
Evaluation of Soil Total Nitrogen as an Indicator of Soil Bacterial Community Response to Biochar and Plant Growth-Promoting Rhizobacteria Applications


Abstract
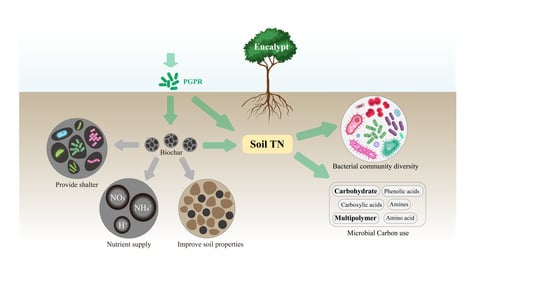
1. Introduction
2. Materials and Methods
2.1. Research Site
2.2. PGPR and Biochar Characterizations
2.3. Experimental Design
2.4. Field Sampling and Lab Measurements
2.5. Illumina High-Throughput Sequencing
2.6. Statistical Analysis of Data
3. Results
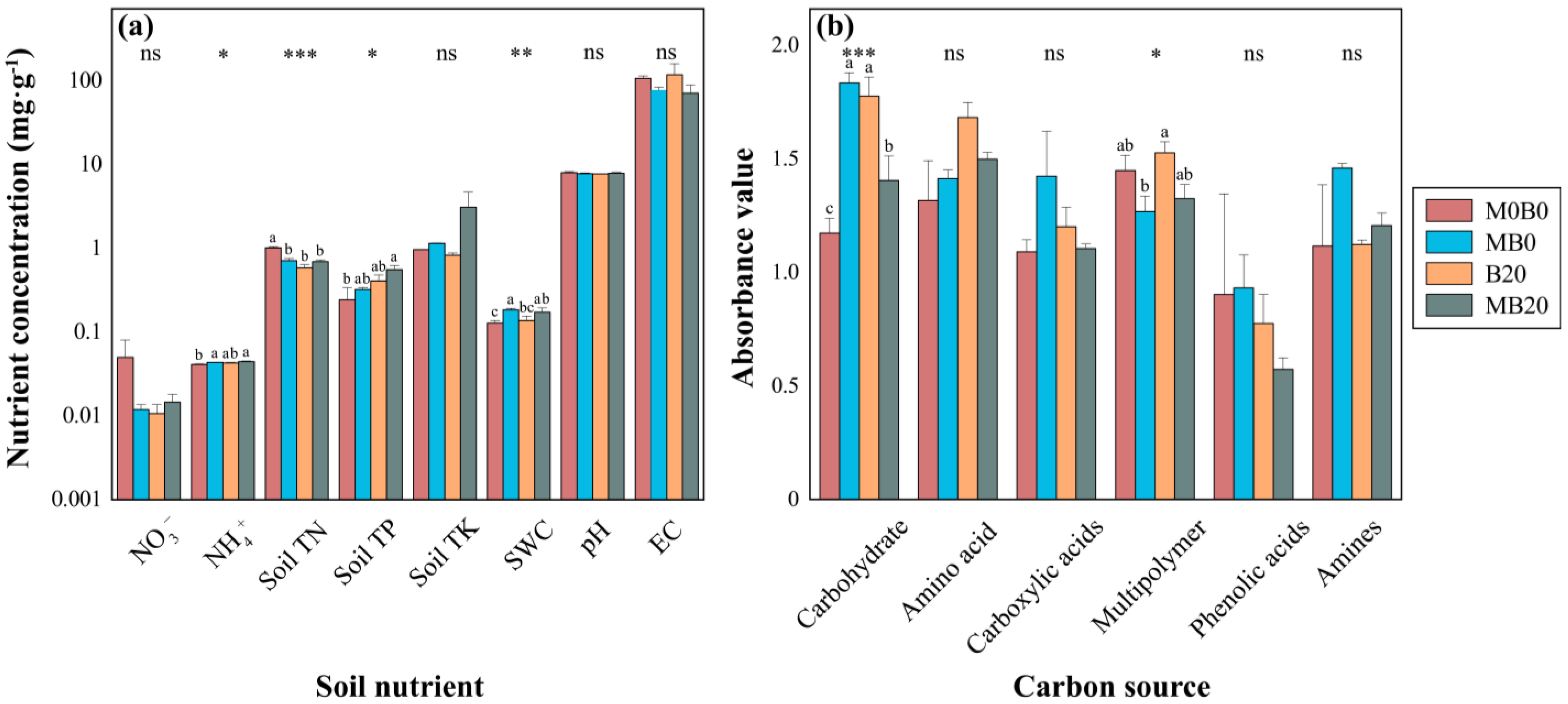
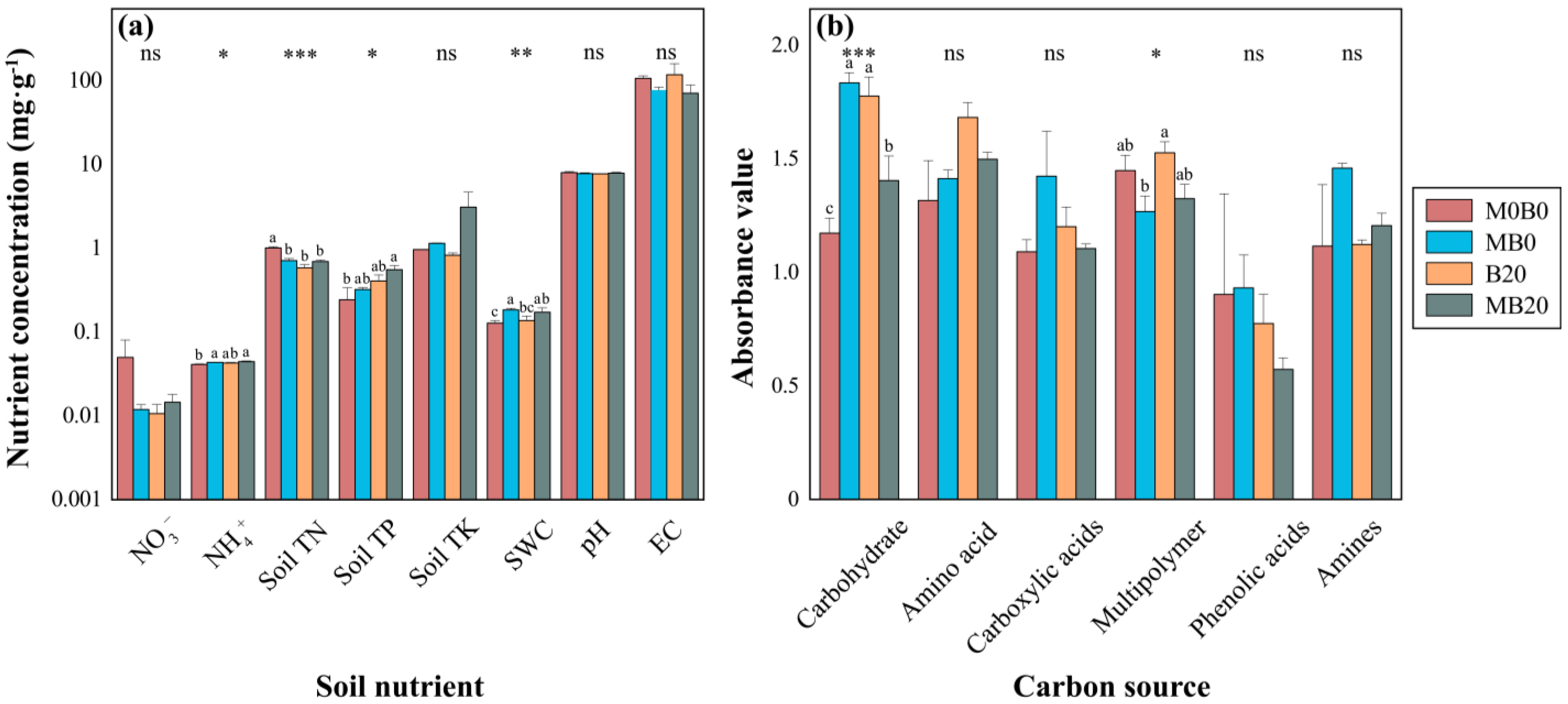
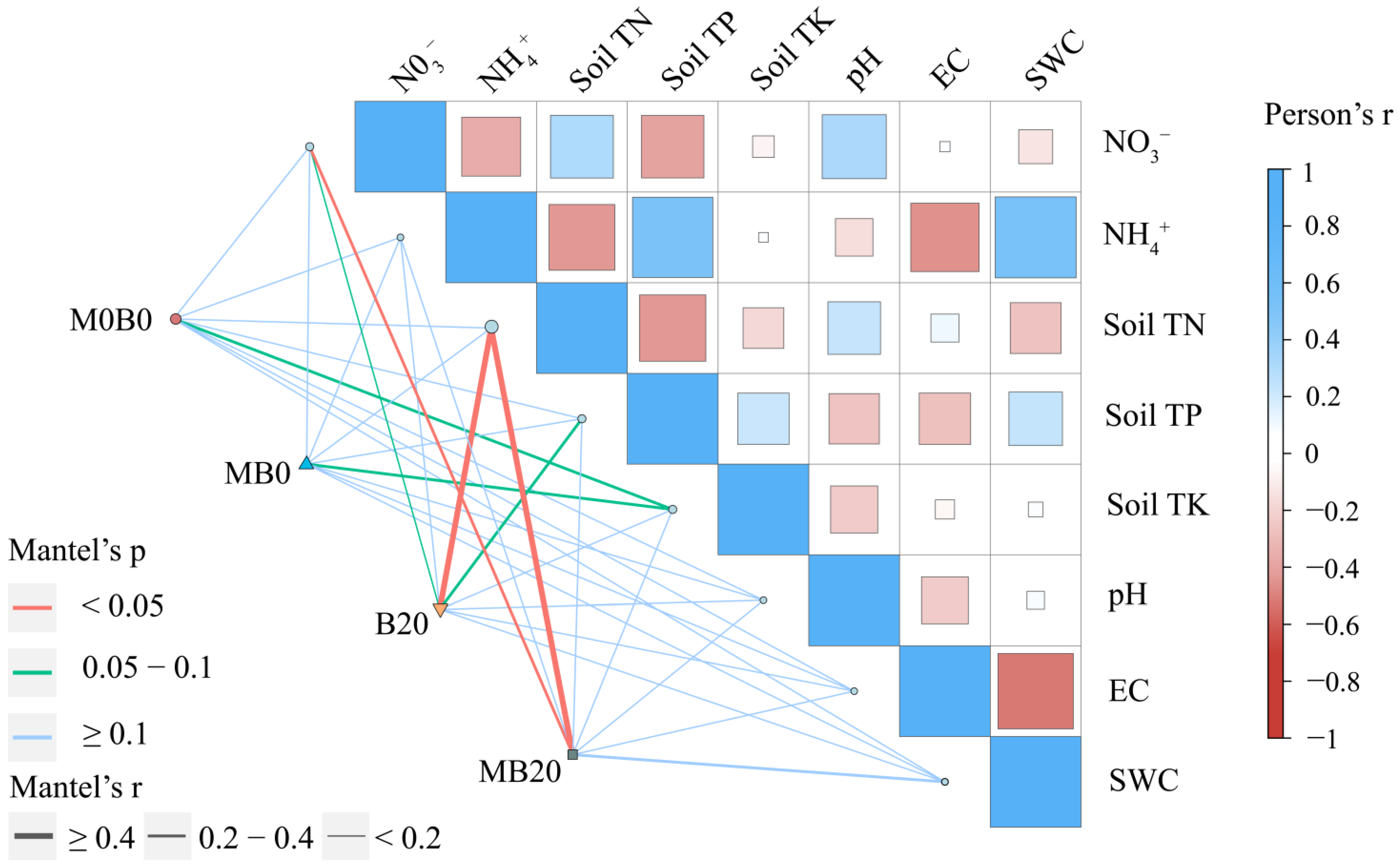
3.1. Soil Physicochemical Properties
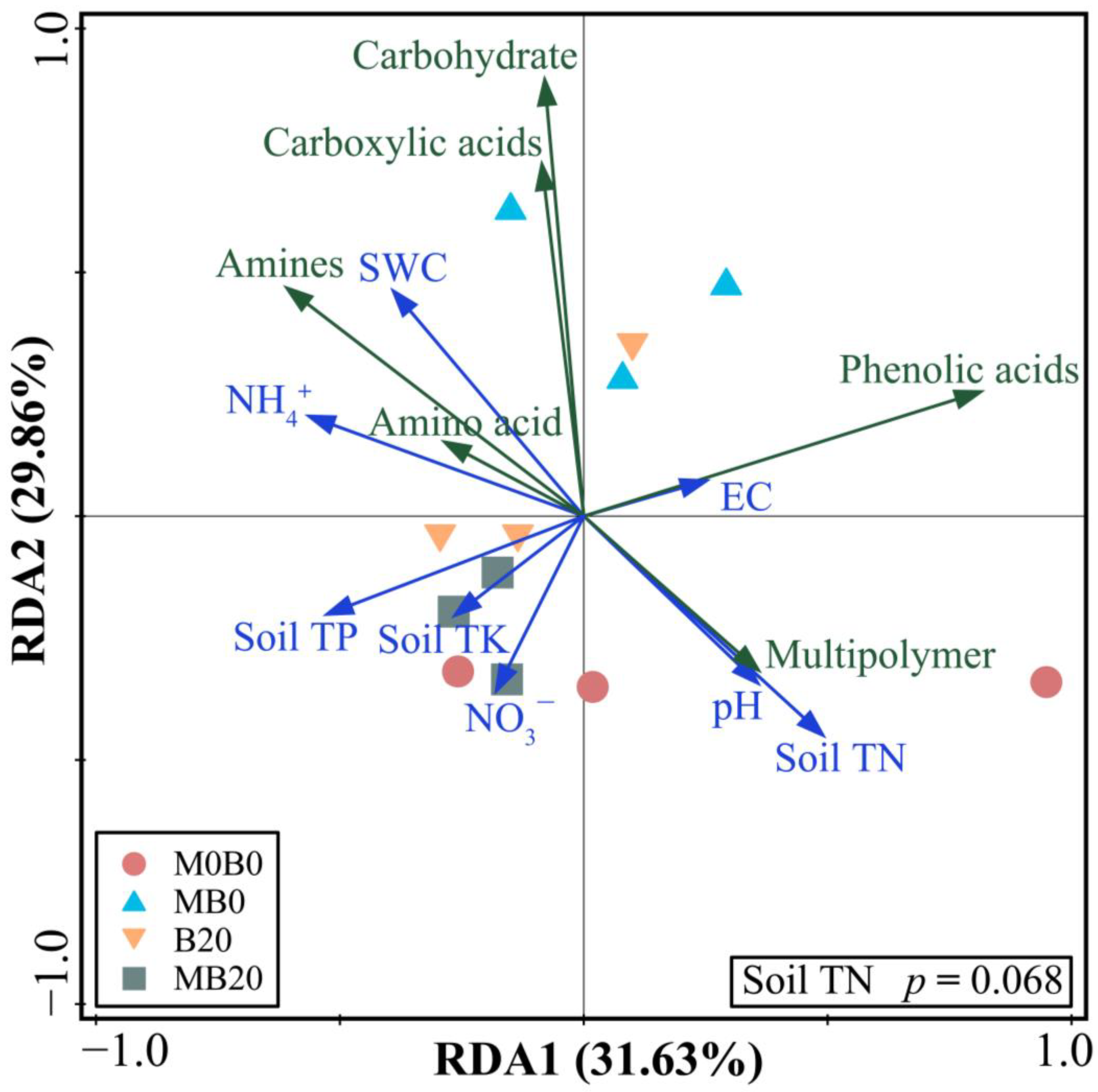
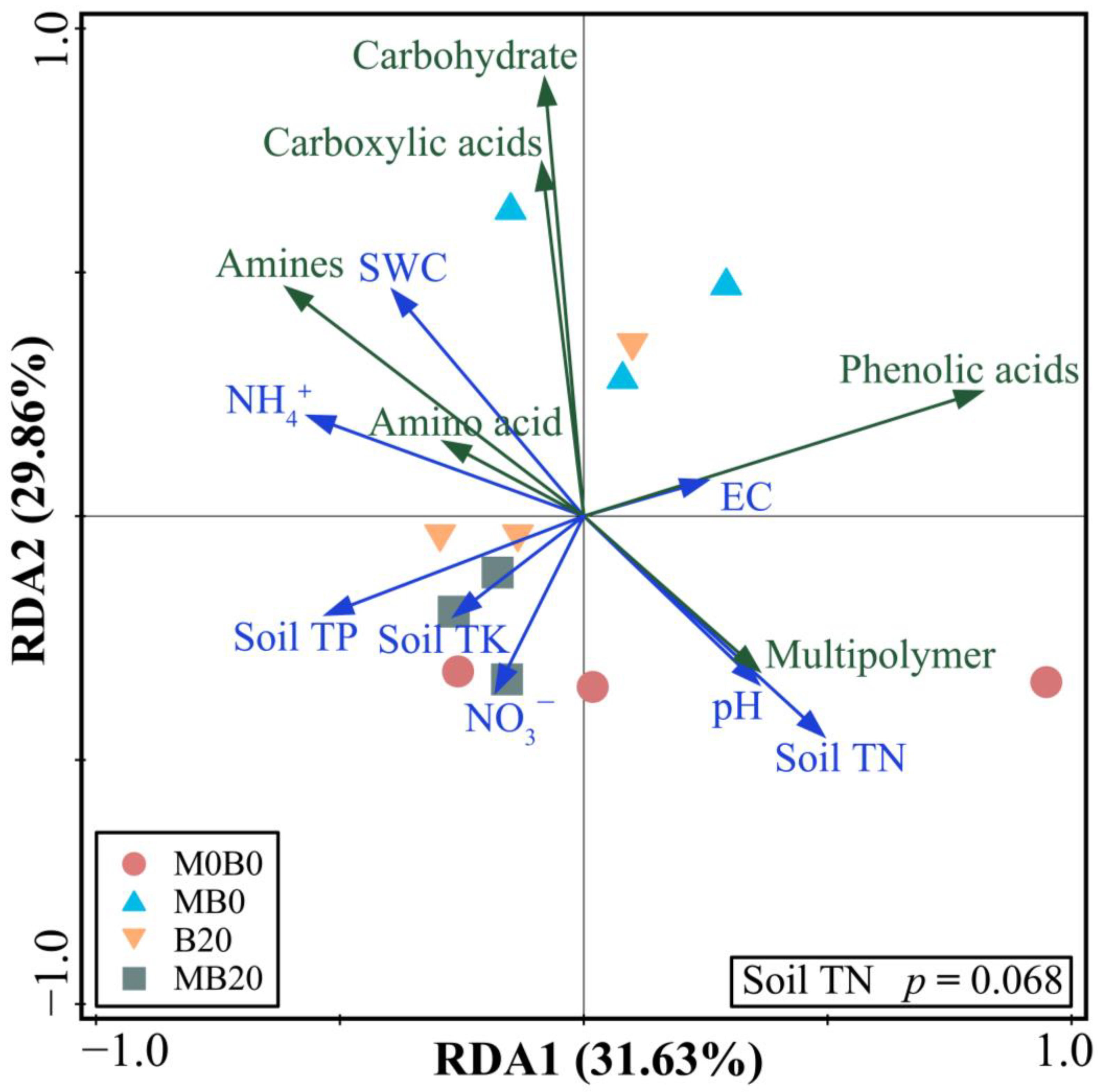
3.2. Soil Microbial Functional Diversity
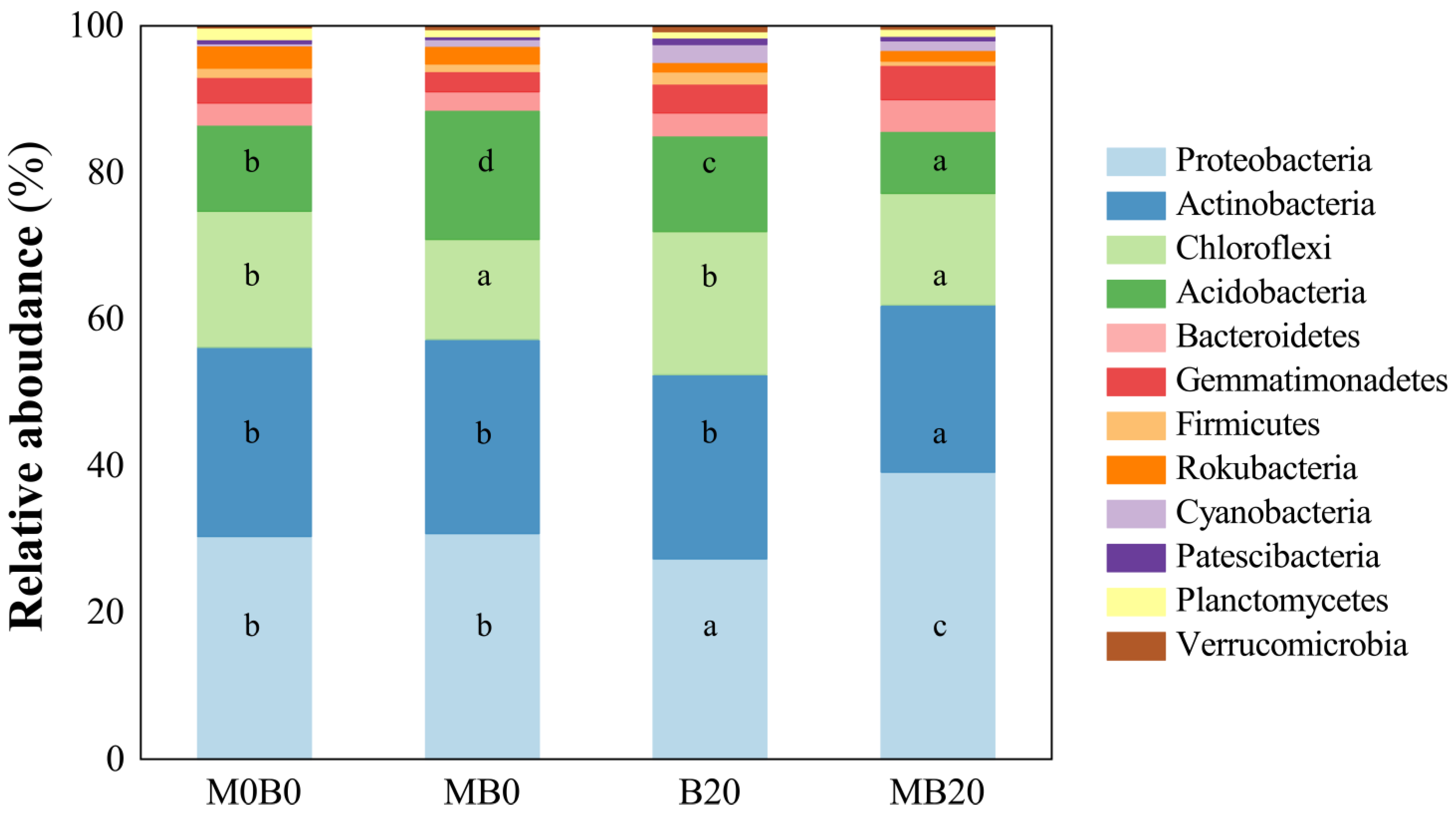
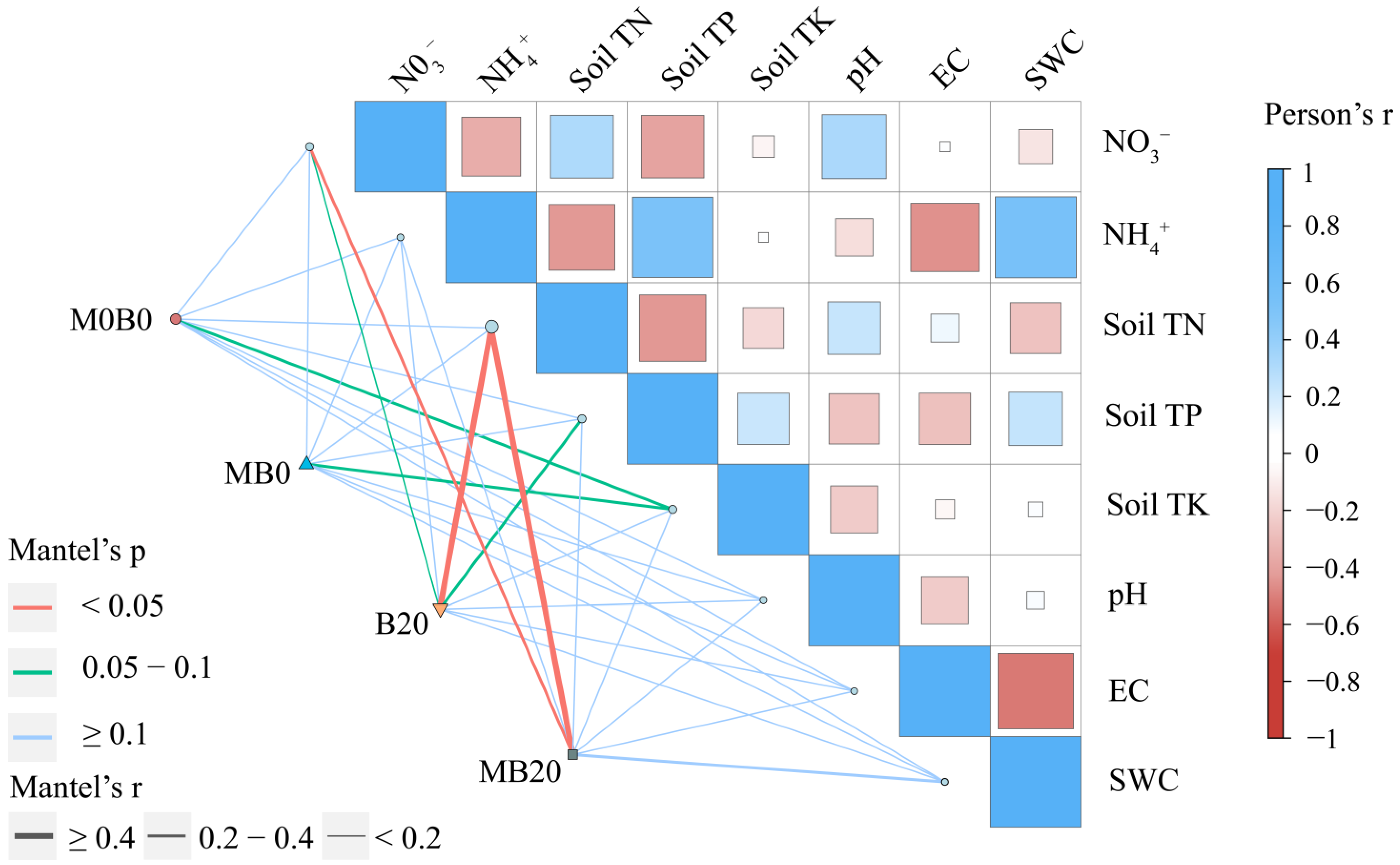
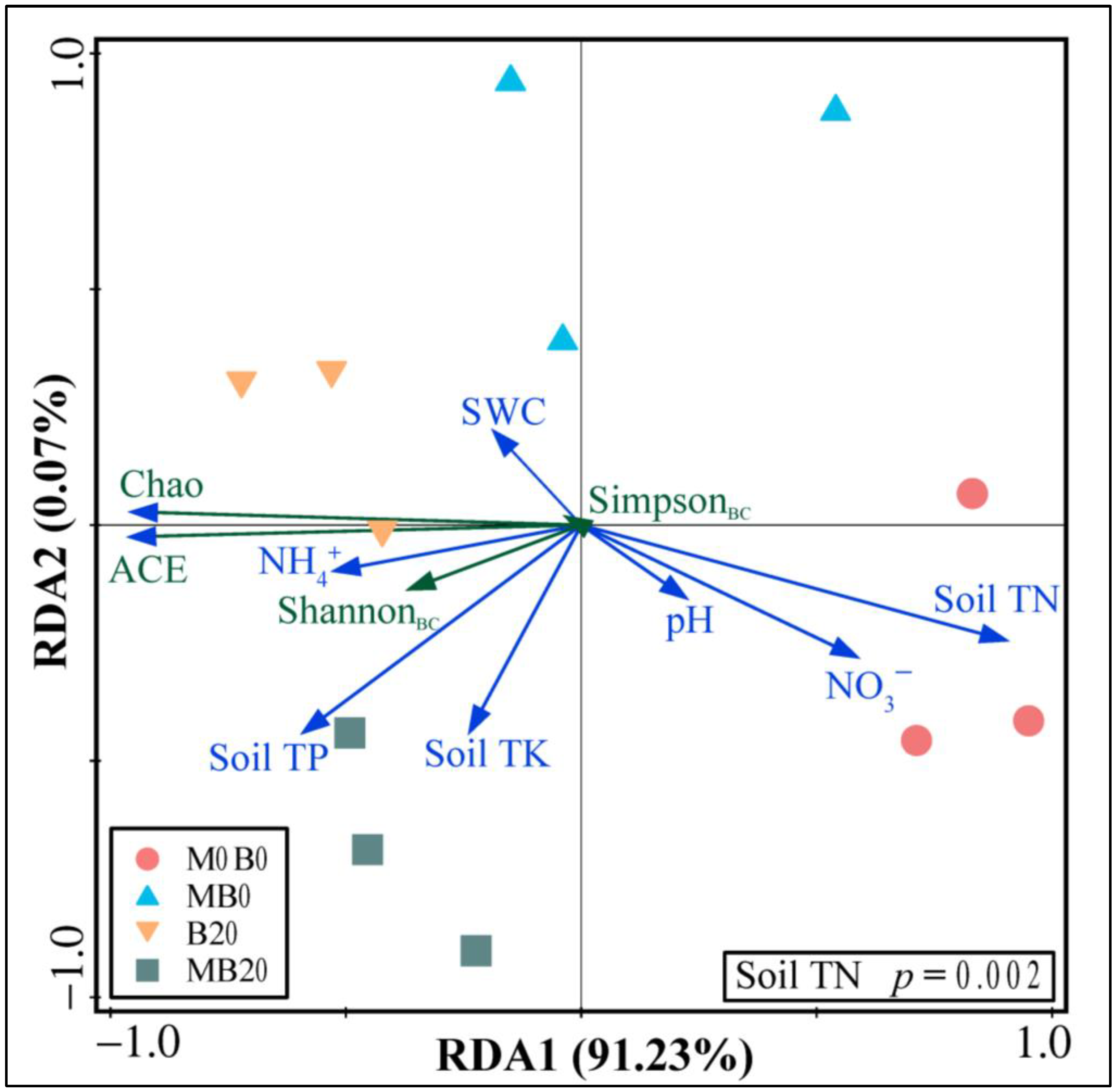
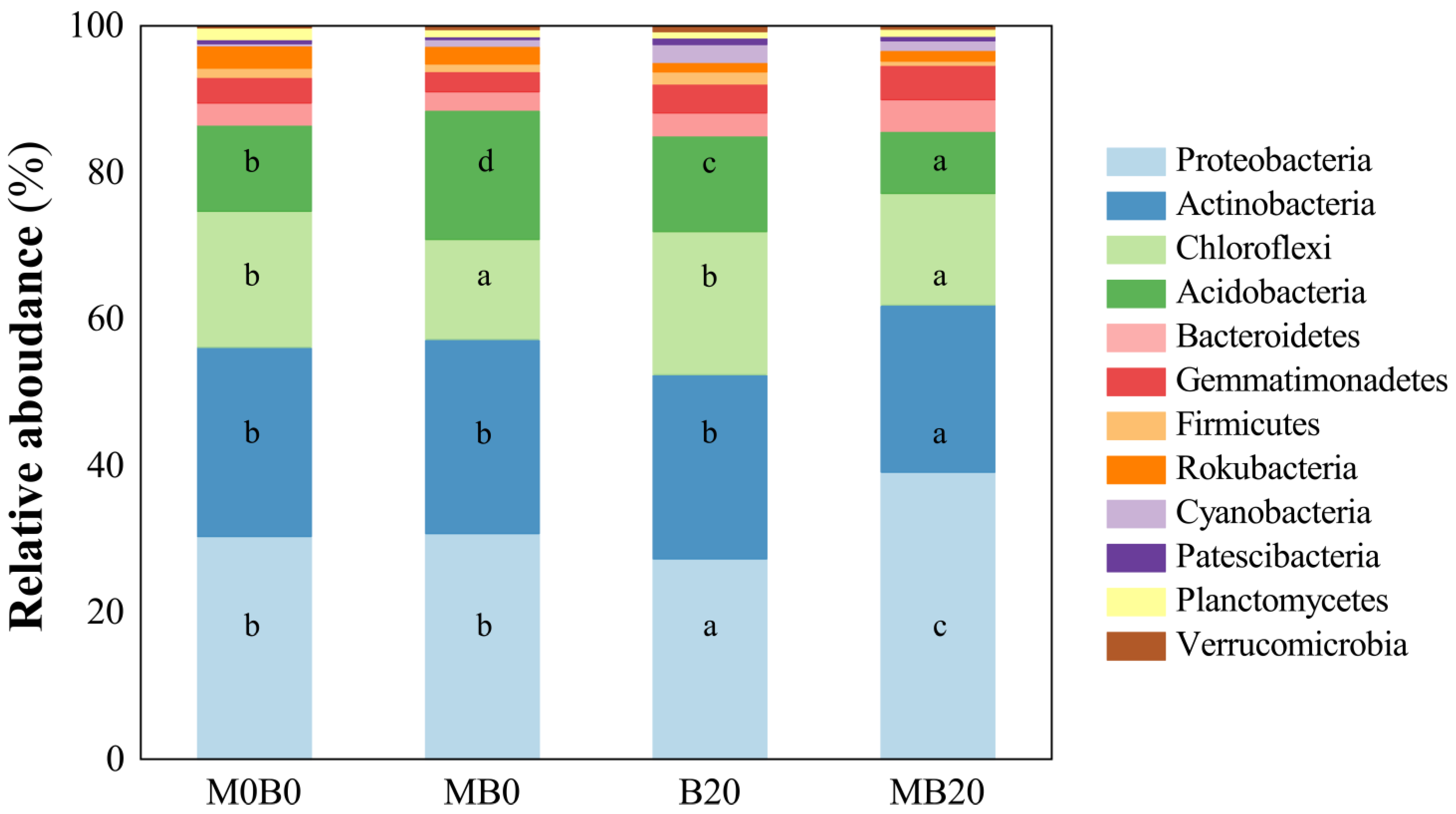
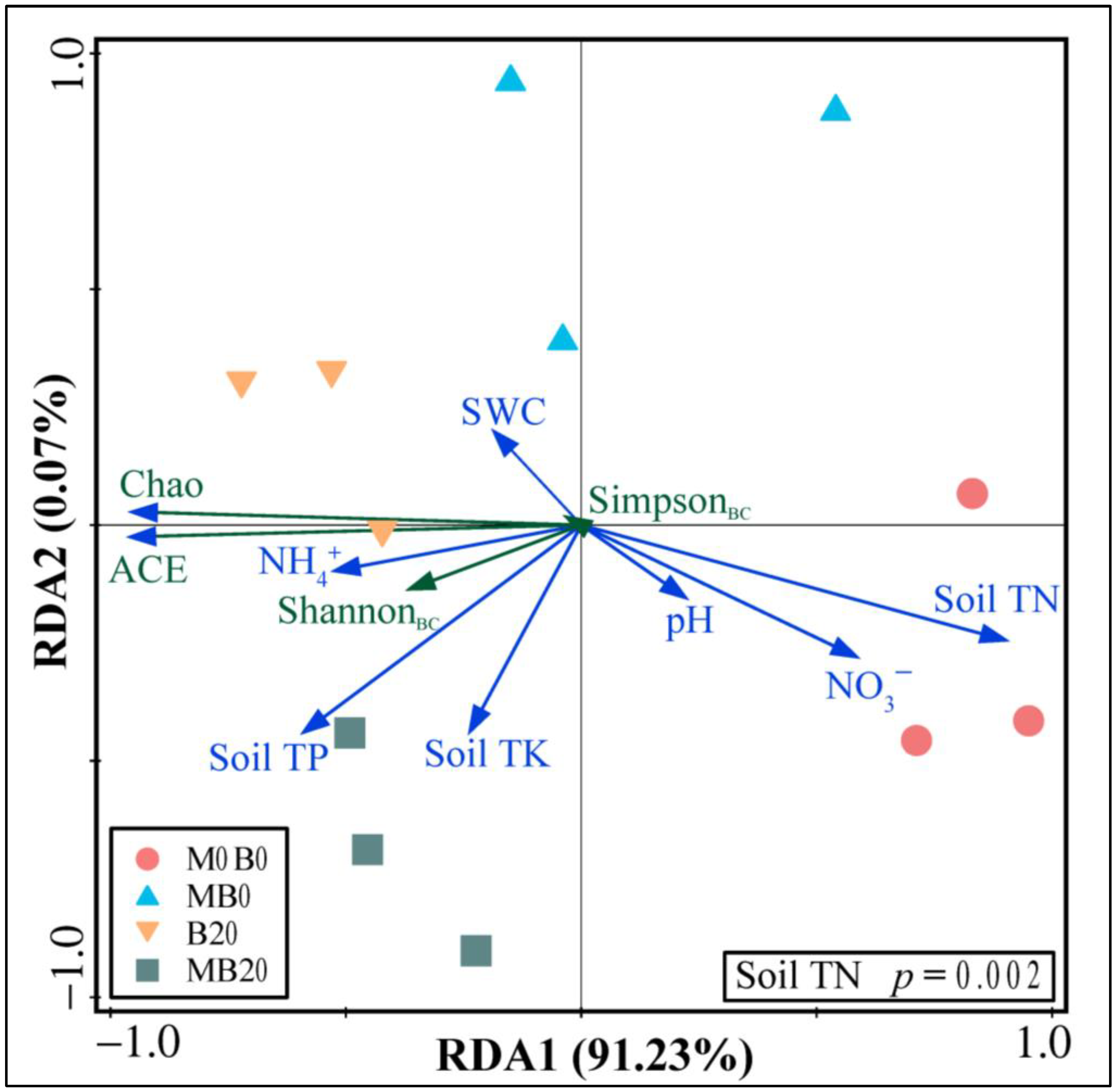
3.3. Soil Bacterial Community Diversity
4. Discussion
4.1. Soil Nutrient Concentration
4.2. Soil Microbial Functional Diversification
4.3. Soil Microbial Evaluation
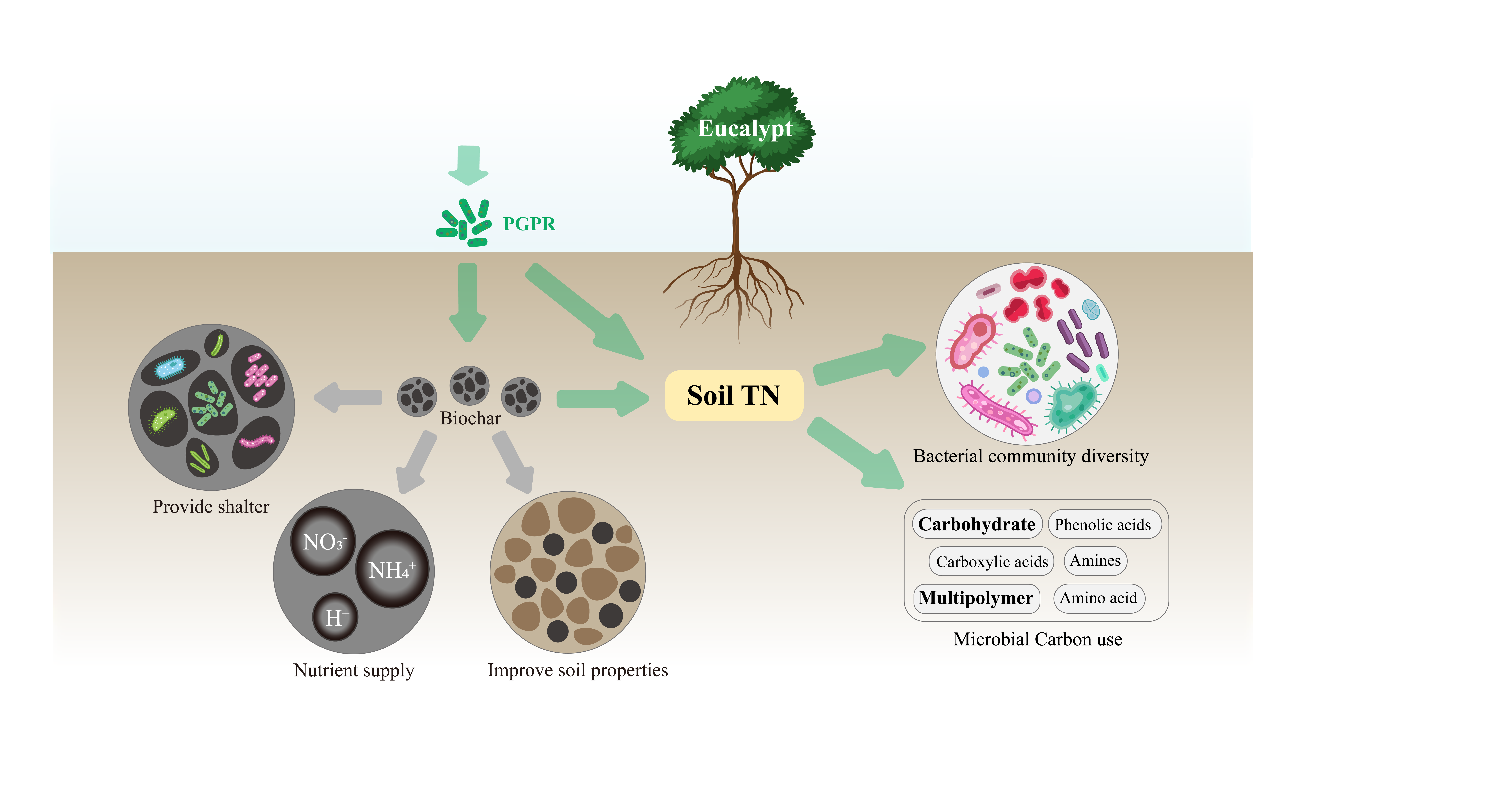
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Malik, L.; Sanaullah, M.; Mahmood, F.; Hussain, S.; Siddique, M.H.; Anwar, F.; Shahzad, T. Unlocking the Potential of Co-Applied Biochar and Plant Growth-Promoting Rhizobacteria (PGPR) for Sustainable Agriculture under Stress Conditions. Chem. Biol. Technol. Agric. 2022, 9, 58. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Dardanelli, M.S.; Manyani, H.; González-Barroso, S.; Rodríguez-Carvajal, M.A.; Gil-Serrano, A.M.; Espuny, M.R.; López-Baena, F.J.; Bellogín, R.A.; Megías, M.; Ollero, F.J. Effect of the Presence of the Plant Growth Promoting Rhizobacterium (PGPR) Chryseobacterium Balustinum Aur9 and Salt Stress in the Pattern of Flavonoids Exuded by Soybean Roots. Plant Soil 2010, 328, 483–493. [Google Scholar] [CrossRef]
