
Physiological and Biochemical Background of Deacclimation in Plants, with Special Attention Being Paid to Crops: A Minireview



Abstract
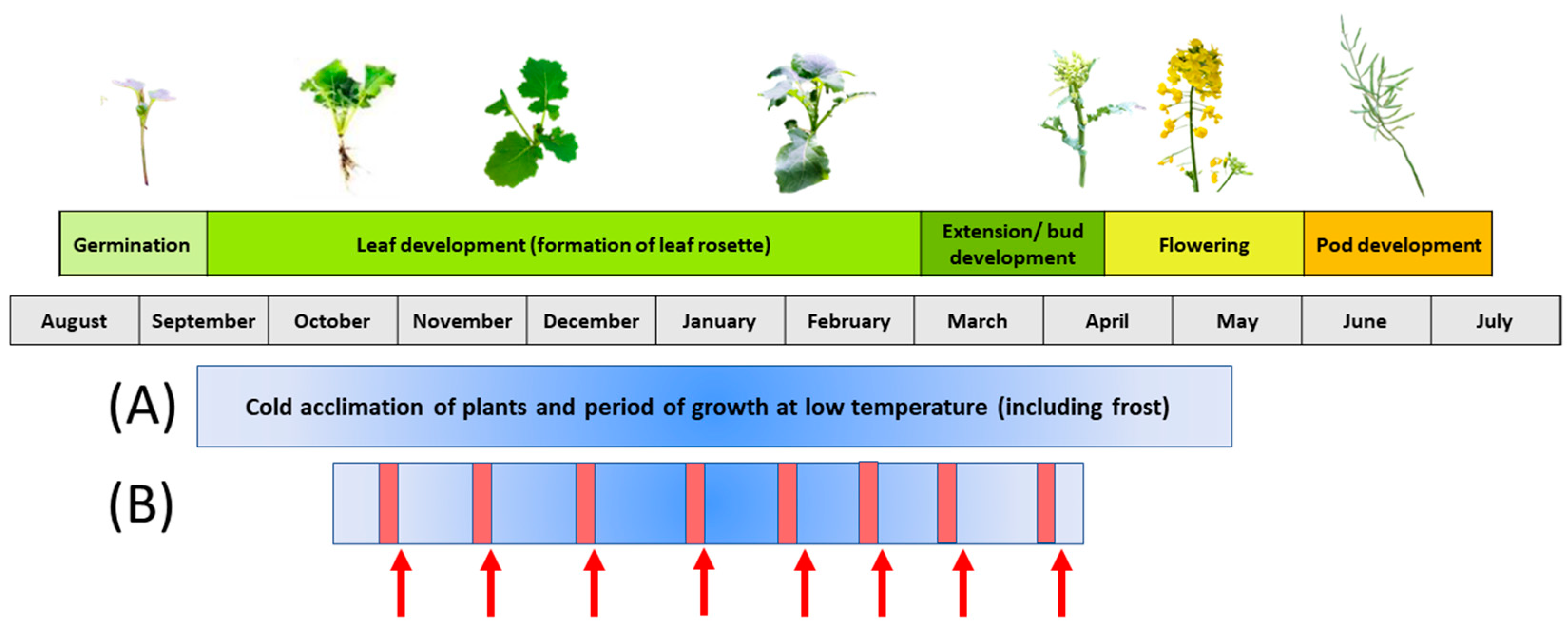
1. Introduction—Significance of Cold Acclimation and Deacclimation for Crops
2. Factors That Influence the Phenomenon of Deacclimation
3. The Physiological-Biochemical Changes That Occur in Plants during Deacclimation
3.1. The Cell Walls and Cell Membranes
3.2. Soluble Sugar Concentration and Water Management
3.3. Accumulation of Selected Proteins
3.4. Hormonal Balance
3.5. Photosynthesis
3.6. Other Changes
4. How Could the Results of Deacclimation Studies Be Used in Practice?
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPCC Summary for Policymakers: Synthesis Report. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2023; pp. 1–34. [Google Scholar]
- Nauka o Klimacie. Available online: https://naukaoklimacie.pl/aktualnosci/zmiana-klimatu-w-polsce-na-mapkach-468/ (accessed on 28 January 2024).
