
Optimized Tomato Production in Chinese Solar Greenhouses: The Impact of an East–West Orientation and Wide Row Spacing


Abstract
1. Introduction
2. Materials and Methods
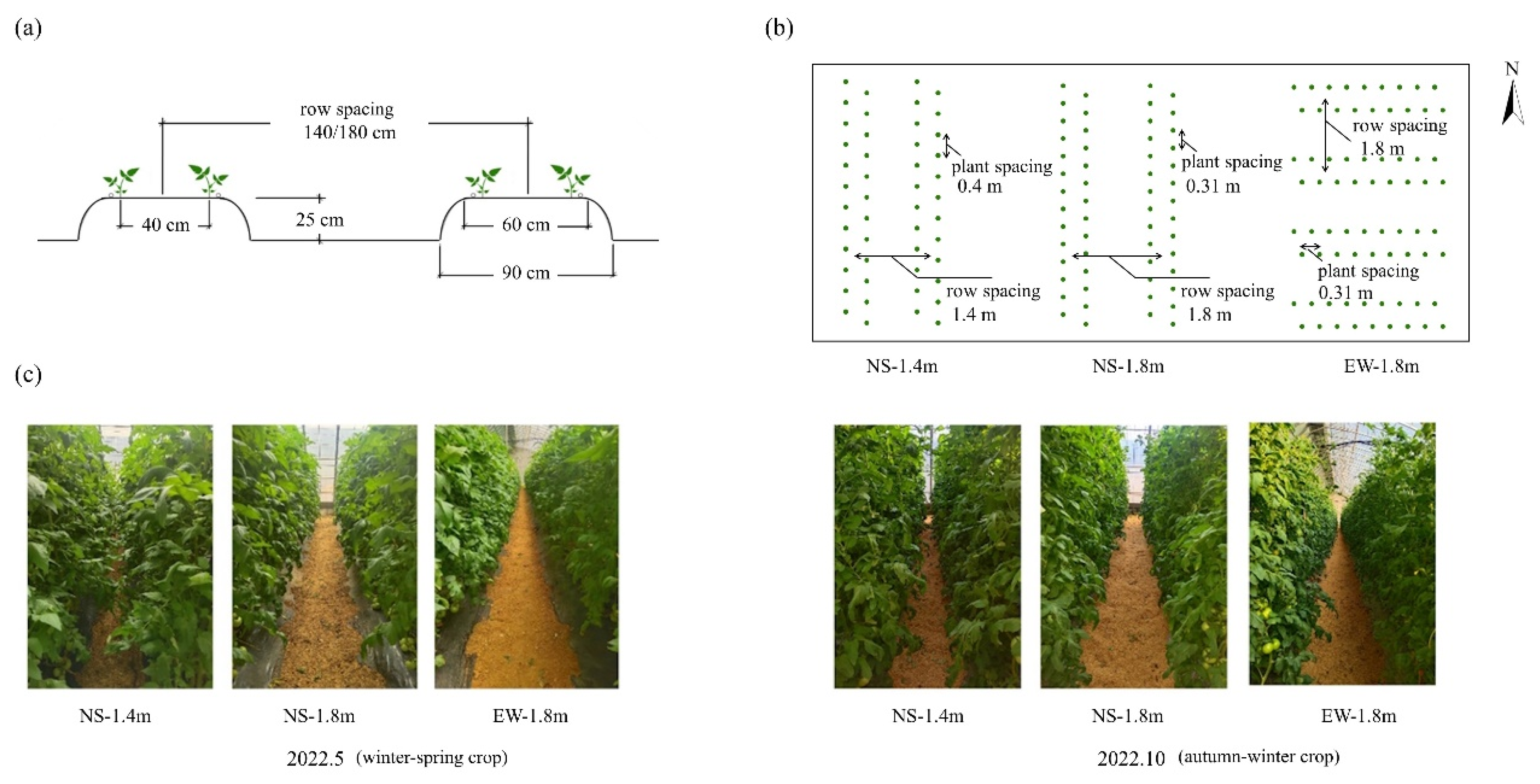
2.1. Experiment Material and Experiment Design
2.2. Plant Measurements
2.2.1. Morphology and Growth
2.2.2. Gas Exchange Parameter
2.2.3. Yield and Quality
2.3. Model Description
2.3.1. Solar Greenhouse Module
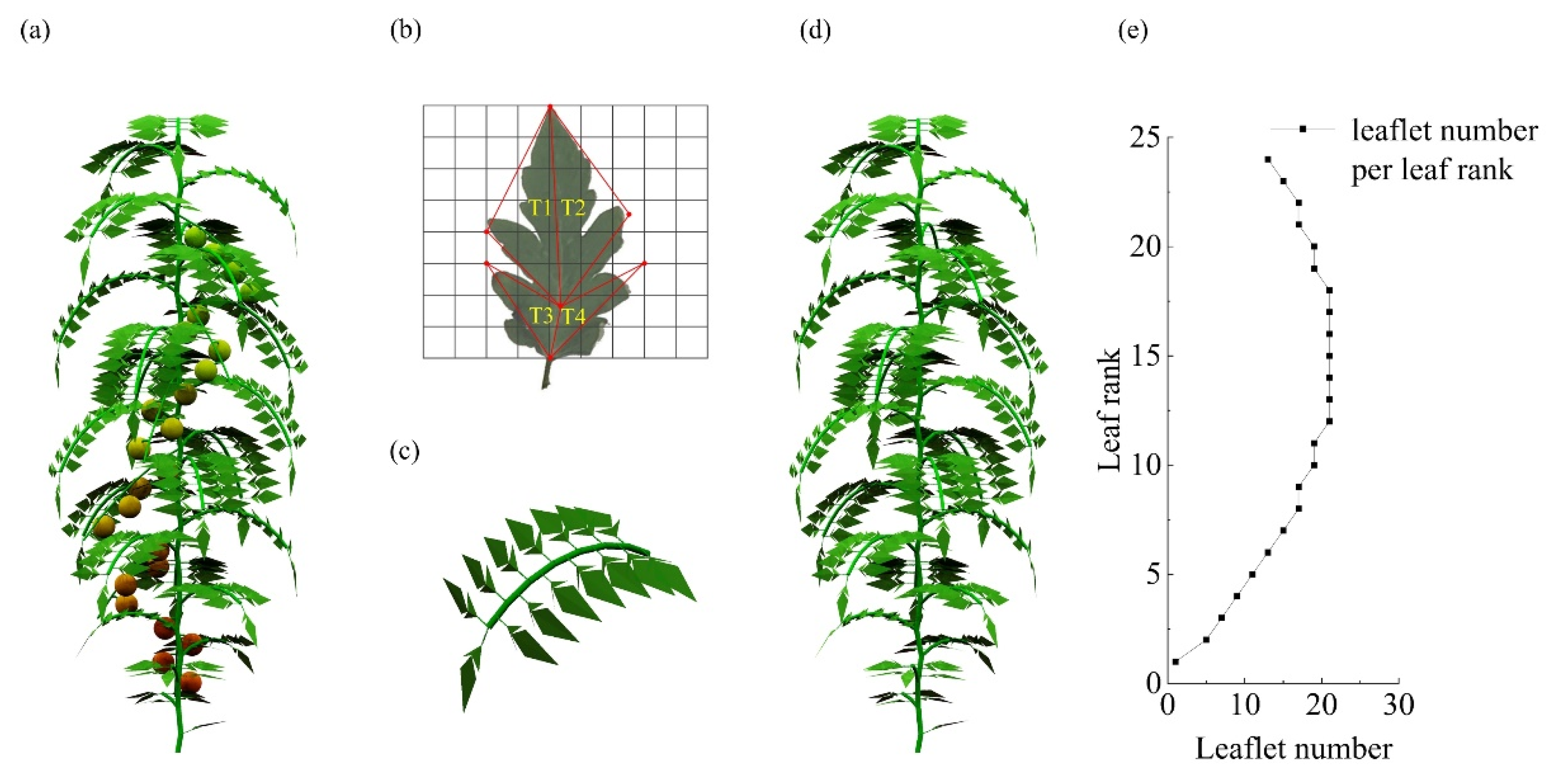
2.3.2. Tomato Plant Module
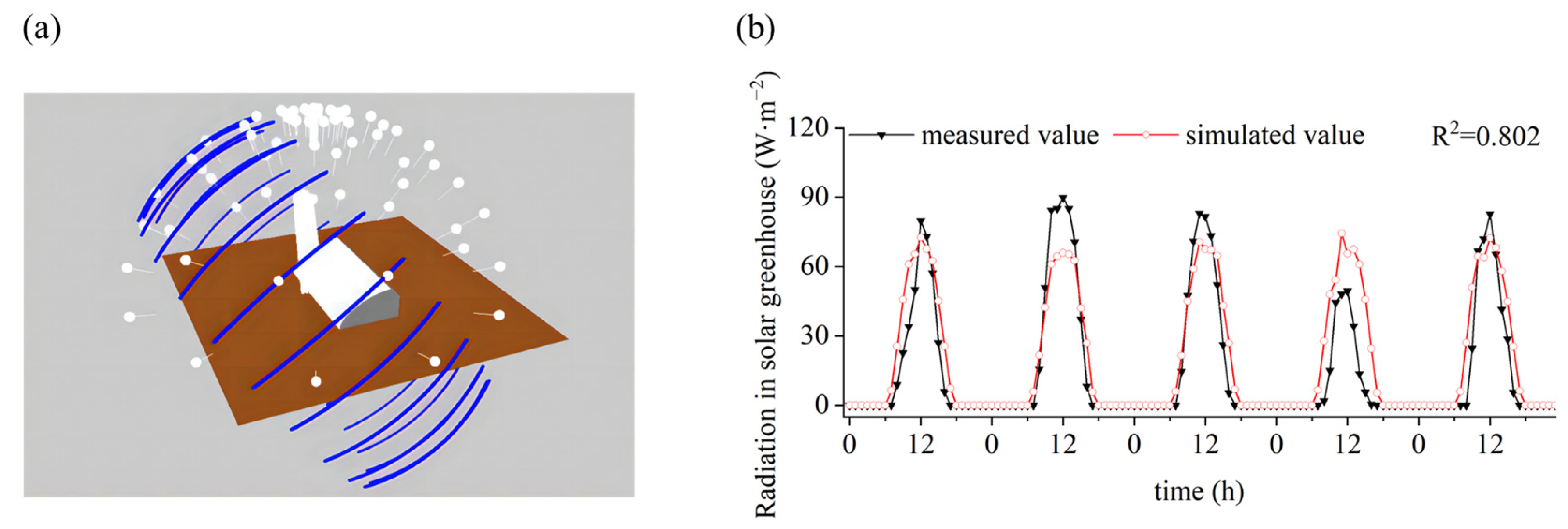
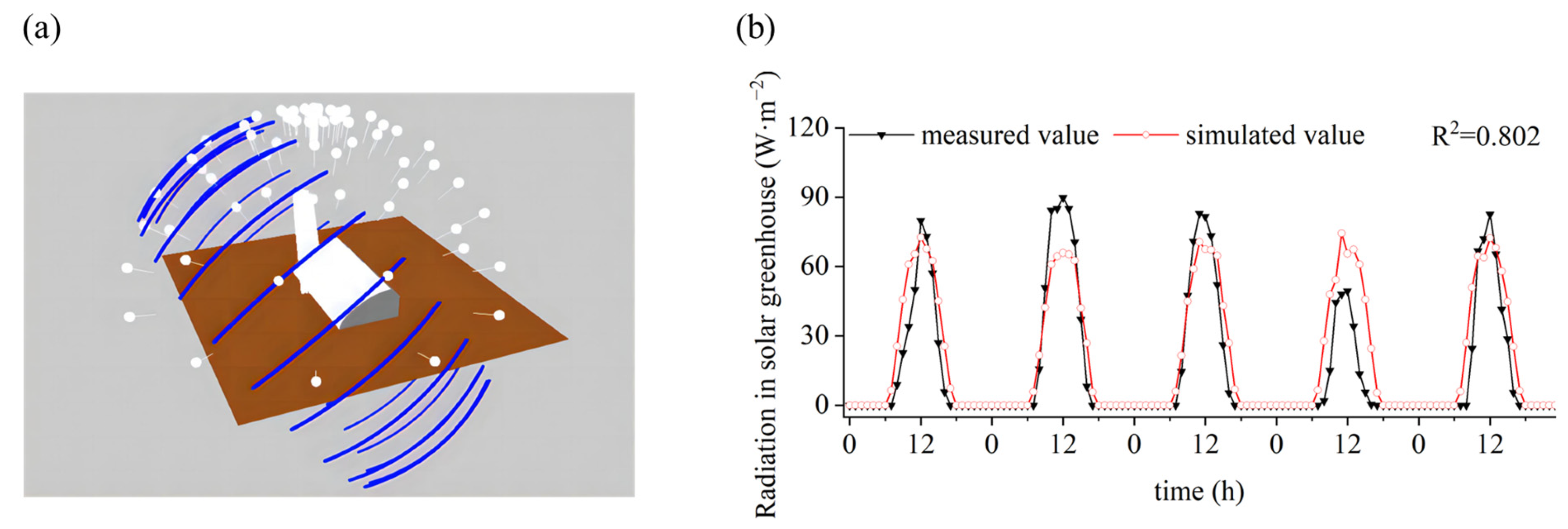
2.3.3. Sky and Sunlight Modules
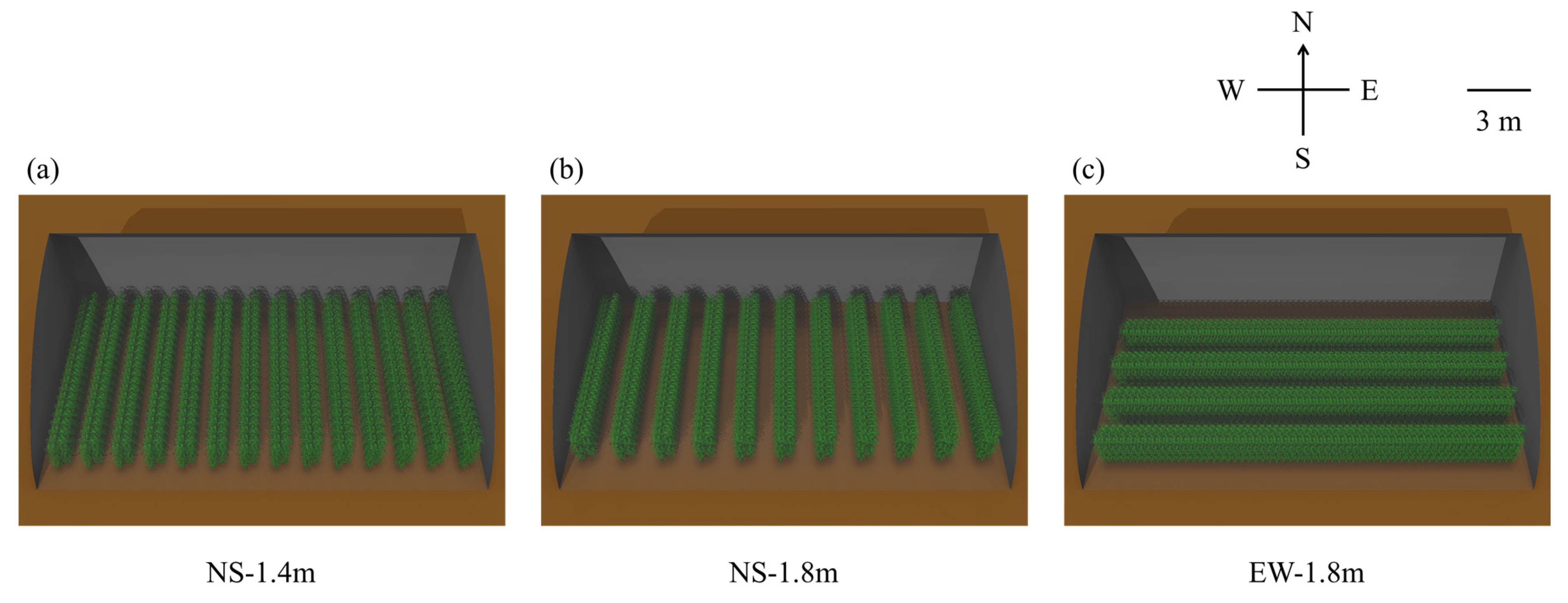
2.3.4. Model Scenarios
2.4. Data Analysis
3. Results
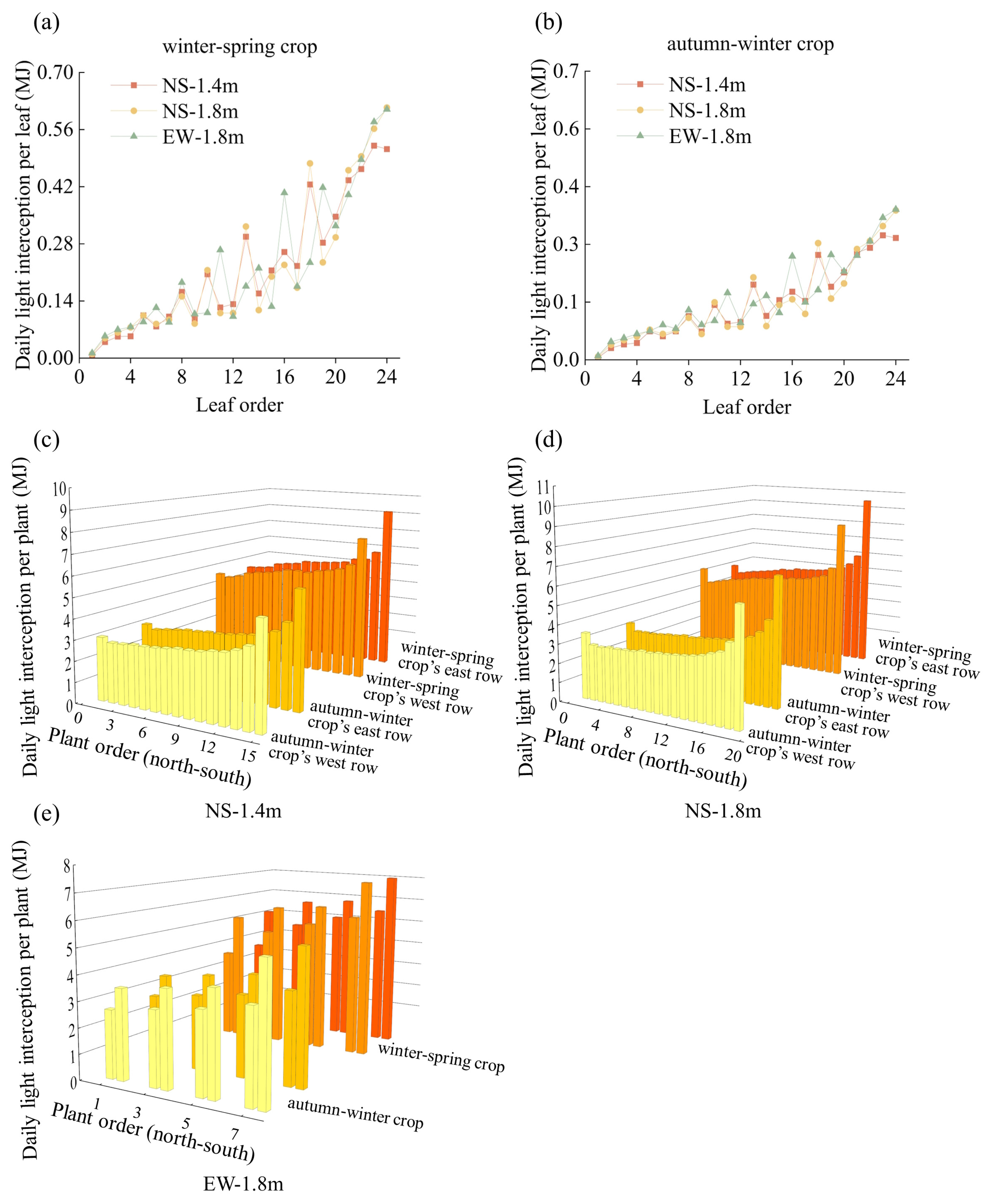
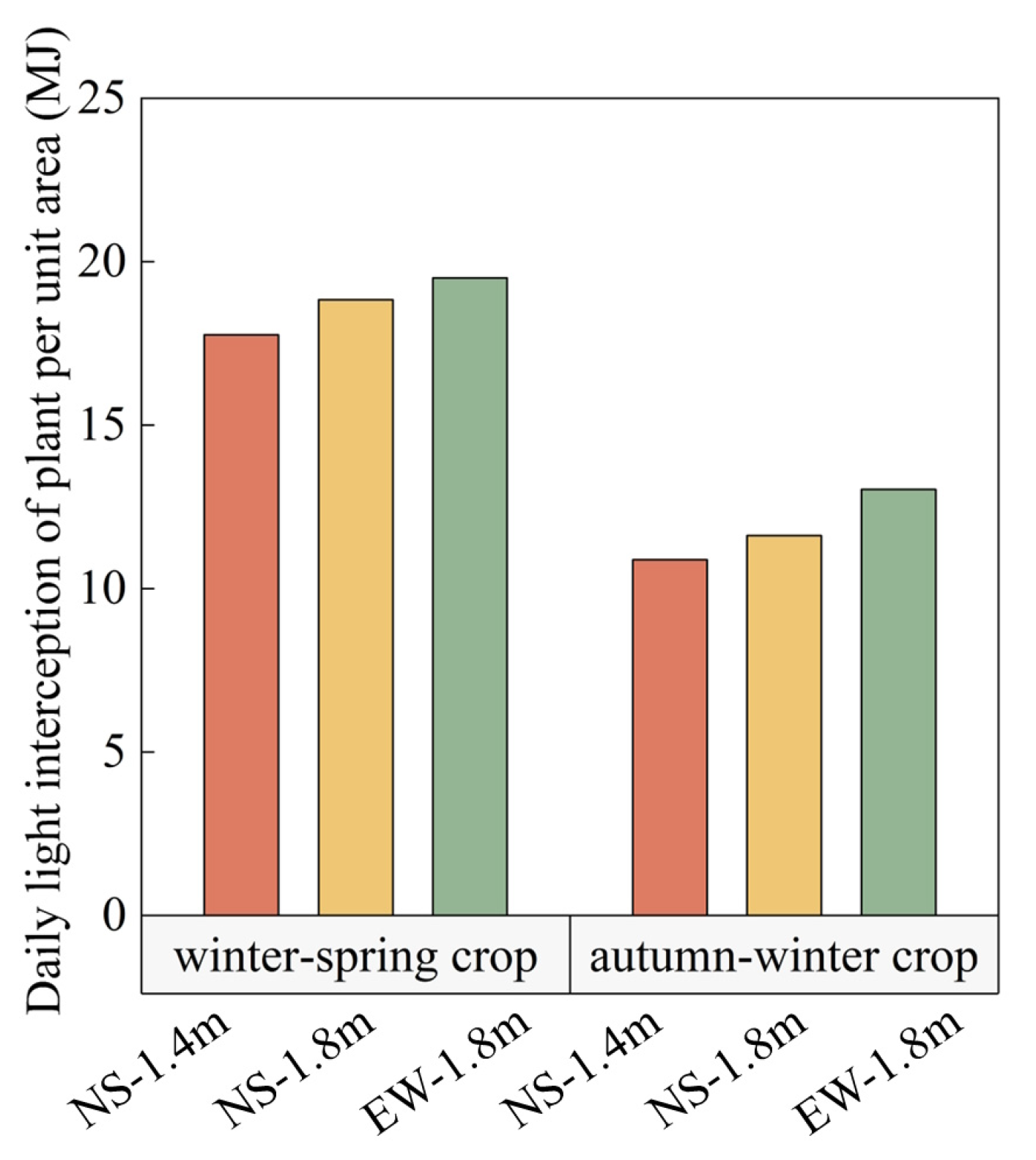
3.1. Light Interception in Tomato Canopies
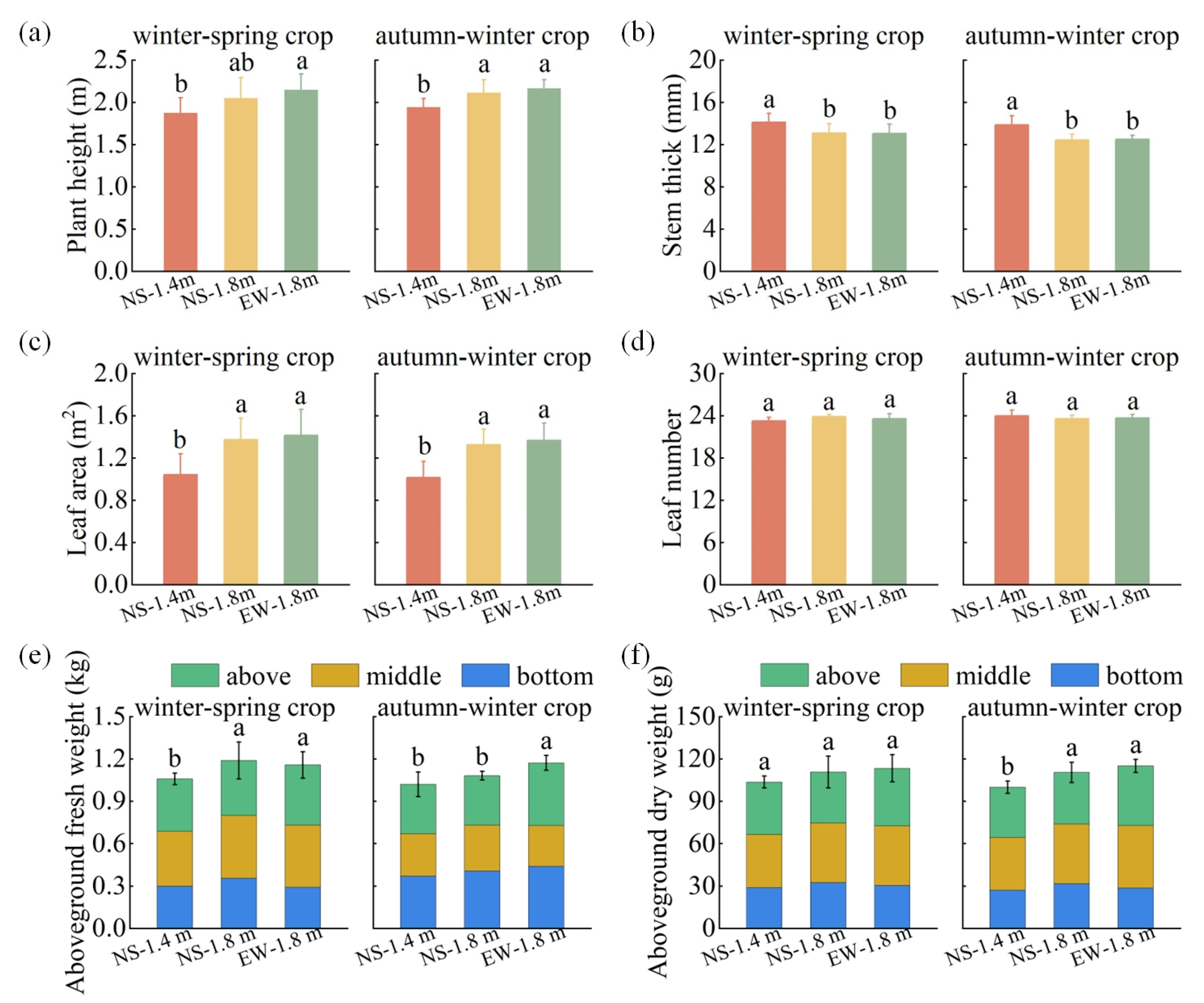
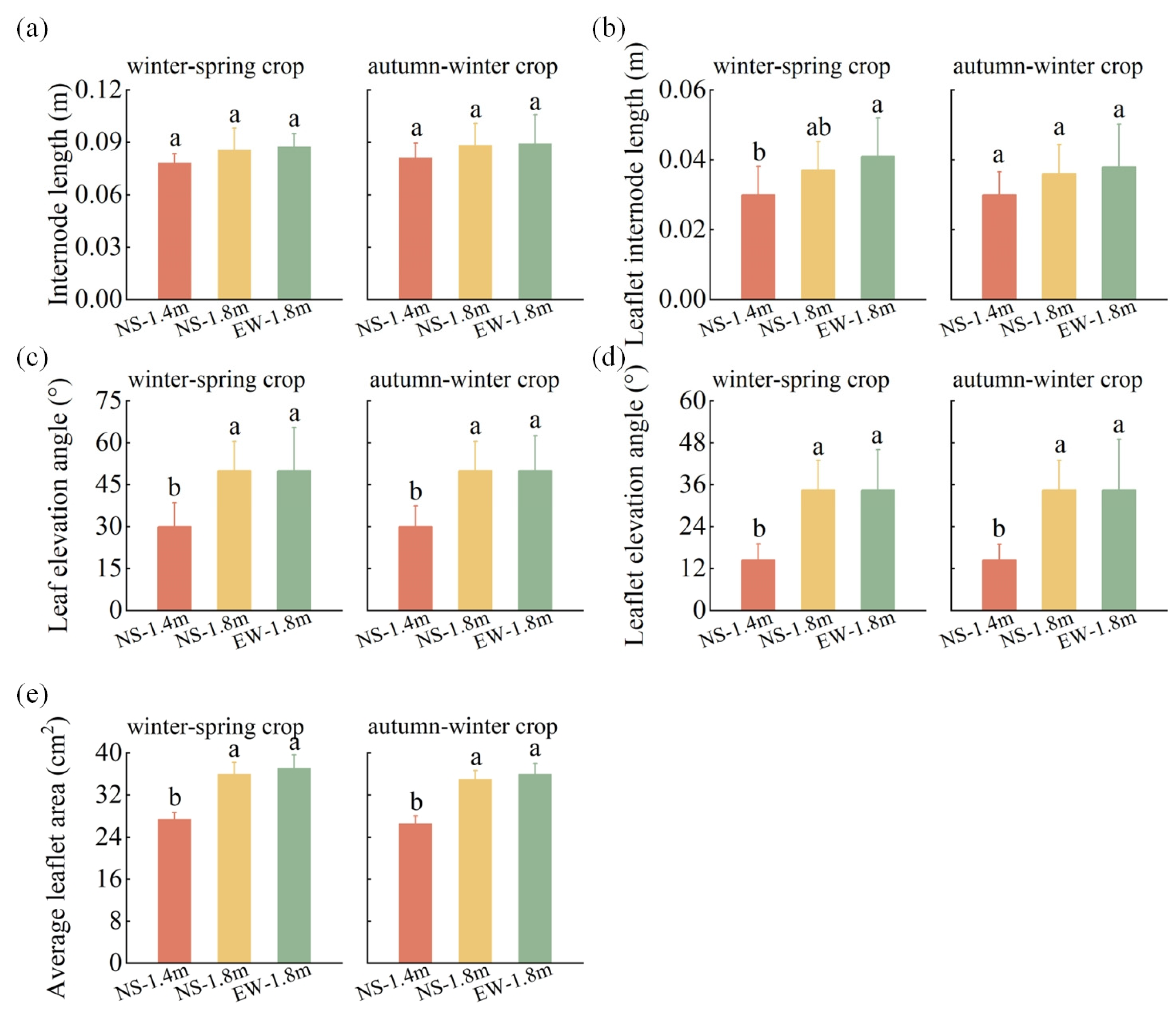
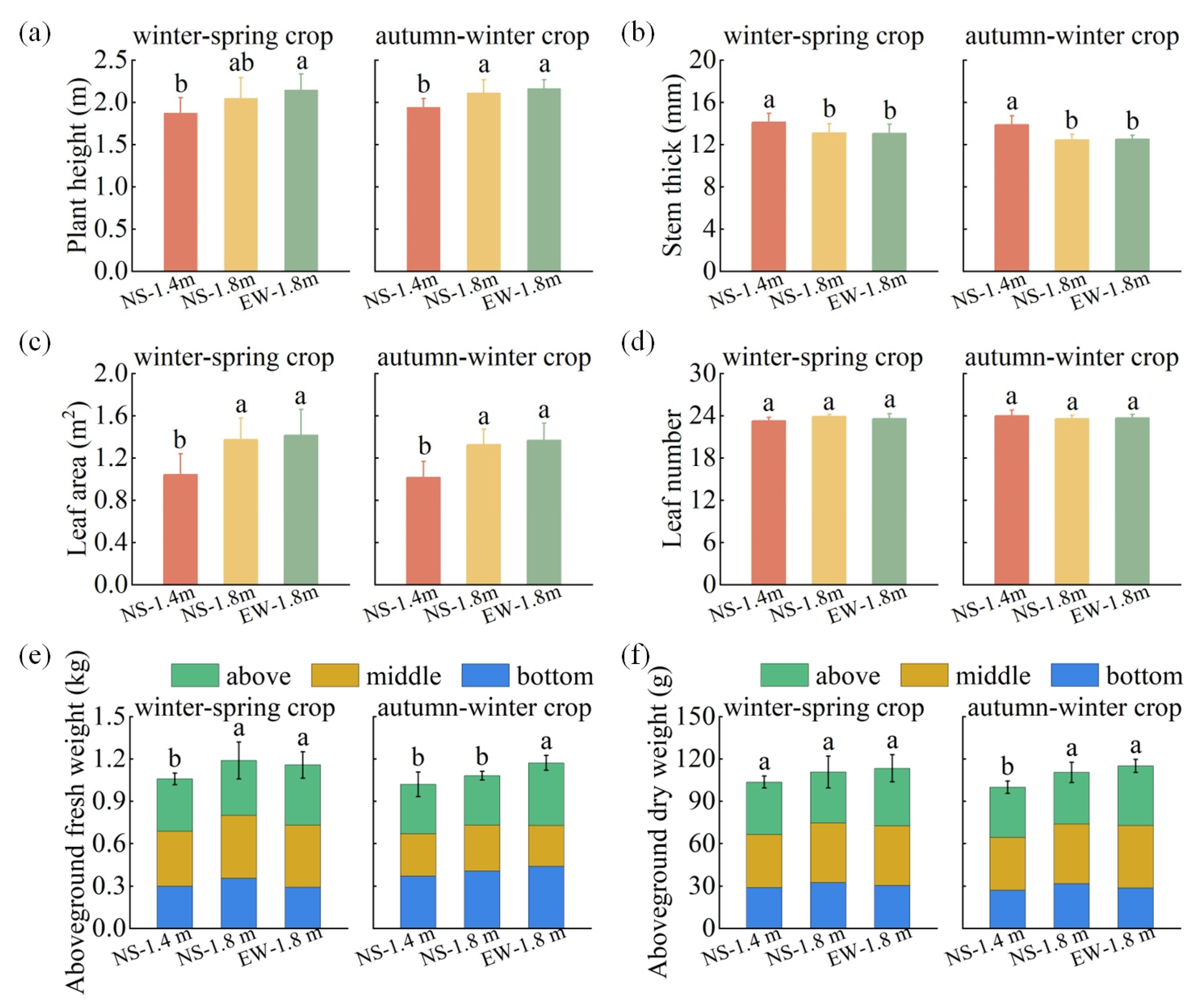
3.2. Tomato Growth Parameters
3.3. Gas Exchange Parameter
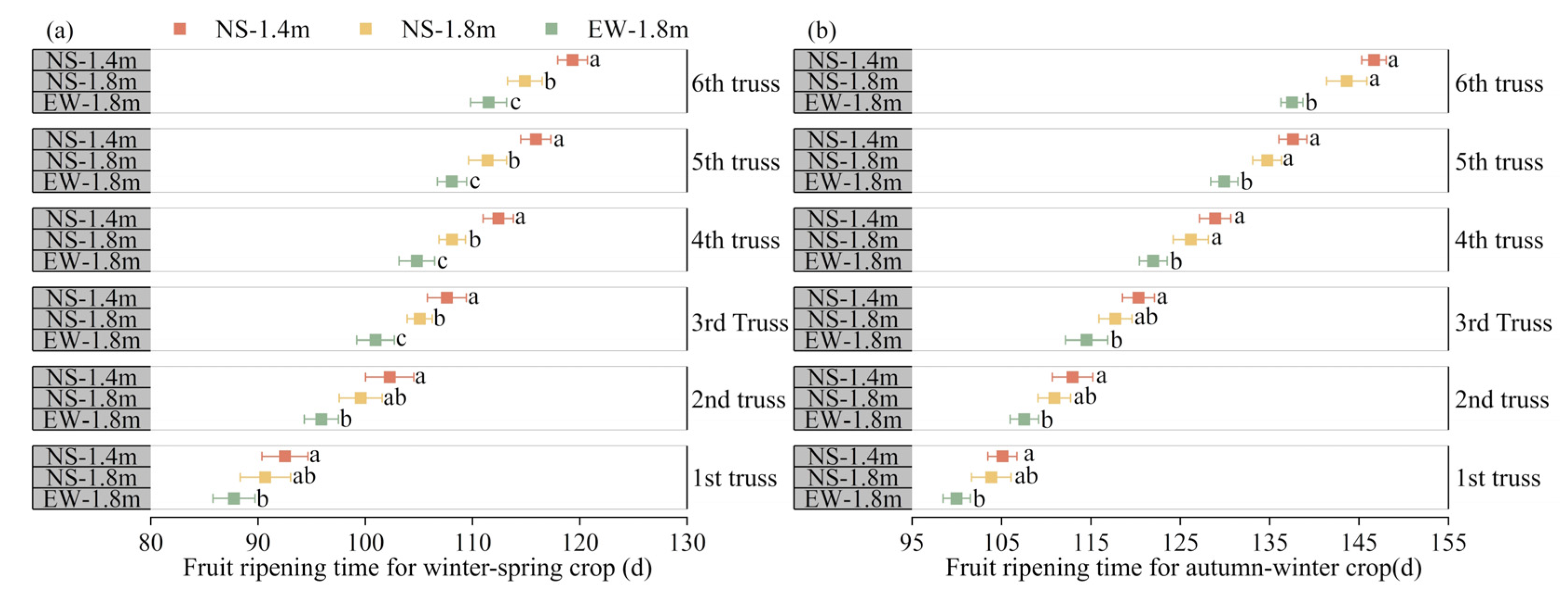
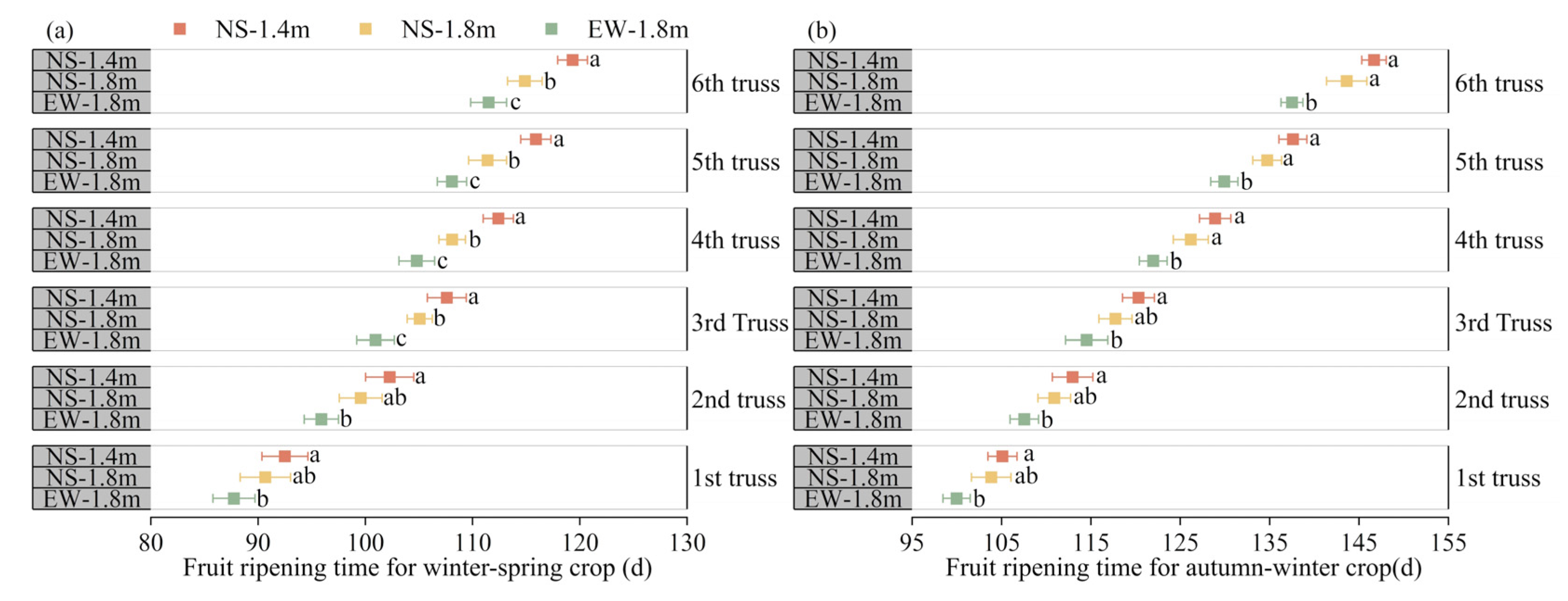
3.4. Fruit Ripening
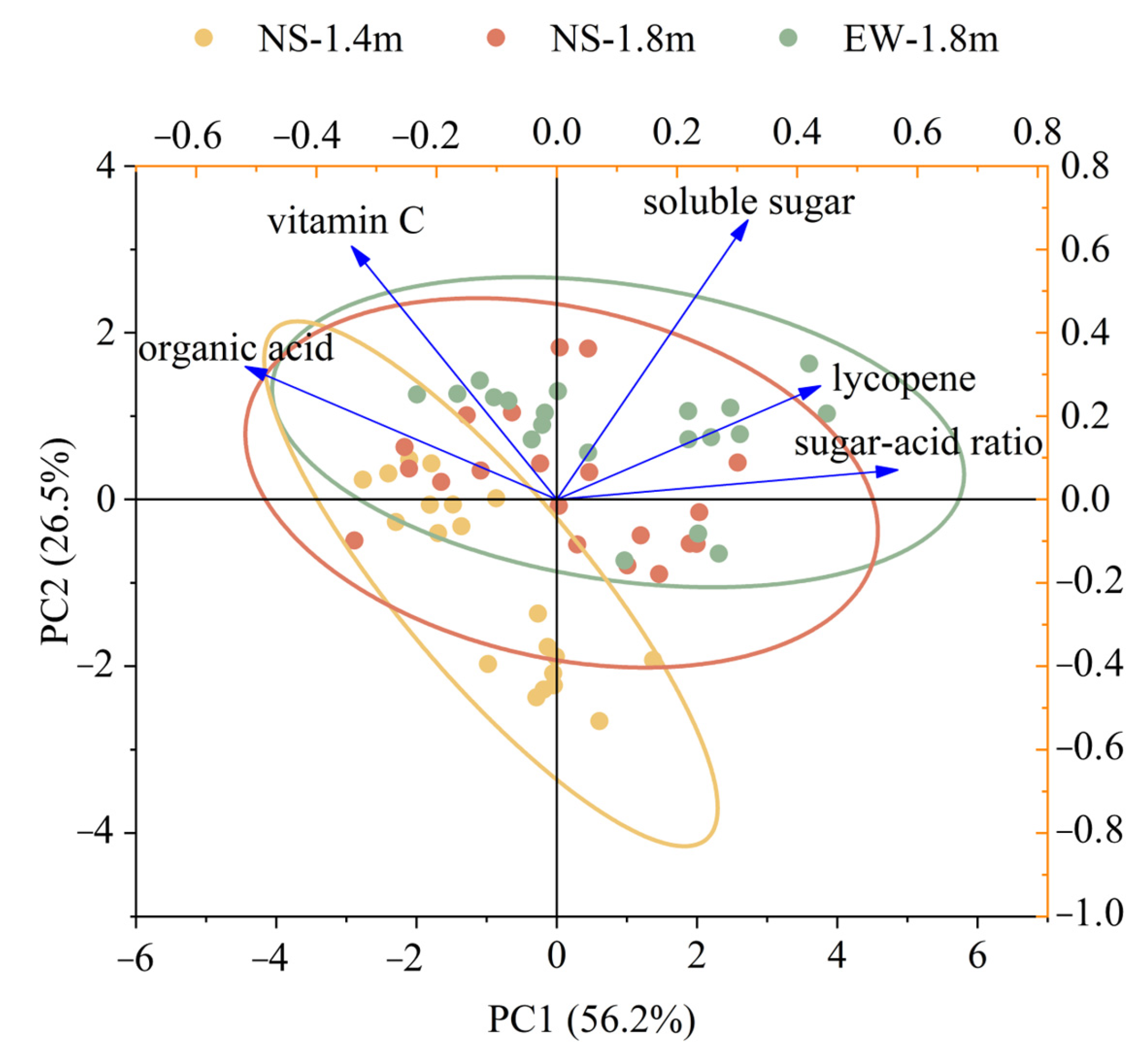
3.5. Fruit Quality and Yield
4. Discussion
4.1. Canopy Light Interception and Growth Characteristics of Tomatoes among Different Cultivation Patterns
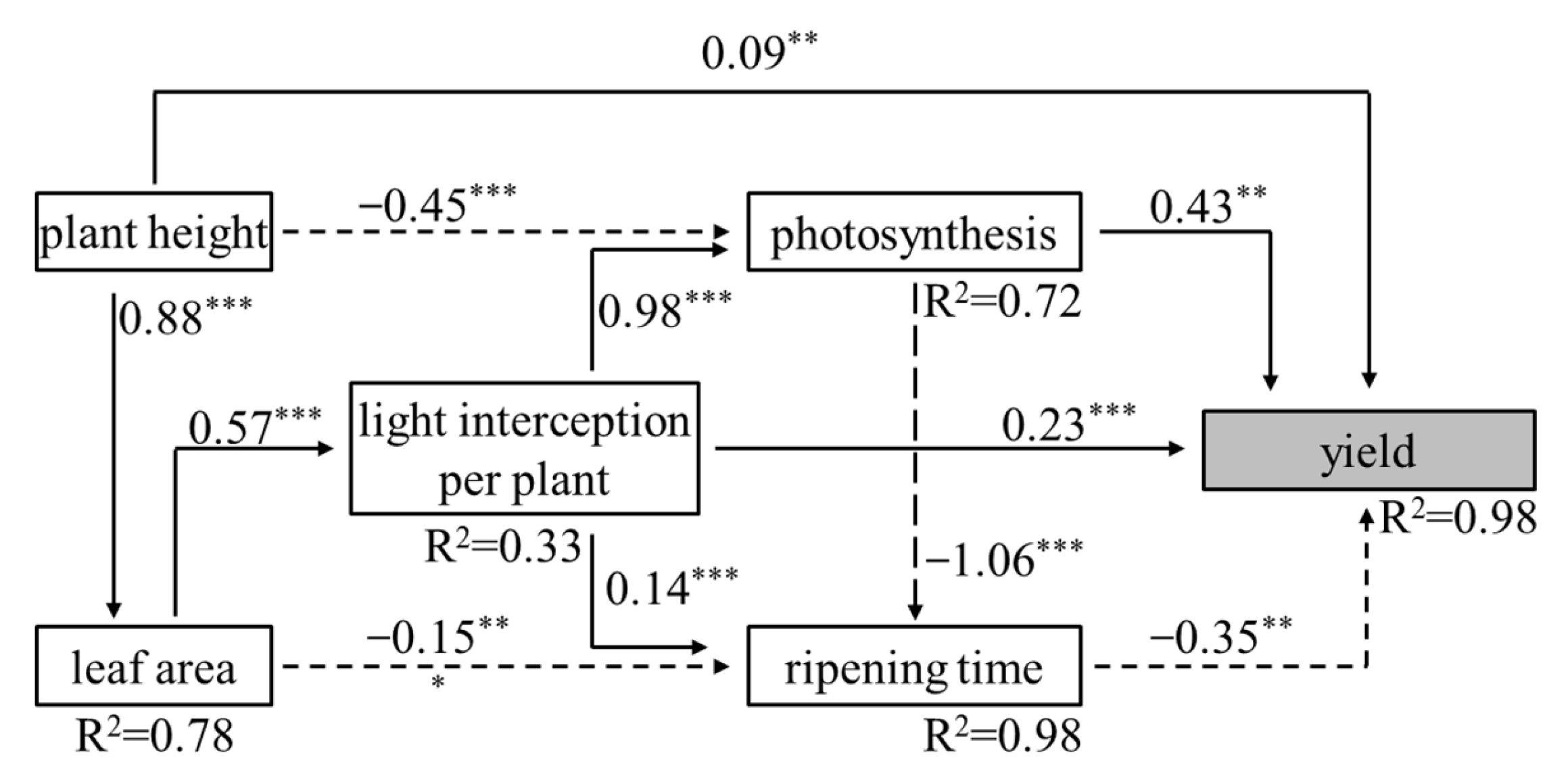
4.2. Fruit Ripening, Yield and Quality of Tomatoes among Different Cultivation Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kang, M.Z.; Fan, X.R.; Hua, J.; Wang, H.Y.; Wang, X.J.; Wang, F.Y. Managing traditional solar greenhouse with CPSS: A just-for-fit philosophy. IEEE Trans. Cybern. 2018, 48, 3371–3380. [Google Scholar] [CrossRef] [PubMed]
- Gürbüz Çolak, N.; Eken, N.T.; Ülger, M.; Frary, A.; Doğanlar, S. Mapping of quantitative trait loci for antioxidant molecules in tomato fruit: Carotenoids, vitamins C and E, glutathione and phenolic acids. Plant Sci. 2020, 292, 110393. [Google Scholar] [CrossRef] [PubMed]
- Sotelo-Cardona, P.; Lin, M.Y.; Srinivasan, R. Growing tomato under protected cultivation conditions: Overall effects on productivity, nutritional yield, and pest incidences. Crops 2021, 1, 97–110. [Google Scholar] [CrossRef]
- Song, W.T.; Li, C.X.; Sun, X.G.; Wang, P.Z.; Zhao, S.M. Effects of ridge direction on growth and yield of tomato in solar greenhouse with diffuse film. Trans. Chin. Soc. Agric. Eng. 2017, 33, 242–248. [Google Scholar]