- Tkacz, A.; Poole, P. Role of Root Microbiota in Plant Productivity. J. Exp. Bot. 2015, 66, 2167–2175. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Huang, B.; Fernandez-Garcia, V.; Miesel, J.; Yan, L.; Lv, C. Biochar and Rhizobacteria Amendments Improve Several Soil Properties and Bacterial Diversity. Microorganisms 2020, 8, 502. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yu, Y.; Chang, E.; Wang, R.; Hong, Z.; Cui, J.; Zhang, F.; Jiang, J.; Xu, R. Effect of Biochar Incorporation on Phosphorus Supplementation and Availability in Soil: A Review. J. Soils Sediments 2023, 23, 672–686. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Chang, S.X.; Yang, Y.; Fu, S.; Jiang, P.; Luo, Y.; Yang, M.; Chen, Z.; Hu, S.; et al. Biochar Reduces Soil Heterotrophic Respiration in a Subtropical Plantation through Increasing Soil Organic Carbon Recalcitrancy and Decreasing Carbon Degrading Microbial Activity. Soil Biol. Biochem. 2018, 122, 173–185. [Google Scholar] [CrossRef]
- Awad, Y.M.; Ok, Y.S.; Abrigata, J.; Beiyuan, J.; Beckers, F.; Tsang, D.C.W.; Rinklebe, J. Pine Sawdust Biomass and Biochars at Different Pyrolysis Temperatures Change Soil Redox Processes. Sci. Total Environ. 2018, 625, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Kolton, M.; Graber, E.R.; Tsehansky, L.; Elad, Y.; Cytryn, E. Biochar-Stimulated Plant Performance Is Strongly Linked to Microbial Diversity and Metabolic Potential in the Rhizosphere. New Phytol. 2017, 213, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Warnock, D.D.; Tiemann, L.K.; Quigley, K.; Miesel, J.R. Evaluating Foliar Characteristics as Early Indicators of Plant Response to Biochar Amendments. For. Ecol. Manag. 2021, 489, 119047. [Google Scholar] [CrossRef]
- Nasiri, S.; Andalibi, B.; Tavakoli, A.; Delavar, M.A.; El-Keblawy, A.; Zwieten, L.V.; Mastinu, A. The Mineral Biochar Alters the Biochemical and Microbial Properties of the Soil and the Grain Yield of Hordeum vulgare L. under Drought Stress. Land 2023, 12, 559. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, B.; Zhu, L.; Xing, B. Effects and Mechanisms of Biochar-Microbe Interactions in Soil Improvement and Pollution Remediation: A Review. Environ. Pollut. 2017, 227, 98–115. [Google Scholar] [CrossRef]
- Farrell, M.; Kuhn, T.K.; Macdonald, L.M.; Maddern, T.M.; Murphy, D.V.; Hall, P.A.; Singh, B.P.; Baumann, K.; Krull, E.S.; Baldock, J.A. Microbial Utilisation of Biochar-Derived Carbon. Sci. Total Environ. 2013, 465, 288–297. [Google Scholar] [CrossRef]