- Chouard, P. Vernalization and its Relations to Dormancy. Annu. Rev. Plant Physiol. 1960, 11, 191–238. [Google Scholar] [CrossRef]
- Rapacz, M.; Janowiak, F. Physiological effects of winter rape (Brassica napus var. oleifera) prehardening to frost. I. Frost resistance and photosynthesis during cold acclimation. J. Agron. Crop Sci. 1998, 181, 13–20. [Google Scholar] [CrossRef]
- Sasaki, H.; Ichimura, K.; Oda, M. Changes in sugar content during cold acclimation and deacclimation of cabbage seedlings. Ann. Bot. 1996, 78, 365–369. [Google Scholar] [CrossRef]
- Uemura, M.; Joseph, R.A.; Steponkus, P.L. Cold acclimation of Arabidopsis thaliana: Effect on plasma membrane lipid composition and freeze-induced lesions. Plant Physiol. 1995, 109, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Filek, M.; Rudolphi-Skórska, E.; Sieprawska, A.; Kvasnica, M.; Janeczko, A. Regulation of the membrane structure by brassinosteroids and progesterone in winter wheat seedlings exposed to low temperature. Steroids 2017, 128, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Prášil, I.T.; Vítámvás, P.; Dobrev, P.; Motyka, V.; Floková, K.; Novák, O.; Turečková, V.; Rolčik, J.; Pešek, B.; et al. Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter Samanta and spring Sandra. J. Plant Physiol. 2012, 169, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Wang, L.J.; Pan, Q.H.; Wang, Y.Z.; Zhan, J.C.; Huang, W.D. Accumulation and subcellular localization of heat shock proteins in young grape leaves during cross-adaptation to temperature stresses. Sci. Hortic. 2008, 117, 231–240. [Google Scholar] [CrossRef]
- Sadura, I.; Libik-Konieczny, M.; Jurczyk, B.; Gruszka, D.; Janeczko, A. HSP transcript and protein accumulation in brassinosteroid barley mutants acclimated to low and high temperatures. Int. J. Mol. Sci. 2020, 21, 1889. [Google Scholar] [CrossRef]
- Giorni, E.; Crosatti, C.; Baldi, P.; Grossi, M.; Marè, C.; Stanca, A.M.; Cattivelli, L. Cold-regulated gene expression during winter in frost tolerant and frost susceptible barley cultivars grown under field conditions. Euphytica 1999, 106, 149–157. [Google Scholar] [CrossRef]
- Stitt, M.; Hurry, V. A plant for all seasons: Alterations in photosynthetic carbon metabolism during cold acclimation in Arabidopsis. Curr. Opin. Plant Biol. 2002, 5, 199–206. [Google Scholar] [CrossRef]
- Stachurska, J.; Rys, M.; Pociecha, E.; Kalaji, H.M.; Dąbrowski, P.; Oklestkova, J.; Jurczyk, B.; Janeczko, A. Deacclimation-Induced Changes of Photosynthetic Efficiency, Brassinosteroid Homeostasis and BRI1 Expression in Winter Oilseed Rape (Brassica napus L.)—Relation to Frost Tolerance. Int. J. Mol. Sci. 2022, 23, 5224. [Google Scholar] [CrossRef]
- Nick, P. Microtubules as Sensors for Abiotic Stimuli. In Plant Microtubules; Plant Cell Monographs; Nick, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 11, pp. 175–203. [Google Scholar]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Rapacz, M.; Jurczyk, B.; Sasal, M. Deacclimation may be crucial for winter survival of cereals under warming climate. Plant Sci. 2017, 256, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Vyse, K.; Pagter, M.; Zuther, E.; Hincha, D.K. Deacclimation after cold acclimation—A crucial, but widely neglected part of plant winter survival. J. Exp. Bot. 2019, 70, 4595–4604. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Kasuga, J. Bark cells and xylem cells in Japanese white birch twigs initiate deacclimation at different temperatures. Cryobiology 2018, 80, 96–100. [Google Scholar] [CrossRef]
- Yoshida, S. Reverse changes in plasma membrane properties upon deacclimation of mulberry trees (Morus bombysis Koidz). J. Sericult. Sci. Jpn. 1987, 56, 440. [Google Scholar] [CrossRef]
