
Antioxidant and Ultrastructural Alterations in Wheat During Drought-Induced Leaf Senescence

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation of Plant Material
2.2. Chlorophyll Content
2.3. Membrane Stability Index (MSI)
2.4. Lipid Peroxidation Assay
2.5. Total Protein Content
2.6. Enzyme Activity Measurement
2.6.1. Preparation of Enzyme Extract
2.6.2. Ascorbate Peroxidase (APX, EC 1.11.1.11)
2.6.3. Guaiacol Peroxidase (GPX, EC 1.11.1.7)
2.6.4. Benzidine Peroxidase (BPX, EC 1.11.1.7)
2.6.5. Catalase (CAT, E.C. 1.11.1.6)
2.6.6. Superoxide Dismutase (SOD, EC 1.15.1.1)
2.7. Gene Expression Analysis
2.7.1. RNA Extraction and cDNA Synthesis
2.7.2. Quantitative Real-Time PCR
2.8. Analysis of Leaf Mesophyll Cells by Light and Transmission Electron Microscopy (TEM)
2.9. Statistical Analysis
3. Results
3.1. Analysis of Variance
3.2. Chlorophyll Content
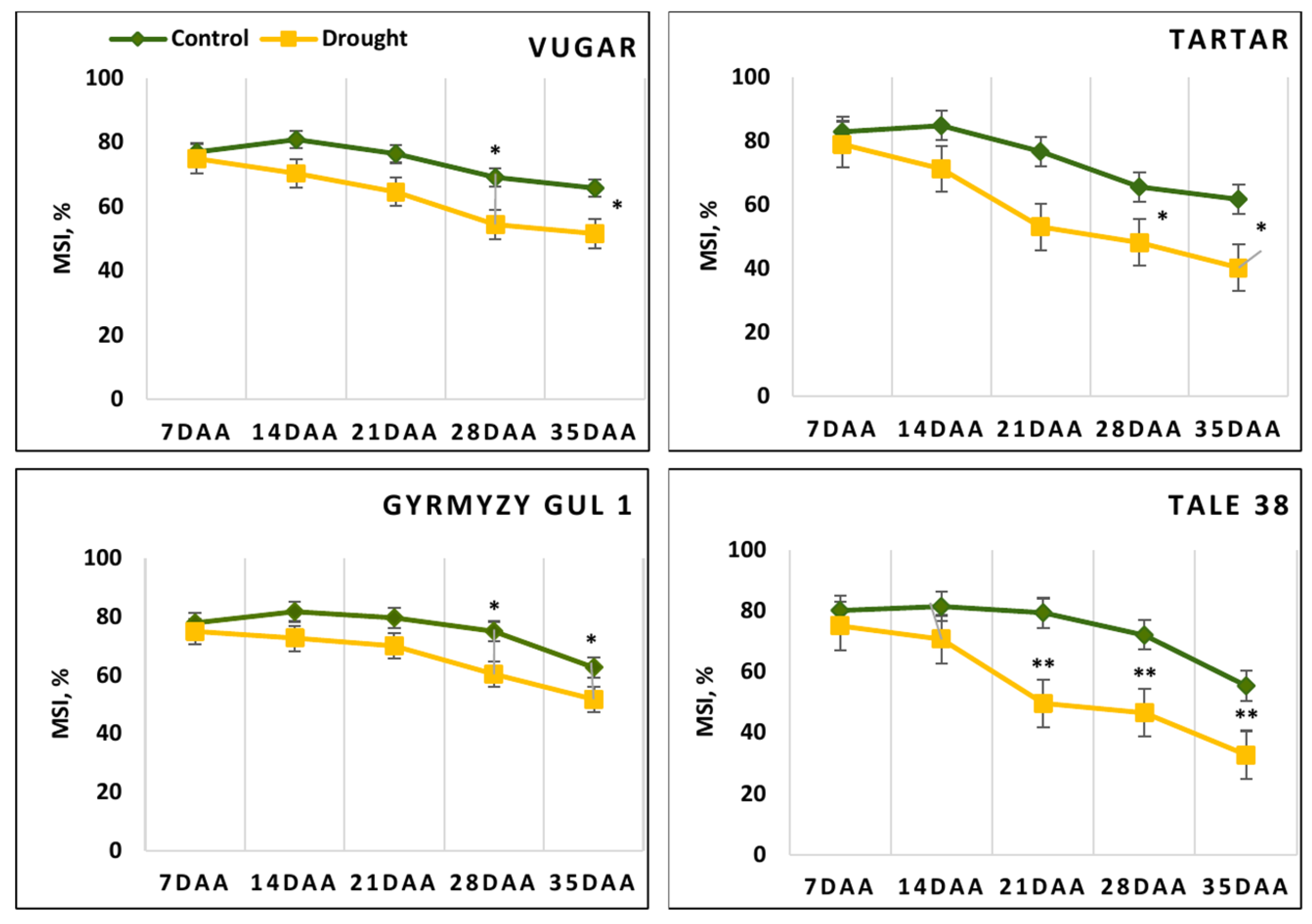