- Choppala, G.; Bolan, N.; Kunhikrishnan, A.; Bush, R. Differential Effect of Biochar upon Reduction-Induced Mobility and Bioavailability of Arsenate and Chromate. Chemosphere 2016, 144, 374–381. [Google Scholar] [CrossRef]
- Wong, J.T.F.; Chen, X.; Deng, W.; Chai, Y.; Ng, C.W.W.; Wong, M.H. Effects of Biochar on Bacterial Communities in a Newly Established Landfill Cover Topsoil. J. Environ. Manag. 2019, 236, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Gong, T.; Wang, J.; Li, G.; Liu, Y.; Zhen, J.; Ning, M.; Yue, D.; Du, Z.; Chen, G. Effects of Compound Microbial Fertilizer on Soil Characteristics and Yield of Wheat (Triticum aestivum L.). J. Soil Sci. Plant Nutr. 2020, 20, 2740–2748. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Shi, P.; Bi, Z.; Shan, Z.; Ren, L. The Deep Challenge of Nitrate Pollution in River Water of China. Sci. Total Environ. 2021, 770, 144674. [Google Scholar] [CrossRef] [PubMed]
- Lazaratou, C.V.; Vayenas, D.V.; Papoulis, D. The Role of Clays, Clay Minerals and Clay-Based Materials for Nitrate Removal from Water Systems: A Review. Appl. Clay Sci. 2020, 185, 105377. [Google Scholar] [CrossRef]
- Wang, R.-C.; Wang, H.-M.; Xiang, X.; Gao, Y.; Song, Q.-W.; Gong, L.-F. Temporal and Spatial Variations of Microbial Carbon Utilization in Water Bodies from the Dajiuhu Peatland, Central China. J. Earth Sci. 2018, 29, 969–976. [Google Scholar] [CrossRef]
- Kim, Y.C.; Anderson, A.J. Rhizosphere Pseudomonads as Probiotics Improving Plant Health. Mol. Plant Pathol. 2018, 19, 2349–2359. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Xu, C.-Y.; Tahmasbian, I.; Che, R.; Xu, Z.; Zhou, X.; Wallace, H.M.; Bai, S.H. Effects of Biochar on Soil Available Inorganic Nitrogen: A Review and Meta-Analysis. Geoderma 2017, 288, 79–96. [Google Scholar] [CrossRef]
- D’Elia, C.F.; Steudler, P.A.; Corwin, N. Determination of Total Nitrogen in Aqueous Samples Using Persulfate Digestion1. Limnol. Oceanogr. 1977, 22, 760–764. [Google Scholar] [CrossRef]
- Ren, H.; Li, Z.; Chen, H.; Zhou, J.; Lv, C. Effects of Biochar and Plant Growth-Promoting Rhizobacteria on Plant Performance and Soil Environmental Stability. Sustainability 2022, 14, 10922. [Google Scholar] [CrossRef]
- Ker, K.; Seguin, P.; Driscoll, B.T.; Fyles, J.W.; Smith, D.L. Evidence for Enhanced N Availability during Switchgrass Establishment and Seeding Year Production Following Inoculation with Rhizosphere Endophytes. Arch. Agron. Soil Sci. 2014, 60, 1553–1563. [Google Scholar] [CrossRef]