- Tinus, R.W.; Burr, K.E.; Atzmon, N.; Riov, J. Relationship between carbohydrate concentration and root growth potential in coniferous seedlings from three climates during cold hardening and dehardening. Tree Physiol. 2000, 20, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- López, D.; Larama, G.; Sáez, P.L.; Bravo, L.A. Transcriptome Analysis of Diurnal and Nocturnal-Warmed Plants, the Molecular Mechanism Underlying Cold Deacclimation Response in Deschampsia antarctica. Int. J. Mol. Sci. 2023, 24, 11211. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.; DaCosta, M.; Bertrand, A.; Castonguay, Y.; Ebdon, J.S. Comparative assessment of metabolic responses to cold acclimation and deacclimation in annual bluegrass and creeping bentgrass. Environ. Exp. Bot. 2014, 106, 197–206. [Google Scholar] [CrossRef]
- Oono, Y.; Seki, M.; Satou, M.; Iida, K.; Akiyama, K.; Sakurai, T.; Fujita, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Monitoring expression profiles of Arabidopsis genes during cold acclimation and deacclimation using DNA microarrays. Funct. Integr. Genom. 2006, 6, 212–234. [Google Scholar] [CrossRef]
- Pagter, M.; Alpers, J.; Erban, A.; Kopka, J.; Zuther, E.; Hincha, D.K. Rapid transcriptional and metabolic regulation of the deacclimation process in cold acclimated Arabidopsis thaliana. BMC Genom. 2017, 18, 731. [Google Scholar] [CrossRef] [PubMed]
- Rys, M.; Pociecha, E.; Oliwa, J.; Ostrowska, A.; Jurczyk, B.; Saja, D.; Janeczko, A. Deacclimation of winter oilseed rape-insight into physiological changes. Agronomy 2020, 10, 1565. [Google Scholar] [CrossRef]
- Miki, Y.; Takahashi, D.; Kawamura, Y.; Uemura, M. Temporal proteomics of Arabidopsis plasma membrane during cold- and de-acclimation. J. Proteom. 2019, 197, 71–81. [Google Scholar] [CrossRef]
- Chen, H.-H.; Li, P.H. Characteristics of Cold Acclimation and Deacclimation in Tuber-bearing Solanum Species. Plant Physiol. 1980, 65, 1146–1148. [Google Scholar] [CrossRef] [PubMed]
- Fürtauer, L.; Weiszmann, J.; Weckwerth, W.; Nägele, T. Dynamics of plant metabolism during cold acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Ichmirua, K.; Imada, S.; Oda, M. Loss of Freezing Tolerance associated with decrease in sugar concentrations by short-term deacclimation in cabbage seedlings. J. Japan. Soc. Hort. Sci. 2001, 70, 294–298. [Google Scholar] [CrossRef][Green Version]
- Rapacz, M. Cold-deacclimation of oilseed rape (Brassica napus var. oleifera) in response to fluctuating temperatures and photoperiod. Ann. Bot. 2002, 89, 543–549. [Google Scholar] [CrossRef]
- Burbulis, N.; Jonytiene, V.; Kupriene, R.; Blinstrubiene, A.; Liakas, V. Biochemical and physiological factors related to cold de-acclimation and re-acclimation in rapeseed shoots in vitro. J. Food Agric. Environ. 2011, 9, 483–487. [Google Scholar]
- Vega, S.E.; Palta, J.P.; Bamberg, J.B. Variability in the rate of cold acclimation and deacclimation among tuber-bearing Solanum (potato) species. J. Am. Soc. Hortic. Sci. 2000, 125, 205–211. [Google Scholar] [CrossRef]
- Pagter, M.; Hausman, J.F.; Arora, R. Deacclimation kinetics and carbohydrate changes in stem tissues of Hydrangea in response to an experimental warm spell. Plant Sci. 2011, 180, 140–148. [Google Scholar] [CrossRef]
- Pociecha, E.; Janeczko, A.; Dziurka, M.; Gruszka, D. Disturbances in the Biosynthesis or Signalling of Brassinosteroids That Are Caused by Mutations in the HvDWARF, HvCPD and HvBRI1 Genes Increase the Tolerance of Barley to the Deacclimation Process. J. Plant Growth Regul. 2020, 39, 1625–1637. [Google Scholar] [CrossRef]
- Wójcik-Jagła, M.; Rapacz, M. Freezing tolerance and tolerance to de-acclimation of European accessions of winter and facultative barley. Sci. Rep. 2023, 13, 19931. [Google Scholar] [CrossRef] [PubMed]
- Taulavuori, K.M.J.; Taulavuori, E.B.; Skre, O.; Nilsen, J.; Igeland, B.; Laine, K.M. Dehardening of mountain birch (Betula pubescens ssp. czerepanovii) ecotypes at elevated winter temperatures. New Phytol. 2004, 162, 427–436. [Google Scholar] [CrossRef]