- Zhang, X.D.; Lv, J.; Xie, J.M.; Yu, J.H.; Zhang, J.; Tang, C.N.; Li, J.; He, Z.X.; Wang, C. Solar radiation allocation and spatial distribution in Chinese solar greenhouses: Model development and application. Energies 2020, 13, 1108. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Y.C.; Zhu, D.L.; Zhang, X.M.; Lu, L.Q.; Gao, F.; Zheng, C.J. Dynamics of shaded areas in a typical-shaped solar greenhouse and their effects on tomato growth-A case study in winter. Sci. Hortic. 2023, 312, 111882. [Google Scholar] [CrossRef]
- Piao, L.; Zhang, S.Y.; Yan, J.Y.; Xiang, T.X.; Chen, Y.; Li, M.; Gu, W.R. Contribution of fertilizer, density and row spacing practices for maize yield and efficiency enhancement in northeast China. Plants 2022, 11, 2985. [Google Scholar] [CrossRef] [PubMed]
- Gomez-del-Campo, M.; Connor, D.J.; Trentacoste, E.R. Long-term effect of intra-row spacing on growth and productivity of super-high density hedgerow olive orchards (cv. Arbequina). Front. Plant Sci. 2017, 8, 1790. [Google Scholar] [CrossRef]
- Gómez-del-Campo, M.; Trentacoste, E.R.; Connor, D.J. Long-term effects of row spacing on radiation interception, fruit characteristics and production of hedgerow olive orchard (cv. Arbequina). Sci. Hortic. 2020, 272, 109583. [Google Scholar] [CrossRef]
- Brunel-Muguet, S.; Beauclair, P.; Bataillé, M.-P.; Avice, J.-C.; Trouverie, J.; Etienne, P.; Ourry, A. Light restriction delays leaf senescence in winter oilseed rape (Brassica napus L.). J. Plant Growth Regul. 2013, 32, 506–518. [Google Scholar] [CrossRef]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How does tomato quality (sugar, acid, and nutritional quality) vary with ripening stage, temperature, and irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Hernández, V.; Hellín, P.; Fenoll, J.; Flores, P. Interaction of nitrogen and shading on tomato yield and quality. Sci. Hortic. 2019, 255, 255–259. [Google Scholar] [CrossRef]
- Tang, J.N.; Chen, Q.F.; Yu, Z.G.; Fu, H.D.; Liu, C.C.; Zheng, S.R.; Song, W.L.; Sun, Z.P. Effect of east and west ridge cluster planting on the growth of autumn and winter tomato and the temperature and light environment in solar greenhouse. Chin. Agric. Sci. Bull. 2022, 38, 65–71. [Google Scholar] [CrossRef]
- Cui, D.Y.; Li, C.Q.; Sun, Y.X.; Wang, J.Q.; Zou, G.Y.; Yang, J.G. Effects of dwarf close planting on growth and yield of tomato under east-west cultivation in greenhouse. Acta Hortic. Sin. 2022, 49, 875–884. [Google Scholar]