- Sahin, O.; Taskin, M.B.; Kaya, E.C.; Atakol, O.; Emir, E.; Inal, A.; Gunes, A. Effect of Acid Modification of Biochar on Nutrient Availability and Maize Growth in a Calcareous Soil. Soil Use Manag. 2017, 33, 447–456. [Google Scholar] [CrossRef]
- Chen, L.; Wang, L.; Lu, X.; Yin, Q.; Liu, D. Utilization of Plant Litter to Enhance the Effect of Lakebank on Nitrogen Interception/Removal from Runoff: A Microcosm Simulation Study. Environ. Eng. Sci. 2022, 39, 235–247. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Z.; Liu, S. HMGB1 Induced Inflammatory Effect Is Blocked by CRISPLD2 via MiR155 in Hepatic Fibrogenesis. Mol. Immunol. 2016, 69, 1–6. [Google Scholar] [CrossRef]
- Sun, B.; Gu, L.; Bao, L.; Zhang, S.; Wei, Y.; Bai, Z.; Zhuang, G.; Zhuang, X. Application of Biofertilizer Containing Bacillus subtilis Reduced the Nitrogen Loss in Agricultural Soil. Soil Biol. Biochem. 2020, 148, 107911. [Google Scholar] [CrossRef]
- Song, Z.; Su, X.; Li, P.; Sun, F.; Dong, W.; Zhao, Z.; Wen, Z.; Liao, R. Facial Fabricated Biocompatible Homogeneous Biocarriers Involving Biochar to Enhance Denitrification Performance in an Anoxic Moving Bed Biofilm Reactor. Bioresour. Technol. 2021, 341, 125866. [Google Scholar] [CrossRef]
- Kuan, K.B.; Othman, R.; Abdul Rahim, K.; Shamsuddin, Z.H. Plant Growth-Promoting Rhizobacteria Inoculation to Enhance Vegetative Growth, Nitrogen Fixation and Nitrogen Remobilisation of Maize under Greenhouse Conditions. PLoS ONE 2016, 11, e0152478. [Google Scholar] [CrossRef]
- Yousuf, J.; Thajudeen, J.; Rahiman, M.; Krishnankutty, S.; Alikunj, A.P.; Abdulla, M.H.A. Nitrogen Fixing Potential of Various Heterotrophic Bacillus Strains from a Tropical Estuary and Adjacent Coastal Regions. J. Basic Microbiol. 2017, 57, 922–932. [Google Scholar] [CrossRef]
- Nelissen, V.; Rutting, T.; Huygens, D.; Staelens, J.; Ruysschaert, G.; Boeckx, P. Maize Biochars Accelerate Short-Term Soil Nitrogen Dynamics in a Loamy Sand Soil. Soil Biol. Biochem. 2012, 55, 20–27. [Google Scholar] [CrossRef]