- Byun, Y.J.; Koo, M.Y.; Joo, H.J.; Ha-Lee, Y.M.; Lee, D.H. Comparative analysis of gene expression under cold acclimation, deacclimation and reacclimation in Arabidopsis. Physiol. Plant. 2014, 152, 256–274. [Google Scholar] [CrossRef] [PubMed]
- Zuther, E.; Juszczak, I.; Ping Lee, Y.; Baier, M.; Hincha, D.K. Time-dependent deacclimation after cold acclimation in Arabidopsis thaliana accessions. Sci. Rep. 2015, 5, 12199. [Google Scholar] [CrossRef] [PubMed]
- Kutsuno, T.; Chowhan, S.; Kotake, T.; Takahashi, D. Temporal cell wall changes during cold acclimation and deacclimation and their potential involvement in freezing tolerance and growth. Physiol. Plant. 2023, 175, e13837. [Google Scholar] [CrossRef] [PubMed]
- Rapacz, M. Regulation of frost resistance during cold de-acclimation and re-acclimation in oilseed rape. A possible role of PSII redox state. Physiol. Plant. 2002, 115, 236–243. [Google Scholar] [CrossRef]
- Cen, W.; Liu, J.; Lu, S.; Jia, P.; Yu, K.; Han, Y.; Li, R.; Luo, J. Comparative proteomic analysis of QTL CTS-12 derived from wild rice (Oryza rufipogon Griff.), in the regulation of cold acclimation and de-acclimation of rice (Oryza sativa L.) in response to severe chilling stress. BMC Plant Biol. 2018, 18, 163. [Google Scholar] [CrossRef]
- Vaitkevičiūtė, G.; Aleliūnas, A.; Gibon, Y.; Armonienė, R. The effect of cold acclimation, deacclimation and reacclimation on metabolite profiles and freezing tolerance in winter wheat. Front. Plant Sci. 2022, 13, 959118. [Google Scholar] [CrossRef]
- Rapacz, M.; Macko-Podgórni, A.; Jurczyk, B.; Kuchar, L. Modeling wheat and triticale winter hardiness under current and predicted winter scenarios for Central Europe: A focus on deacclimation. Agric. For. Meteorol. 2022, 313, 108739. [Google Scholar] [CrossRef]
- Solecka, D.; Zebrowski, J.; Kacperska, A. Are pectins involved in cold acclimation and de-acclimation of winter oil-seed rape plants? Ann. Bot. 2008, 101, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Ford, R.C.; Barber, J. Incorporation of sterol into chloroplast thylakoid membranes and its effect on fluidity and function. Planta 1983, 158, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S. The role of α-tocopherol in plant stress tolerance. J. Plant Physiol. 2005, 162, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Iivonen, S.; Saranpää, P.; Sutinen, M.L.; Vapaavuori, E. Effects of temperature and nutrient availability on plasma membrane lipid composition in Scots pine roots during growth initiation. Tree Physiol. 2004, 24, 437–446. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rys, M.; Stachurska, J.; Rudolphi-Szydło, E.; Dziurka, M.; Waligórski, P.; Filek, M.; Janeczko, A. Does deacclimation reverse the changes in structural/physicochemical properties of the chloroplast membranes that are induced by cold acclimation in oilseed rape? Plant Physiol. Biochem. 2024, in press. [Google Scholar]
- Niki, E.; Abe, K. CHAPTER 1: Vitamin E: Structure, Properties and Functions. In Food Chemistry, Function and Analysis; Royal Society of Chemistry: Cambridge, UK, 2019; pp. 1–11. [Google Scholar] [CrossRef]
- Havaux, M.; Eymery, F.; Porfirova, S.; Rey, P.; Dörmann, P. Vitamin E protects against photoinhibition and photooxidative stress in Arabidopsis thaliana. Plant Cell 2005, 17, 3451–3469. [Google Scholar] [CrossRef] [PubMed]
- Krieger-Liszkay, A.; Trebst, A. Tocopherol is the scavenger of singlet oxygen produced by the triplet states of chlorophyll in the PSII reaction centre. J. Exp. Bot. 2006, 57, 1677–1684. [Google Scholar] [CrossRef]
- Hincha, D.K. Effects of α-tocopherol (vitamin E) on the stability and lipid dynamics of model membranes mimicking the lipid composition of plant chloroplast membranes. FEBS Lett. 2008, 582, 3687–3692. [Google Scholar] [CrossRef]