- Buck-Sorlin, G.H.; Hemmerling, R.; Vos, J.; de Visser, P.H.B. Modelling of spatial light distribution in the greenhouse: Description of the model. In Proceedings of the 2009 Third International Symposium on Plant Growth Modeling, Simulation, Visualization and Applications, Beijing, China, 9–13 November 2009; pp. 79–86. [Google Scholar]
- Bongers, F.J.; Evers, J.B.; Anten, N.P.; Pierik, R. From shade avoidance responses to plant performance at vegetation level: Using virtual plant modelling as a tool. New Phytol. 2014, 204, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Vos, J.; Evers, J.B.; Buck-Sorlin, G.H.; Andrieu, B.; Chelle, M.; de Visser, P.H.B. Functional-structural plant modelling: A new versatile tool in crop science. J. Exp. Bot. 2009, 61, 2101–2115. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.L.; Zhang, L.Z.; Evers, J.B.; Henke, M.; van der Werf, W.; Liu, S.D.; Zhang, S.P.; Zhao, X.H.; Wang, B.M.; Li, Z.H. Identification of plant configurations maximizing radiation capture in relay strip cotton using a functional-structural plant model. Field Crops Res. 2016, 187, 1–11. [Google Scholar] [CrossRef]
- Barillot, R.; Chambon, C.; Fournier, C.; Combes, D.; Pradal, C.; Andrieu, B. Investigation of complex canopies with a functional-structural plant model as exemplified by leaf inclination effect on the functioning of pure and mixed stands of wheat during grain filling. Ann. Bot. 2018, 123, 727–742. [Google Scholar] [CrossRef]
- Li, S.W.; van der Werf, W.; Zhu, J.Q.; Guo, Y.; Li, B.G.; Ma, Y.T.; Evers, J.B. Estimating the contribution of plant traits to light partitioning in simultaneous maize/soybean intercropping. J. Exp. Bot. 2021, 72, 3630–3646. [Google Scholar] [CrossRef]
- Chang, T.G.; Zhao, H.l.; Wang, N.; Song, Q.F.; Xiao, Y.; Qu, M.n.; Zhu, X.G. A three-dimensional canopy photosynthesis model in rice with a complete description of the canopy architecture, leaf physiology, and mechanical properties. J. Exp. Bot. 2019, 70, 2479–2490. [Google Scholar] [CrossRef]
- Chen, T.W.; Henke, M.; de Visser, P.H.B.; Buck-Sorlin, G.; Wiechers, D.; Kahlen, K.; Stützel, H. What is the most prominent factor limiting photosynthesis in different layers of a greenhouse cucumber canopy? Ann. Bot. 2014, 114, 677–688. [Google Scholar] [CrossRef] [PubMed]