- Uzinger, N.; Takács, T.; Szili-Kovács, T.; Radimszky, L.; Füzy, A.; Draskovits, E.; Szűcs-Vásárhelyi, N.; Molnár, M.; Farkas, É.; Kutasi, J.; et al. Fertility Impact of Separate and Combined Treatments with Biochar, Sewage Sludge Compost and Bacterial Inocula on Acidic Sandy Soil. Agronomy 2020, 10, 1612. [Google Scholar] [CrossRef]
- Soltangheisi, A.; Rodrigues, M.; Coelho, M.J.A.; Gasperini, A.M.; Sartor, L.R.; Pavinato, P.S. Changes in Soil Phosphorus Lability Promoted by Phosphate Sources and Cover Crops. Soil Tillage Res. 2018, 179, 20–28. [Google Scholar] [CrossRef]
- Hossain, M.A.; Islam, S.M.S.; Hasan, M.M. Changes in Soil Properties with Combined Use of Probiotic Cultures and Organic Farming Practices in Degraded Soils of Bangladesh. Sustainability 2023, 15, 4430. [Google Scholar] [CrossRef]
- Zheng, W.; Zeng, S.; Bais, H.; LaManna, J.M.; Hussey, D.S.; Jacobson, D.L.; Jin, Y. Plant Growth-Promoting Rhizobacteria (PGPR) Reduce Evaporation and Increase Soil Water Retention. Water Resour. Res. 2018, 54, 3673–3687. [Google Scholar] [CrossRef]
- Chung, K.-S.; Shin, J.-S.; Lee, J.-H.; Park, S.-E.; Han, H.-S.; Rhee, Y.K.; Cho, C.-W.; Hong, H.-D.; Lee, K.-T. Protective Effect of Exopolysaccharide Fraction from Bacillus subtilis against Dextran Sulfate Sodium-Induced Colitis through Maintenance of Intestinal Barrier and Suppression of Inflammatory Responses. Int. J. Biol. Macromol. 2021, 178, 363–372. [Google Scholar] [CrossRef]
- Yang, H.; Deng, J.; Yuan, Y.; Fan, D.; Zhang, Y.; Zhang, R.; Han, B. Two Novel Exopolysaccharides from Bacillus Amyloliquefaciens C-1: Antioxidation and Effect on Oxidative Stress. Curr. Microbiol. 2015, 70, 298–306. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Shavit, U.; Furman, A. Water Retention Curves of Biofilm-Affected Soils Using Xanthan as an Analogue. Soil Sci. Soc. Am. J. 2012, 76, 61–69. [Google Scholar] [CrossRef]
- Song, M.; Zhang, X.; Yang, J.; Gao, C.; Wei, Y.; Chen, S.; Liesche, J. Arabidopsis Plants Engineered for High Root Sugar Secretion Enhance the Diversity of Soil Microorganisms. Biotechnol. J. 2022, 17, 2100638. [Google Scholar] [CrossRef]
- Adomako, M.O.; Roiloa, S.; Yu, F.-H. Potential Roles of Soil Microorganisms in Regulating the Effect of Soil Nutrient Heterogeneity on Plant Performance. Microorganisms 2022, 10, 2399. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Shi, Y.; Zhang, Y.; Yang, D.; Guo, C. Effects of Plant-Growth-Promoting Rhizobacteria on Soil Bacterial Community, Soil Physicochemical Properties, and Soil Enzyme Activities in the Rhizosphere of Alfalfa under Field Conditions. Diversity 2023, 15, 537. [Google Scholar] [CrossRef]