- Gzyl-Malcher, B.; Zembala, M.; Filek, M. Effect of tocopherol on surface properties of plastid lipids originating from wheat calli cultivated in cadmium presence. Chem. Phys. Lipids 2010, 163, 74–81. [Google Scholar] [CrossRef]
- Liu, W.; Yu, K.; He, T.; Li, F.; Zhang, D.; Liu, J. The low temperature induced physiological responses of Avena nuda L., a cold-tolerant plant species. Sci. World J. 2013, 2013, 658793. [Google Scholar] [CrossRef]
- Janeczko, A.; Dziurka, M.; Pociecha, E. Increased leaf tocopherol and β-carotene content is associated with the tolerance of winter wheat cultivars to frost. J. Agron. Crop Sci. 2018, 204, 594–602. [Google Scholar] [CrossRef]
- Svenning, M.M.; Røsnes, K.; Junttila, O. Frost tolerance and biochemical changes during hardening and dehardening in contrasting white clover populations. Physiol. Plant. 1997, 101, 31–37. [Google Scholar] [CrossRef]
- Shin, H.; Oh, Y.; Kim, D. Differences in cold hardiness, carbohydrates, dehydrins and related gene expressions under an experimental deacclimation and reacclimation in Prunus persica. Physiol. Plant. 2015, 154, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Winde, J.; Andersen, U.B.; Kjaer, K.H.; Pagter, M. Variation in freezing tolerance, water content and carbohydrate metabolism of floral buds during deacclimation of contrasting blackcurrant cultivars. Acta Physiol. Plant. 2017, 39, 201. [Google Scholar] [CrossRef]
- Ogren, E. Premature Dehardening in Vaccinium myrtillus during a Mild Winter: A Cause for Winter Dieback? Funct. Ecol. 1996, 10, 724–732. [Google Scholar] [CrossRef]
- Gerbeau, P.; Amodeo, G.; Henzler, T.; Santoni, V.; Ripoche, P.; Maurel, C. The water permeability of Arabidopsis plasma membrane is regulated by divalent cations and pH. Plant J. 2002, 30, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Hao, X.; Wang, L.; Li, N.; Huang, J.; Zeng, J.; Yang, Y.; Wang, X. iTRAQ-based quantitative proteomic analysis of tea plant (Camellia sinensis (L.) O. Kuntze) during cold acclimation and de-acclimation procedures. Beverage Plant Res. 2023, 2023, 16. [Google Scholar] [CrossRef]
- Trischuk, R.G.; Schilling, B.S.; Low, N.H.; Gray, G.R.; Gusta, L.V. Cold acclimation, de-acclimation and re-acclimation of spring canola, winter canola and winter wheat: The role of carbohydrates, cold-induced stress proteins and vernalization. Environ. Exp. Bot. 2014, 106, 156–163. [Google Scholar] [CrossRef]
- Stachurska, J.; Sadura, I.; Rys, M.; Dziurka, M.; Janeczko, A. Insight into Hormonal Homeostasis and the Accumulation of Selected Heat Shock Proteins in Cold Acclimated and Deacclimated Winter Oilseed Rape. Agriculture 2023, 13, 641. [Google Scholar] [CrossRef]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Park, C.J.; Seo, Y.S. Heat shock proteins: A review of the molecular chaperones for plant immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Kontunen-Soppela, S.; Taulavuori, K.; Taulavuori, E.; Lähdesmäki, P.; Laine, K. Soluble proteins and dehydrins in nitrogen-fertilized Scots pine seedlings during deacclimation and the onset of growth. Physiol. Plant. 2000, 109, 404–409. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Prášil, I.T. The role of dehydrins in plant response to cold. Biol. Plant. 2007, 51, 601–617. [Google Scholar] [CrossRef]
- Kalberer, S.R.; Wisniewski, M.; Arora, R. Deacclimation and reacclimation of cold-hardy plants: Current understanding and emerging concepts. Plant Sci. 2006, 171, 3–16. [Google Scholar] [CrossRef]
- Kovaleski, A.P.; Londo, J.P. Tempo of gene regulation in wild and cultivated Vitis species shows coordination between cold deacclimation and budbreak. Plant Sci. 2019, 287, 110178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ervin, E.H.; Waltz, C.; Murphy, T. Metabolic changes during cold acclimation and deacclimation in five bermudagrass varieties: II. Cytokinin and abscisic acid metabolism. Crop Sci. 2011, 51, 847–853. [Google Scholar] [CrossRef]