- De Visser, P.H.; Buck-Sorlin, G.H.; van Der Heijden, G.W. Optimizing illumination in the greenhouse using a 3D model of tomato and a ray tracer. Front. Plant Sci. 2014, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, M.; de Visser, P.H.B.; Heuvelink, E.; Marcelis, L.F.M. Row orientation affects the uniformity of light absorption, but hardly affects crop photosynthesis in hedgerow tomato crops. In Silico Plants 2021, 3, diab025. [Google Scholar] [CrossRef]
- Zhang, Y.; Henke, M.; Li, Y.M.; Xu, D.M.; Liu, A.H.; Liu, X.A.; Li, T.L. Analyzing the impact of greenhouse planting strategy and plant architecture on tomato plant physiology and estimated dry matter. Front. Plant Sci. 2022, 13, 828252. [Google Scholar] [CrossRef] [PubMed]
- Sarlikioti, V.; de Visser, P.H.B.; Marcelis, L.F.M. Exploring the spatial distribution of light interception and photosynthesis of canopies by means of a functional-structural plant model. Ann. Bot. 2011, 107, 875–883. [Google Scholar] [CrossRef]
- Qian, T.T.; Lu, S.L.; Zhao, C.J.; Guo, X.Y.; Wen, W.L.; Du, J.J. Heterogeneity analysis of cucumber canopy in the solar greenhouse. J. Integr. Agric. 2014, 13, 2645–2655. [Google Scholar] [CrossRef]
- Schneider, H.M. Characterization, costs, cues and future perspectives of phenotypic plasticity. Ann. Bot. 2022, 130, 131–148. [Google Scholar] [CrossRef]
- Chen, Z.Z.; Li, Y.M.; Luan, H.; Xu, L.H.; Hu, H.; Wei, M. Effects of ridge direction and plant and row spacing on growth and yield of tomato in solar greenhouse. Shandong Agric. Sci. 2022, 54, 63–67. [Google Scholar]
- Liu, L.; Wang, B.; Liu, D.; Zou, C.L.; Wu, P.; Wang, Z.Y.; Wang, Y.B.; Li, C.F. Transcriptomic and metabolomic analyses reveal mechanisms of adaptation to salinity in which carbon and nitrogen metabolism is altered in sugar beet roots. BMC Plant Biol. 2020, 20, 138. [Google Scholar] [CrossRef]
- Kaur, C.; Walia, S.; Nagal, S.; Walia, S.; Singh, J.; Singh, B.B.; Saha, S.; Singh, B.; Kalia, P.; Jaggi, S.; et al. Functional quality and antioxidant composition of selected tomato (Solanum lycopersicon L) cultivars grown in Northern India. LWT-Food Sci. Technol. 2013, 50, 139–145. [Google Scholar] [CrossRef]
- Qu, Z.M.; Qi, X.C.; Shi, R.G.; Zhao, Y.J.; Hu, Z.P.; Chen, Q.; Li, C.L. Reduced N fertilizer application with optimal blend of controlled-release urea and urea improves tomato yield and quality in greenhouse production system. J. Soil Sci. Plant Nutr. 2020, 20, 1741–1750. [Google Scholar] [CrossRef]