- Nuccio, E.E.; Starr, E.; Karaoz, U.; Brodie, E.L.; Zhou, J.; Tringe, S.G.; Malmstrom, R.R.; Woyke, T.; Banfield, J.F.; Firestone, M.K.; et al. Niche Differentiation Is Spatially and Temporally Regulated in the Rhizosphere. ISME J. 2020, 14, 999–1014. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Hu, W.; Zheng, J.; Du, F.; Zhang, X. Estimating Soil Organic Carbon Storage and Distribution in a Catchment of Loess Plateau, China. Geoderma 2010, 154, 261–266. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of Plant Growth Promoting Rhizobacteria for Sustainable Development in Agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Dallaire, K.; Skousen, J. Early Tree Growth in Reclaimed Mine Soils in Appalachia USA. Forests 2019, 10, 549. [Google Scholar] [CrossRef]
- Banerjee, S.; van der Heijden, M.G.A. Soil Microbiomes and One Health. Nat. Rev. Microbiol. 2023, 21, 6–20. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial Diversity and Soil Functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Zhong, W.; Bian, B.; Gao, N.; Min, J.; Shi, W.; Lin, X.; Shen, W. Nitrogen Fertilization Induced Changes in Ammonia Oxidation Are Attributable Mostly to Bacteria Rather than Archaea in Greenhouse-Based High N Input Vegetable Soil. Soil Biol. Biochem. 2016, 93, 150–159. [Google Scholar] [CrossRef]
- Duan, Y.; Awasthi, S.K.; Liu, T.; Verma, S.; Wang, Q.; Chen, H.; Ren, X.; Zhang, Z.; Awasthi, M.K. Positive Impact of Biochar Alone and Combined with Bacterial Consortium Amendment on Improvement of Bacterial Community during Cow Manure Composting. Bioresour. Technol. 2019, 280, 79–87. [Google Scholar] [CrossRef]
- Han, G.; Chen, Q.; Zhang, S.; Li, G.; Yi, X.; Feng, C.; Wang, X.; Yu, C.; Lan, J. Biochar Effects on Bacterial Community and Metabolic Pathways in Continuously Cotton-Cropped Soil. J. Soil Sci. Plant Nutr. 2019, 19, 249–261. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the Rhizosphere Microbiome by Biofertilizer Application to Suppress Banana Fusarium Wilt Disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Guo, J.; Liu, W.; Zhu, C.; Luo, G.; Kong, Y.; Ling, N.; Wang, M.; Dai, J.; Shen, Q.; Guo, S. Bacterial Rather than Fungal Community Composition Is Associated with Microbial Activities and Nutrient-Use Efficiencies in a Paddy Soil with Short-Term Organic Amendments. Plant Soil 2018, 424, 335–349. [Google Scholar] [CrossRef]
- Huang, F.; Liu, Z.; Mou, H.; Li, J.; Zhang, P.; Jia, Z. Impact of Farmland Mulching Practices on the Soil Bacterial Community Structure in the Semiarid Area of the Loess Plateau in China. Eur. J. Soil Biol. 2019, 92, 8–15. [Google Scholar] [CrossRef]
- Kim, H.M.; Jung, J.Y.; Yergeau, E.; Hwang, C.Y.; Hinzman, L.; Nam, S.; Hong, S.G.; Kim, O.-S.; Chun, J.; Lee, Y.K. Bacterial Community Structure and Soil Properties of a Subarctic Tundra Soil in Council, Alaska. FEMS Microbiol. Ecol. 2014, 89, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Rui, J.; Mao, Y.; Yannarell, A.; Mackie, R. Dynamics of the Bacterial Community Structure in the Rhizosphere of a Maize Cultivar. Soil Biol. Biochem. 2014, 68, 392–401. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The Microbial Nitrogen-Cycling Network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Spain, A.M.; Krumholz, L.R.; Elshahed, M.S. Abundance, Composition, Diversity and Novelty of Soil Proteobacteria. ISME J. 2009, 3, 992–1000. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Eichorst, S.A.; Trojan, D.; Roux, S.; Herbold, C.; Rattei, T.; Woebken, D. Genomic Insights into the Acidobacteria Reveal Strategies for Their Success in Terrestrial Environments. Environ. Microbiol. 2018, 20, 1041–1063. [Google Scholar] [CrossRef] [PubMed]
- Pajares, S.; Bohannan, B.J.M. Ecology of Nitrogen Fixing, Nitrifying, and Denitrifying Microorganisms in Tropical Forest Soils. Front. Microbiol. 2016, 7, 1045. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, L.; Wen, H.; Zhou, T.; Zhang, T.; Gao, X. 454 Pyrosequencing Analysis of Bacterial Diversity Revealed by a Comparative Study of Soils from Mining Subsidence and Reclamation Areas. J. Microbiol. Biotechnol. 2014, 24, 313–323. [Google Scholar] [CrossRef]