- Janeczko, A.; Pociecha, E.; Dziurka, M.; Jurczyk, B.; Libik-Konieczny, M.; Oklestkova, J.; Novák, O.; Pilarska, M.; Filek, M.; Rudolphi-Skórska, E.; et al. Changes in content of steroid regulators during cold hardening of winter wheat—Steroid physiological/biochemical activity and impact on frost tolerance. Plant Physiol. Biochem. 2019, 139, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Stachurska, J.; Sadura, I.; Rys, M.; Jurczyk, B.; Oklestkova, J.; Janeczko, A. The role of brassinosteroids in the cold acclimation of oilseed rape. In Proceedings of the Conference “The XXX International Symposium Molecular and Physiological Aspects of Regulatory Processes in the Organism”, Jagiellonian University, Krakow, Poland, 14 October 2023. [Google Scholar]
- Janeczko, A.; Oklestkova, J. Analysis of Brassinosteroids in Crowns of Winter Wheat. Unpublished Data. 2019. [Google Scholar]
- Pukacki, P.; Veselovsky, V.A.; Veselova, T.V. Effect of Cold Deacclimation on Delayed Fluorescence of Spruce Needles. Z. Pflanzenphysiol. 1983, 109, 267–273. [Google Scholar] [CrossRef]
- Repo, T.; Leinonen, I.; Wang, K.Y.; Hänninen, H. Relation between photosynthetic capacity and cold hardiness in Scots pine. Physiol. Plant. 2006, 126, 224–231. [Google Scholar] [CrossRef]
- Rapacz, M.; Hura, K. The pattern of changes in photosynthetic apparatus in response to cold acclimation and de-acclimation in two contrasting cultivars of oilseed rape. Photosynthetica 2002, 40, 63–69. [Google Scholar] [CrossRef]
- Stachurska, J. Nieinwazyjne metody pomiarowe do oceny występowania zjawiska rozhartowania u rzepaku [Non-invasive measurements methods to assess the deacclimation process in oilseed rape]. In Proceedings of the Conference “X Forum Młodych Naukowców Nauki Przyrodnicze i Medyczne”, Lublin, Poland, 1–2 December 2023. [Google Scholar]
- Horvath, D.P.; Zhang, J.; Chao, W.S.; Mandal, A.; Rahman, M.; Anderson, J.V. Genome-wide association studies and transcriptome changes during acclimation and deacclimation in divergent Brassica napus varieties. Int. J. Mol. Sci. 2020, 21, 9148. [Google Scholar] [CrossRef] [PubMed]
- Pukacki, P.M.; Kamińska-Rożek, E. Reactive species, antioxidants and cold tolerance during deacclimation of Picea abies populations. Acta Physiol. Plant. 2013, 35, 129–138. [Google Scholar] [CrossRef]
- Vaitkevičiūtė, G.; Aleliūnas, A.; Gibon, Y.; Armonienė, R. Comparative Analysis of Antioxidant Accumulation under Cold Acclimation, Deacclimation and Reacclimation in Winter Wheat. Plants 2022, 11, 2818. [Google Scholar] [CrossRef] [PubMed]
- Saadati, S.; Baninasab, B.; Mobli, M.; Gholami, M. Cold tolerance in olive leaves of three cultivars related to some physiological parameters during cold acclimation and de-acclimation stages. J. Agric. Sci. Technol. 2020, 22, 1313–1326. [Google Scholar]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Natural variation in flavonol and anthocyanin metabolism during cold acclimation in Arabidopsis thaliana accessions. Plant Cell Environ. 2015, 38, 1658–1672. [Google Scholar] [CrossRef] [PubMed]
- Christie, P.J.; Alfenito, M.R.; Walbot, V. Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: Enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. 2005, 105, 319–330. [Google Scholar] [CrossRef]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Zhang, B.; Hu, Z.; Zhang, Y.; Li, Y.; Zhou, S.; Chen, G. A putative functional MYB transcription factor induced by low temperature regulates anthocyanin biosynthesis in purple kale (Brassica oleracea var. acephala f. tricolor). Plant Cell Rep. 2012, 31, 281–289. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Jonytiene, V.; Burbulis, N.; Kupriene, R.; Blinstrubiene, A. Effect of exogenous proline and de-acclimation treatment on cold tolerance in Brassica napus shoots cultured in vitro. J. Food Agric. Environ. 2012, 10, 327–330. [Google Scholar]