- Sun, Q.Q.; Zhang, N.; Wang, J.F.; Zhang, H.J.; Li, D.B.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S.X.; et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2014, 66, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.M.; Yang, Q.; Hou, M.M. The effects of saline water drip irrigation on tomato yield, quality, and blossom-end rot incidence-A 3a case study in the south of China. PLoS ONE 2015, 10, e0142204. [Google Scholar] [CrossRef] [PubMed]
- Kniemeyer, O. Design and Implementation of a Graph Grammar Based Language for Functional-Structural Plant Modelling. Doctoral Dissertation, BTU Cottbus-Senftenberg, Cottbus, Germany, 2008. [Google Scholar]
- Hemmerling, R.; Kniemeyer, O.; Lanwert, D.; Kurth, W.; Buck-Sorlin, G. The rule-based language XL and the modelling environment GroIMP illustrated with simulated tree competition. Funct. Plant Biol. 2008, 35, 739–750. [Google Scholar] [CrossRef]
- Park, S.J.; Jiang, K.; Tal, L.; Yichie, Y.; Gar, O.; Zamir, D.; Eshed, Y.; Lippman, Z.B. Optimization of crop productivity in tomato using induced mutations in the florigen pathway. Nat. Genet. 2014, 46, 1337–1342. [Google Scholar] [CrossRef] [PubMed]
- Vermeiren, J.; Villers, S.L.Y.; Wittemans, L.; Vanlommel, W.; van Roy, J.; Marien, H.; Coussement, J.R.; Steppe, K. Quantifying the importance of a realistic tomato (Solanum lycopersicum) leaflet shape for 3-D light modelling. Ann. Bot. 2019, 126, 661–670. [Google Scholar] [CrossRef]
- Zhang, Y.; Henke, M.; Li, Y.M.; Yue, X.; Xu, D.M.; Liu, X.A.; Li, T.L. High resolution 3D simulation of light climate and thermal performance of a solar greenhouse model under tomato canopy structure. Renew. Energy 2020, 160, 730–745. [Google Scholar] [CrossRef]
- Trentacoste, E.R.; Gómez-del-Campo, M.; Rapoport, H.F. Olive fruit growth, tissue development and composition as affected by irradiance received in different hedgerow positions and orientations. Sci. Hortic. 2016, 198, 284–293. [Google Scholar] [CrossRef]
- Chapepa, B.; Mudada, N.; Mapuranga, R. The impact of plant density and spatial arrangement on light interception on cotton crop and seed cotton yield: An overview. J. Cotton Res. 2020, 3, 18. [Google Scholar] [CrossRef]
- Feng, L.Y.; Raza, M.A.; Li, Z.C.; Chen, Y.K.; Khalid, M.H.B.; Du, J.B.; Liu, W.G.; Wu, X.L.; Song, C.; Yu, L.; et al. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef]
- Chang, J.Y.; Ma, X.L.; Wu, Y.L.; Li, J.M. Effects of row spacing and irrigation amount on canopy light interception and photosynthetic capacity, matter accumulation and fruit quality of tomato. Sci. Agric. Sin. 2023, 56, 2141–2157. [Google Scholar] [CrossRef]
