
- Nothmann, J.; Rylski, I.; Spigelman, M. Color and variations in color intensity of fruit of eggplant cultivars. Sci. Hortic. 1976, 4, 191–197. [Google Scholar] [CrossRef]
- Stommel, J.R.; Dumm, J.M. Coordinated Regulation of Biosynthetic and Regulatory Genes Coincides with Anthocyanin Accumulation in Developing Eggplant Fruit. J. Am. Soc. Hortic. Sci. 2015, 140, 129–135. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Ren, L.; Lian, H.; Chen, H. Molecular cloning and characterization of anthocyanin biosynthesis genes in eggplant (Solanum melongena L.). Acta Physiol. Plant. 2016, 38, 163. [Google Scholar] [CrossRef]
- Jiang, M.; Ren, L.; Lian, H.; Liu, Y.; Chen, H. Novel insight into the mechanism underlying light-controlled anthocyanin accumulation in eggplant (Solanum melongena L.). Plant Sci. 2016, 249, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chu, G.; Hu, Z.; Gao, Q.; Cui, B.; Tian, S.; Wang, B.; Chen, G. Genetically engineered anthocyanin pathway for high health-promoting pigment production in eggplant. Mol. Breed. 2016, 36, 54. [Google Scholar] [CrossRef]
- Babak, O.G.; Nekrashevich, N.A.; Nikitinskaya, T.V.; Yatsevich, K.K.; Kilchevsky, A.V. Study of the Myb-factor polymorphism based on comparative genomics of vegetable Solanaceae crops (tomato, pepper, eggplant) to search for DNA markers that differentiate samples by the anthocyans accumulation. Dokl. Natl. Acad. Sci. Belarus 2019, 63, 721–729. [Google Scholar] [CrossRef]
- Zhou, L.; He, Y.; Li, J.; Liu, Y.; Chen, H. CBFs Function in Anthocyanin Biosynthesis by Interacting with MYB113 in Eggplant (Solanum melongena L.). Plant Cell Physiol. 2020, 61, 416–426. [Google Scholar] [CrossRef]
- Li, L.; He, Y.; Ge, H.; Liu, Y.; Chen, H. Functional characterization of SmMYB86, a negative regulator of anthocyanin biosynthesis in eggplant (Solanum melongena L.). Plant Sci. 2021, 302, 110696. [Google Scholar] [CrossRef]
- Montefiori, M.; Brendolise, C.; Dare, A.P.; Lin-Wang, K.; Davies, K.M.; Hellens, R.P.; Allan, A.C. In the Solanaceae, a hierarchy of bHLHs confer distinct target specificity to the anthocyanin regulatory complex. J. Exp. Bot. 2015, 66, 1427–1436. [Google Scholar] [CrossRef]
- Li, W.; Wang, B.; Wang, M.; Chen, M.; Yin, J.M.; Kaleri, G.M.; Zhang, R.J.; Zuo, T.N.; You, X.; Yang, Q. Cloning and characterization of a potato StAN11 gene involved in anthocyanin biosynthesis regulation. J. Integr. Plant Biol. 2014, 56, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, J.; Chen, Y.; Tang, H.; Wang, Y.; He, Y.; Ou, Y.; Sun, X.; Wang, S.; Yao, Y. Tomato SlAN11 regulates flavonoid biosynthesis and seed dormancy by interaction with bHLH proteins but not with MYB proteins. Hortic. Res. 2018, 5, 27. [Google Scholar] [CrossRef]
- Collonnier, C.; Fock, I.; Kashyap, V.; Rotino, G.; Daunay, M.; Lian, Y.; Mariska, I.; Rajam, M.; Servaes, A.; Ducreux, G. Applications of biotechnology in eggplant. Plant Cell Tissue Organ Cult. 2001, 65, 91–107. [Google Scholar] [CrossRef]
- Gürbüz, N.; Uluişik, S.; Frary, A.; Frary, A.; Doğanlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, A.; Wu, X.; Zhu, Z.; Yang, Z.; Zhu, Y.; Zha, D. Transcriptome analysis revealed expression of genes related to anthocyanin biosynthesis in eggplant (Solanum melongena L.) under high-temperature stress. BMC Plant Biol. 2019, 19, 387. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Chu, G.; Huang, C.; Tian, S.; Zhao, Z.; Chen, G. Anthocyanin Accumulation and Molecular Analysis of Anthocyanin Biosynthesis-Associated Genes in Eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 2906–2912. [Google Scholar] [CrossRef]
- Ahmed, N.U.; Park, J.-I.; Jung, H.-J.; Hur, Y.; Nou, I.-S. Anthocyanin biosynthesis for cold and freezing stress tolerance and desirable color in Brassica rap. Funct. Integr. Genom. 2015, 15, 383–394. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, D.; Zhu, B.; Yan, H.; Shen, X.; Zuo, J.; Zhu, Y.; Luo, Y. Virus-induced Gene Silencing in Eggplant (Solanum melongena). J. Integr. Plant Biol. 2012, 54, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, J.; Wang, W.; Hu, T.; Hu, H.; Bao, C. A high-quality chromosome-level genome assembly reveals genetics for important traits in eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef]
- Casta eda-Ovando, A.; Pacheco-Hernández, M.d.L.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Holton, T.A.; Cornish, E.C. Genetics and Biochemistry of Anthocyanin Biosynthesis. Plant Cell 1995, 7, 1071. [Google Scholar] [CrossRef]