- Kubacka-Zebalska, M.; Kacperska, A. Low temperature-induced modifications of cell wall content and polysaccharide composition in leaves of winter oilseed rape (Brassica napus L. var. oleifera L.). Plant Sci. 1999, 148, 59–67. [Google Scholar] [CrossRef]
- Hannah, M.A.; Heyer, A.G.; Hincha, D.K. A global survey of gene regulation during cold acclimation in Arabidopsis thaliana. PLoS Genet. 2005, 1, e26. [Google Scholar] [CrossRef]
- Rowland, L.J.; Ogden, E.L.; Ehlenfeldt, M.K.; Vinyard, B. Cold hardiness, deacclimation kinetics, and bud development among 12 diverse blueberry genotypes under field conditions. J. Am. Soc. Hortic. Sci. 2005, 130, 508–514. [Google Scholar] [CrossRef]
- Qian, J.; Zhou, J.; Gong, R.; Liu, Y.; Zhang, G. Freezing resistance evaluation of rose stems during frost dehardening using electrical impedance tomography. BMC Plant Biol. 2021, 21, 199. [Google Scholar] [CrossRef] [PubMed]
- Gusta, L.V.; Fowler, D.B. Effects of temperature on dehardening and rehardening of winter cereals. Can. J. Plant Sci. 1976, 56, 673–678. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Arabidopsis thaliana | Oilseed Rape | |||
|---|---|---|---|---|
| Cold-Acclimation | Deacclimation | Cold-Acclimation | Deacclimation | |
| Cell walls | Down-regulation of the genes encoding cell-wall-related proteins such as putative xyloglucan endotransglycosylase, xylosidase arabinogalactan protein, and xylosidase [23] | Up-regulation of the genes encoding cell-wall-related proteins such as putative xyloglucan endotransglycosylase, xylosidase, xylose isomerase, pectinesterase, and arabinogalactan protein [23] | The content of pectin in the cell walls increased [44] Higher levels of non-covalently bound pectins and an increased content of galactose, arabinose, and glucose in the pectins and of galactose and arabinose in the hemicelluloses [89] | The content of pectin in the cell walls decreased to a level similar to that in the control plants [44] |
| Suppression of the cell wall-related genes [37] | Reactivation of the cell-wall-related genes [37] | |||
| Increase in the arabinogalactan protein content [39] | Decrease in the arabinogalactan protein content [39] | |||
| Cell membranes | Lower electrolyte leakage (membrane permeability) after the frost test [26] | Higher electrolyte leakage (membrane permeability) after the frost tests [26] | Increased ratio of the unsaturated to saturated fatty acids [48] | Decreased ratio in the unsaturated to saturated fatty acids [48] |
| Increase in membrane fluidity [6] | ||||
| Sugars | Increased sugar content [38] | Decreased sugar content [38] | Increased sugar content [25] | Decreased sugar content [25] |
| Increased starch content [39] | Decreased starch content [39] | |||
| Downregulation of the genes associated with carbohydrate metabolism (e.g., β-galactosidase, sucrose synthase) [23] | Upregulation of the genes associated with carbohydrate metabolism (e.g., β-galactosidase, sucrose synthase) [23] | |||
| Water management | Down-regulation of the genes encoding the water channel proteins (such as tonoplast intrinsic protein gamma (TIP)) [23] | Up-regulation of the genes encoding water channel proteins (such as tonoplast intrinsic protein gamma (TIP)) [23] | Decreased osmotic potential, decreased relative water content (RWC), increased accumulation of aquaporin (BnPIP1) [25] | Increased osmotic potential, increased relative water content (RWC) in one cultivar, decreased accumulation of aquaporin BnPIP1 [25] |
| Proteins | Increased COR expression [12] | Reduced amount of COR transcripts [38] | Increased accumulation of heat shock proteins (HSP) [63] | Decreased accumulation in the heat shock proteins (HSP) [63] |