- Liu, X.; Wang, W.X.; Lin, X.; Gu, S.B.; Wang, D. The effects of intraspecific competition and light transmission within the canopy on wheat yield in a wide-precision planting pattern. J. Integr. Agric. 2020, 19, 1577–1585. [Google Scholar] [CrossRef]
- Maldera, F.; Carone, V.; Castellarnau, I.I.; Vivaldi, G.A.; Camposeo, S. Available PAR, growth and yield of a super high-density almond orchard are influenced by different row orientations. Agronomy 2023, 13, 874. [Google Scholar] [CrossRef]
- Papadopoulos, A.P.; Pararajasingham, S. The influence of plant spacing on light interception and use in greenhouse tomato (Lycopersicon esculentum Mill.): A review. Sci. Hortic. 1997, 69, 1–29. [Google Scholar] [CrossRef]
- Wei, H.; Zhao, J.; Hu, J.T.; Jeong, B.R. Effect of supplementary light intensity on quality of grafted tomato seedlings and expression of two photosynthetic genes and proteins. Agronomy 2019, 9, 339. [Google Scholar] [CrossRef]
- Qian, T.; Dieleman, J.A.; Elings, A.; De Gelder, A.; Marcelis, L.F.M. Response of tomato crop growth and development to a vertical temperature gradient in a semi-closed greenhouse. J. Hortic. Sci. Biotechnol. 2015, 90, 578–584. [Google Scholar] [CrossRef]
- Paucek, I.; Pennisi, G.; Pistillo, A.; Appolloni, E.; Crepaldi, A.; Calegari, B.; Spinelli, F.; Cellini, A.; Gabarrell, X.; Orsini, F.; et al. Supplementary LED interlighting improves yield and precocity of greenhouse tomatoes in the Mediterranean. Agronomy 2020, 10, 1002. [Google Scholar] [CrossRef]
- Trentacoste, E.R.; Connor, D.J.; Gómez-del-Campo, M. Effect of row spacing on vegetative structure, fruit characteristics and oil productivity of N-S and E-W oriented olive hedgerows. Sci. Hortic. 2015, 193, 240–248. [Google Scholar] [CrossRef]
- Gil, H.J.; Kim, Y.X.; Sung, J.; Jung, E.S.; Singh, D.; Lee, Y.; Lee, D.; Lee, C.H.; Lee, S. Metabolomic insights of the tomato fruits (Solanum lycopersicum L.) cultivated under different supplemental LED lighting and mineral nutrient conditions. Hortic. Environ. Biotechnol. 2020, 61, 415–427. [Google Scholar] [CrossRef]
- Toor, R.K.; Savage, G.P.; Lister, C.E. Seasonal variations in the antioxidant composition of greenhouse grown tomatoes. J. Food Compos. Anal. 2006, 19, 1–10. [Google Scholar] [CrossRef]
- Appolloni, E.; Orsini, F.; Pennisi, G.; Gabarrell Durany, X.; Paucek, I.; Gianquinto, G. Supplemental LED lighting effectively enhances the yield and quality of greenhouse truss tomato production: Results of a meta-analysis. Front. Plant Sci. 2021, 12, 596927. [Google Scholar] [CrossRef]