- Passeri, V.; Koes, R.; Quattrocchio, F.M. New Challenges for the Design of High Value Plant Products: Stabilization of Anthocyanins in Plant Vacuoles. Front. Plant Sci. 2016, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C.N.; Koski, M.H. The effects of climate change on floral anthocyanin polymorphisms. Proc. R. Soc. B 2021, 288, 20202693. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Sun, L.; Wei, L.; Yuan, J.; Kong, F.; Zhang, Y.; Miao, X.; Xia, G.; Liu, S. Maize SRO1e represses anthocyanin synthesis through regulating the MBW complex in response to abiotic stress. Plant J. 2021, 105, 1010–1025. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.; Visser, R.G.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Giusti, M.M. Anthocyanins: Natural Colorants with Health-Promoting Properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Cho, M.; Choi, M.G.; Das, P.K.; Lee, S.-K.; Choi, S.-B.; Park, Y.-I. Identification of genes that may regulate the expression of the transcription factor production of anthocyanin pigment 1 (PAP1)/MYB75 involved in Arabidopsis anthocyanin biosynthesis. Plant Cell Rep. 2015, 34, 805–815. [Google Scholar] [CrossRef]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, N.A.; Glover, B.J. MYB–bHLH–WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005, 10, 63–70. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ohmiya, A.J.C.o.i.b. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotechnol. 2008, 19, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Yang, T.; Wang, B.; Yang, H.; Wang, J.; Yu, Q. Genome-Wide Identification of the WD40 Gene Family in Tomato (Solanum lycopersicum L.). Genes 2023, 14, 1273. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ye, D.; Li, Y.; Hu, P.; Xu, R.; Wang, X. Genome-wide identification and bioinformatics analysis of the WD40 transcription factor family and candidate gene screening for anthocyanin biosynthesis in Rhododendron simsii. Front. Genet. 2023, 14, 1172321. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Y.; Coulter, J.A.; Shen, B.; Li, Y.; Li, C.; Cao, Z.; Zhang, J. The WD40 gene family in potato (Solanum tuberosum L.): Genome-wide analysis and identification of anthocyanin and drought-related WD40s. Agronomy 2020, 10, 401. [Google Scholar] [CrossRef]
- Gutierrez, N.; Torres, A.M. Characterization and diagnostic marker for TTG1 regulating tannin and anthocyanin biosynthesis in faba bean. Sci. Rep. 2019, 9, 16174. [Google Scholar] [CrossRef]
- Ben-Simhon, Z.; Judeinstein, S.; Nadler-Hassar, T.; Trainin, T.; Bar-Ya’akov, I.; Borochov-Neori, H.; Holland, D. A pomegranate (Punica granatum L.) WD40-repeat gene is a functional homologue of Arabidopsis TTG1 and is involved in the regulation of anthocyanin biosynthesis during pomegranate fruit development. Planta 2011, 234, 865–881. [Google Scholar] [CrossRef]
- Song, S.; Liu, B.; Song, J.; Pang, S.; Song, T.; Gao, S.; Zhang, Y.; Huang, H.; Qi, T. A molecular framework for signaling crosstalk between jasmonate and ethylene in anthocyanin biosynthesis, trichome development, and defenses against insect herbivores in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 1770–1788. [Google Scholar] [CrossRef]
- Clifford, M.N. Anthocyanins–nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1063–1072. [Google Scholar] [CrossRef]
- Timbola, A.K.; Souza, C.D.d.; Giacomelli, C.; Spinelli, A. Electrochemical oxidation of quercetin in hydro-alcoholic solution. J. Braz. Chem. Soc. 2006, 17, 139–148. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 261. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Pei, X.; Zhang, H.; Li, X.; Zhang, X.; Zhao, M.; Chiang, V.L.; Sederoff, R.R.; Zhao, X. MYB-mediated regulation of anthocyanin biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Cai, Q.; Wang, Y.; Li, L.; Sun, Z. Genome-Wide Exploration of the WD40 Gene Family in Eggplant (Solanum melongena L.) and Analysis of Its Function in Fruit Color Formation. Agronomy 2024, 14, 521. https://doi.org/10.3390/agronomy14030521
Yang Y, Cai Q, Wang Y, Li L, Sun Z. Genome-Wide Exploration of the WD40 Gene Family in Eggplant (Solanum melongena L.) and Analysis of Its Function in Fruit Color Formation. Agronomy. 2024; 14(3):521. https://doi.org/10.3390/agronomy14030521
Chicago/Turabian StyleYang, Yanbo, Qihang Cai, Yimei Wang, Liping Li, and Zhenghai Sun. 2024. "Genome-Wide Exploration of the WD40 Gene Family in Eggplant (Solanum melongena L.) and Analysis of Its Function in Fruit Color Formation" Agronomy 14, no. 3: 521. https://doi.org/10.3390/agronomy14030521
APA StyleYang, Y., Cai, Q., Wang, Y., Li, L., & Sun, Z. (2024). Genome-Wide Exploration of the WD40 Gene Family in Eggplant (Solanum melongena L.) and Analysis of Its Function in Fruit Color Formation. Agronomy, 14(3), 521. https://doi.org/10.3390/agronomy14030521