- Loganathachetti, D.S.; Venkatachalam, S.; Jabir, T.; Vipindas, P.V.; Krishnan, K.P. Total Nitrogen Influence Bacterial Community Structure of Active Layer Permafrost across Summer and Winter Seasons in Ny-Ålesund, Svalbard. World J. Microbiol. Biotechnol. 2022, 38, 28. [Google Scholar] [CrossRef] [PubMed]
- Weng, X.; Sui, X.; Liu, Y.; Yang, L.; Zhang, R. Effect of Nitrogen Addition on the Carbon Metabolism of Soil Microorganisms in a Calamagrostis Angustifolia Wetland of the Sanjiang Plain, Northeastern China. Ann. Microbiol. 2022, 72, 18. [Google Scholar] [CrossRef]
- Kong, J.; He, Z.; Chen, L.; Yang, R.; Du, J. Efficiency of Biochar, Nitrogen Addition, and Microbial Agent Amendments in Remediation of Soil Properties and Microbial Community in Qilian Mountains Mine Soils. Ecol. Evol. 2021, 11, 9318–9331. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | ShannonBC | SimpsonBC | ACE | Chao |
|---|---|---|---|---|
| M0B0 | 6.72 ± 0.04 | 0.0024 ± 0.00014 c | 3154.92 ± 18.40 c | 3243.05 ± 18.91 b |
| MB0 | 6.56 ± 0.19 | 0.0030 ± 0.00049 b | 3405.73 ± 56.38 b | 3505.68 ± 58.04 a |
| B20 | 6.80 ± 0.34 | 0.0034 ± 0.00017 a | 3659.33 ± 105.64 a | 3730.09 ± 107.68 a |
| MB20 | 6.70 ± 0.06 | 0.0028 ± 0.00058 b | 3647.77 ± 74.21 a | 3703.99 ± 75.35 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, C.; Lv, C.; Chen, H.; Zhou, J.; Ren, H. Evaluation of Soil Total Nitrogen as an Indicator of Soil Bacterial Community Response to Biochar and Plant Growth-Promoting Rhizobacteria Applications. Agronomy 2024, 14, 428. https://doi.org/10.3390/agronomy14030428
Ding C, Lv C, Chen H, Zhou J, Ren H. Evaluation of Soil Total Nitrogen as an Indicator of Soil Bacterial Community Response to Biochar and Plant Growth-Promoting Rhizobacteria Applications. Agronomy. 2024; 14(3):428. https://doi.org/10.3390/agronomy14030428
Chicago/Turabian StyleDing, Chenke, Chengqun Lv, Hualin Chen, Jiangmin Zhou, and Han Ren. 2024. "Evaluation of Soil Total Nitrogen as an Indicator of Soil Bacterial Community Response to Biochar and Plant Growth-Promoting Rhizobacteria Applications" Agronomy 14, no. 3: 428. https://doi.org/10.3390/agronomy14030428
APA StyleDing, C., Lv, C., Chen, H., Zhou, J., & Ren, H. (2024). Evaluation of Soil Total Nitrogen as an Indicator of Soil Bacterial Community Response to Biochar and Plant Growth-Promoting Rhizobacteria Applications. Agronomy, 14(3), 428. https://doi.org/10.3390/agronomy14030428