- Appolloni, E.; Paucek, I.; Pennisi, G.; Manfrini, L.; Gabarrell, X.; Gianquinto, G.; Orsini, F. Winter greenhouse tomato cultivation: Matching leaf pruning and supplementary lighting for improved yield and precocity. Agronomy 2023, 13, 671. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alkali-Hydrolyzable Nitrogen (mg·kg−1) | Available Phosphorus (mg·kg−1) | Available Potassium (mg·kg−1) | Organic Matter (g·kg−1) | Bulk Density (g·cm−1) |
|---|---|---|---|---|
| 175.8 | 67.5 | 246.2 | 20.1 | 1.14 |
| Parameters | Schematic | Value | Unit | |||||
|---|---|---|---|---|---|---|---|---|
| Winter–Spring Crop | Autumn–Winter Crop | |||||||
| NS-1.4m | NS-1.8m | EW-1.8m | NS-1.4m | NS-1.8m | EW-1.8m | |||
| Leaf phyllotactic angle | 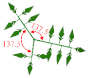 | 137.5 | 137.5 | 137.5 | 137.5 | 137.5 | 137.5 | ° |
| Internode length |  | 0.0784 | 0.0858 | 0.0875 | 0.0812 | 0.0884 | 0.0900 | m |
| Leaf elevation angle | 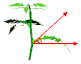 | 30 | 50 | 50 | 30 | 50 | 50 | ° |
| Leaflet internode length | 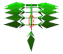 | 0.030 | 0.037 | 0.041 | 0.030 | 0.036 | 0.038 | m |
| Leaflet elevation angle | 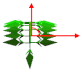 | 15 | 35 | 35 | 15 | 35 | 35 | ° |
| Average leaflet area |  | 0.00274 | 0.00360 | 0.00371 | 0.00266 | 0.00350 | 0.00360 | m2 |
| Treatment | Net Photosynthetic Rate (μmol·m−2·s−1) | Intercellular CO2 Concentration (μmol·mol−1) | Stomatal Conductance (mmol·m−2·s−1) | Transpiration Rate (mmol·m−2·s−1) | |
|---|---|---|---|---|---|
| Winter–spring crop | NS-1.4m | 25.68 ± 0.41 b | 255.35 ± 5.09 b | 389.85 ± 6.38 ab | 6.89 ± 0.51 b |
| NS-1.8m | 26.04 ± 0.78 b | 255.44 ± 7.14 b | 387.01 ± 8.12 b | 7.15 ± 0.58 b | |
| EW-1.8m | 27.01 ± 0.39 a | 267.58 ± 5.82 a | 394.76 ± 4.37 a | 7.69 ± 0.32 a | |
| Autumn–winter crop | NS-1.4m | 15.93 ± 0.55 b | 253.41 ± 5.82 b | 389.68 ± 9.38 a | 6.79 ± 0.45 a |
| NS-1.8m | 16.71 ± 0.46 a | 260.59 ± 2.34 a | 396.17 ± 9.77 a | 7.13 ± 0.40 a | |
| EW-1.8m | 16.97 ± 0.60 a | 265.59 ± 7.23 a | 399.18 ± 8.33 a | 7.16 ± 0.48 a | |
| Treatment | Soluble Sugar (%) | Organic Acid (%) | Sugar–Acid Ratio | Lycopene (μg·g−1) | Vitamin C (mg·100g−1) | |
|---|---|---|---|---|---|---|
| Winter–spring crop | NS-1.4m | 3.57 ± 0.14 c | 0.50 ± 0.06 a | 7.19 ± 1.05 b | 93.44 ± 4.55 c | 18.54 ± 1.85 b |
| NS-1.8m | 4.35 ± 0.26 b | 0.49 ± 0.08 a | 9.02 ± 1.52 a | 101.55 ± 6.25 b | 25.74 ± 2.25 a | |