| Increased accumulation of the cell structure-related proteins and decrease in protein synthesis, destination and storage-related proteins [26] | Reversal of the changes induced by cold in the majority of the protein accumulations (e.g., in the protein synthesis, destination and storage-related proteins, decrease in cell structure-related proteins [26] | Increased accumulation of dehydrins (DHNs) [62] | Decreased accumulation in dehydrins (DHNs) [62] | |
| Increased accumulation of the cold-regulated protein (COR78) [62] | Decreased accumulation in cold-regulated protein (COR78) [62] | |||
| Photosynthesis | Suppression of the photosynthesis-related genes encoding the D2 subunit of the PSII complex [37] | Reactivation of the photosynthesis-related genes encoding the D2 subunit of the PSII complex [37] | Decreased maximum quantum yield of the PSII photochemistry (Fv/Fm) [25] | Increased maximum quantum yield of the PSII photochemistry (Fv/Fm) [25] |
| Inhibition of photosynthesis (reactions of the dark phase) and increase in sucrose-phosphate synthase (SPS) activity [12] | Increased expression of the genes involved in the light reactions of photosynthesis [37] | Limited intensity of the light reactions of photosynthesis [13] | Intensification of the light reactions of photosynthesis [13] | |
| Down-regulation of the photosynthesis-related protein genes, e.g., encoding a small subunit of RuBisCO [23] | Up-regulation of the photosynthesis-related protein genes, e.g., encoding a small subunit of RuBisCO [23] | Increased activity of RuBPCO and SPS [76] | Decreased activity of RuBPCO and SPS [76] | |
| Unchanged or decreased intensity of CO2 assimilation (PN) (cultivar-dependency) [25,77] | Decreased or increased intensity of photosynthesis (CO2 assimilation, PN) (probably dependent by additional factors) [25,77] | |||
| Hormonal homeostasis | Down-regulation of the genes involved in auxin and gibberellin metabolism, down-regulation of the BR biosynthesis pathway [90] | Increased expression of the genes associated with growth-promoting hormones (auxins and gibberellins) and an increased expression of the genes associated with BRs [24] | Increased ABA content and a decrease in growth-promoting hormones [63] | Decreased ABA content and an increase in growth-promoting hormones [63] |
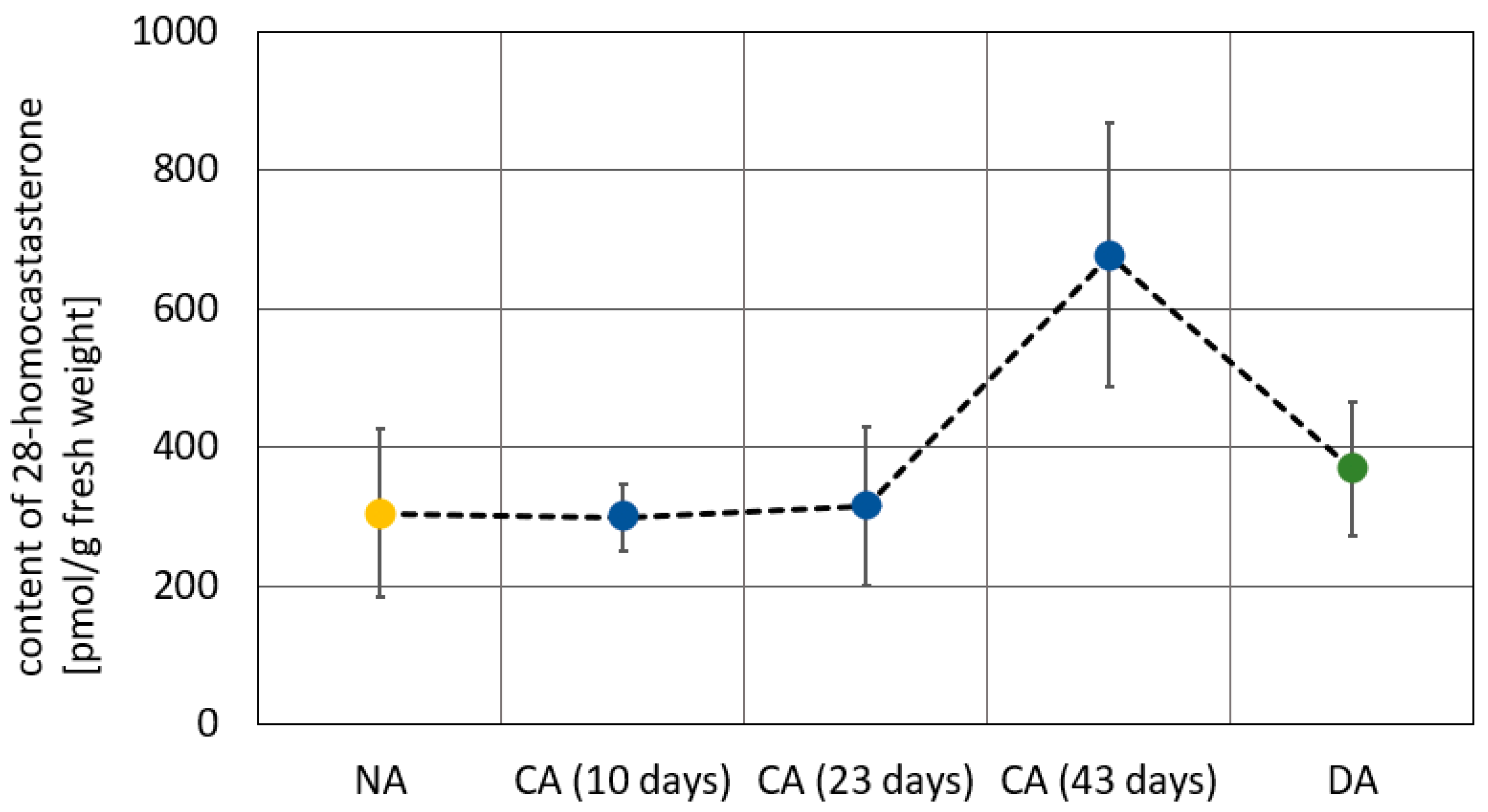
| Tendency to increase the content of brassinosteroids (dependent on the cultivar and on the type of analyzed steroid) [13] | Tendency to decrease the content of brassinosteroids (dependent on the cultivar and on the type of analyzed steroid) [13] | |||
| Decrease in the accumulation of the brassinosteroid receptor (BRI1) [72] | Increase in the accumulation of the brassinosteroid receptor (BRI1) [72] | |||
| Other effects | Increased level of proline [38] | Decreased level of proline [38] | Increased level of proline [88] | Decreased level of proline [88] |
| Increased anthocyanin content [25] | Decreased anthocyanin content [25] | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stachurska, J.; Janeczko, A. Physiological and Biochemical Background of Deacclimation in Plants, with Special Attention Being Paid to Crops: A Minireview. Agronomy 2024, 14, 419. https://doi.org/10.3390/agronomy14030419
Stachurska J, Janeczko A. Physiological and Biochemical Background of Deacclimation in Plants, with Special Attention Being Paid to Crops: A Minireview. Agronomy. 2024; 14(3):419. https://doi.org/10.3390/agronomy14030419
Chicago/Turabian StyleStachurska, Julia, and Anna Janeczko. 2024. "Physiological and Biochemical Background of Deacclimation in Plants, with Special Attention Being Paid to Crops: A Minireview" Agronomy 14, no. 3: 419. https://doi.org/10.3390/agronomy14030419
APA StyleStachurska, J., & Janeczko, A. (2024). Physiological and Biochemical Background of Deacclimation in Plants, with Special Attention Being Paid to Crops: A Minireview. Agronomy, 14(3), 419. https://doi.org/10.3390/agronomy14030419