| EW-1.8m | 4.84 ± 0.46 a | 0.47 ± 0.06 a | 10.29 ± 1.63 a | 107.95 ± 2.75 a | 24.65 ± 3.24 a | |
| Autumn–winter crop | NS-1.4m | 4.05 ± 0.14 b | 0.69 ± 0.05 b | 5.86 ± 0.53 b | 85.55 ± 6.74 b | 41.05 ± 6.34 b |
| NS-1.8m | 4.35 ± 0.26 a | 0.66 ± 0.06 ab | 6.63 ± 0.97 ab | 91.35 ± 13.8 ab | 44.54 ± 2.85 b | |
| EW-1.8m | 4.29 ± 0.10 a | 0.61 ± 0.08 a | 7.09 ± 1.02 a | 100.14 ± 5.65 a | 51.65 ± 4.15 a | |
| Treatment | Number of Fruits per Plant | Individual Fruit Weight (g) | Yield per Plant (kg) | Yield per Unit Area (t∙ha−1) | |
|---|---|---|---|---|---|
| Winter–spring crop | NS-1.4m | 22.1 ± 0.87 a | 217.42 ± 9.84 b | 4.80 ± 0.21 b | 172.86 ± 7.85 b |
| NS-1.8m | 22.0 ± 0.47 a | 226.86 ± 12.74 ab | 4.98 ± 0.27 ab | 179.63 ± 9.82 ab | |
| EW-1.8m | 22.2 ± 0.63 a | 229.72 ± 8.61 a | 5.09 ± 0.20 a | 183.55 ± 7.52 a | |
| Autumn–winter crop | NS-1.4m | 18.2 ± 0.78 a | 189.30 ± 9.33 b | 3.44 ± 0.25 b | 124.10 ± 9.24 b |
| NS-1.8m | 18.4 ± 1.26 a | 195.09 ± 12.67 ab | 3.59 ± 0.35 ab | 129.27 ± 12.75 ab | |
| EW-1.8m | 18.5 ± 0.52 a | 204.49 ± 10.26 a | 3.78 ± 0.24 a | 136.24 ± 8.70 a | |
| Indicator | Direct Effect | Indirect Effect | Total Effect |
|---|---|---|---|
| Plant height | 0.09 | 0.18 | 0.27 |
| Leaf area | 0.00 | 0.61 | 0.61 |
| Light interception per plant | 0.23 | 0.75 | 0.98 |
| Photosynthesis | 0.43 | 0.38 | 0.81 |
| Ripening time | −0.35 | 0.00 | −0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Henke, M.; Zhang, D.; Wang, C.; Wei, M. Optimized Tomato Production in Chinese Solar Greenhouses: The Impact of an East–West Orientation and Wide Row Spacing. Agronomy 2024, 14, 314. https://doi.org/10.3390/agronomy14020314
Li Y, Henke M, Zhang D, Wang C, Wei M. Optimized Tomato Production in Chinese Solar Greenhouses: The Impact of an East–West Orientation and Wide Row Spacing. Agronomy. 2024; 14(2):314. https://doi.org/10.3390/agronomy14020314
Chicago/Turabian StyleLi, Yiman, Michael Henke, Dalong Zhang, Chuanqing Wang, and Min Wei. 2024. "Optimized Tomato Production in Chinese Solar Greenhouses: The Impact of an East–West Orientation and Wide Row Spacing" Agronomy 14, no. 2: 314. https://doi.org/10.3390/agronomy14020314
APA StyleLi, Y., Henke, M., Zhang, D., Wang, C., & Wei, M. (2024). Optimized Tomato Production in Chinese Solar Greenhouses: The Impact of an East–West Orientation and Wide Row Spacing. Agronomy, 14(2), 314. https://doi.org/10.3390/agronomy14020314