3.3. Membrane Stability
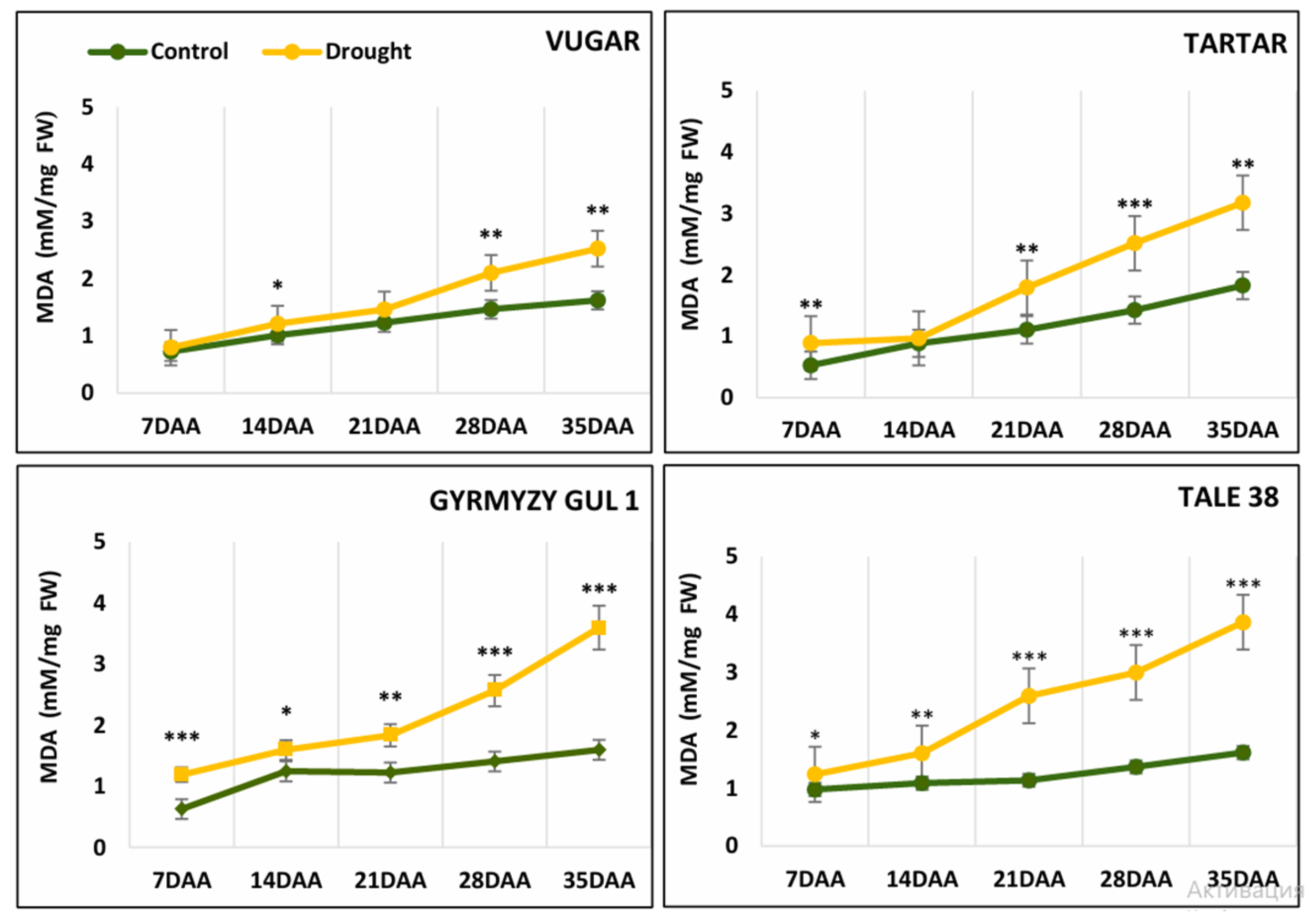
3.4. Malondialdehyde Content
3.5. Soluble Protein Content
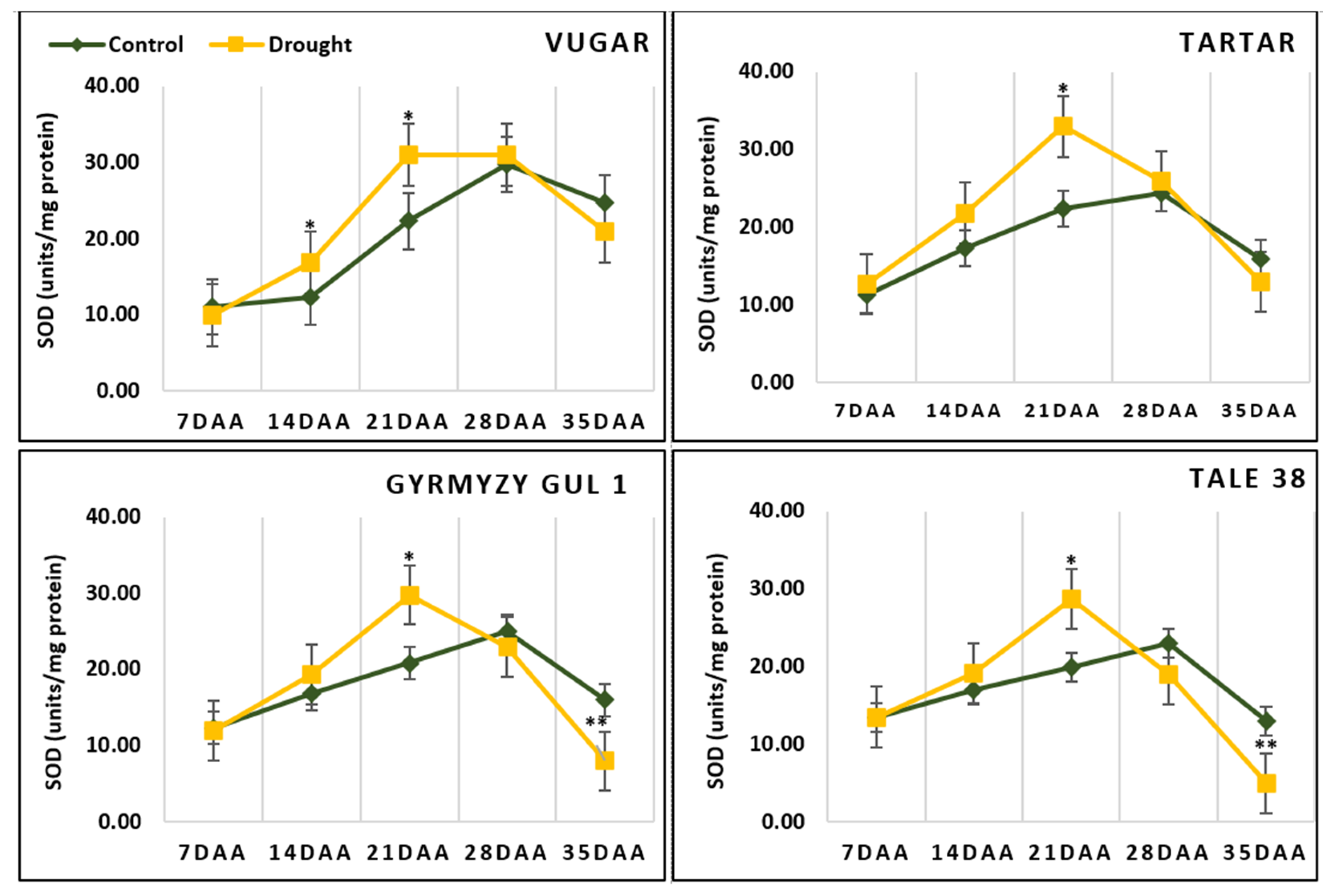
3.6. The Activity of SOD
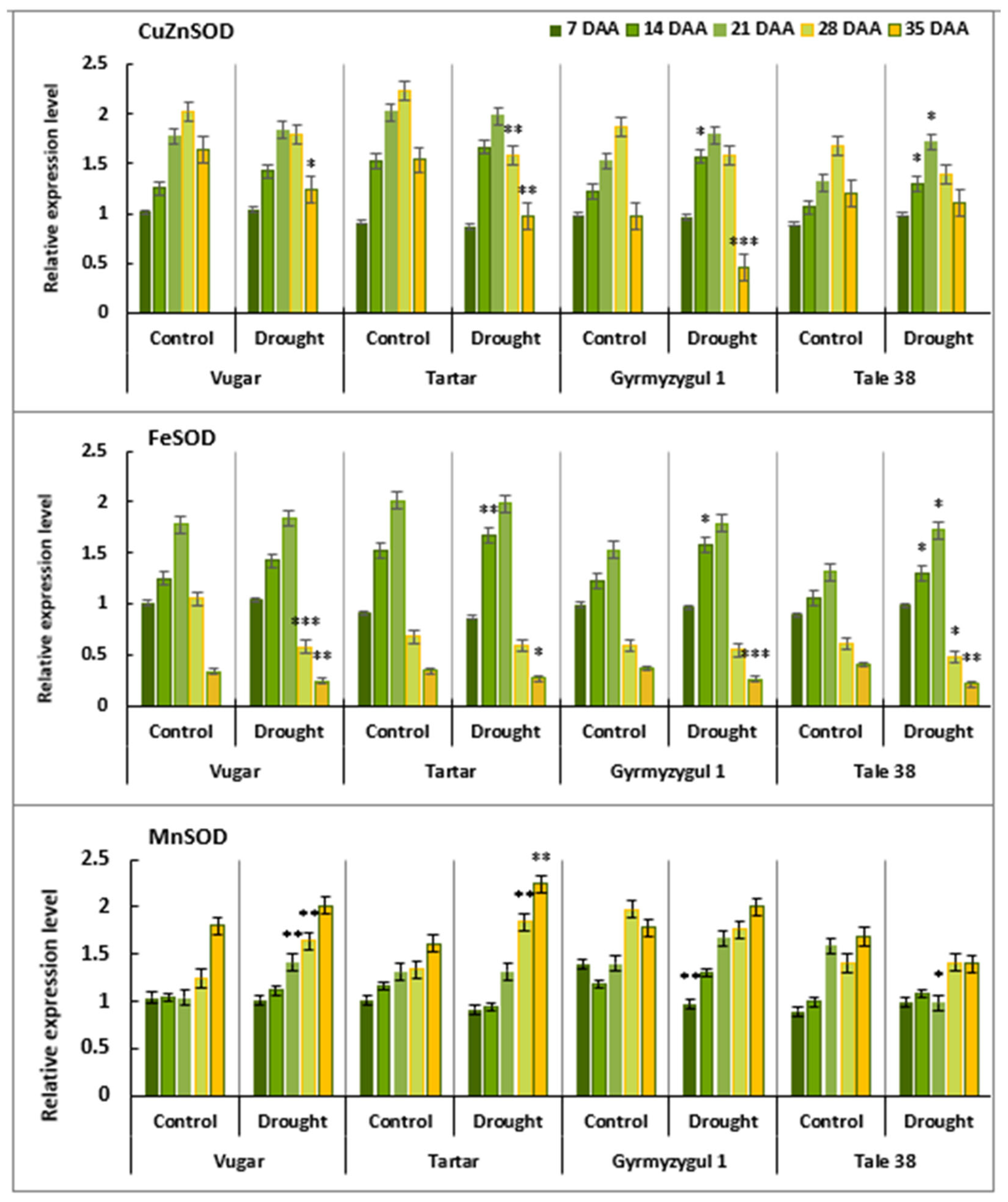
3.7. Gene Expression Analysis of SOD Isoforms
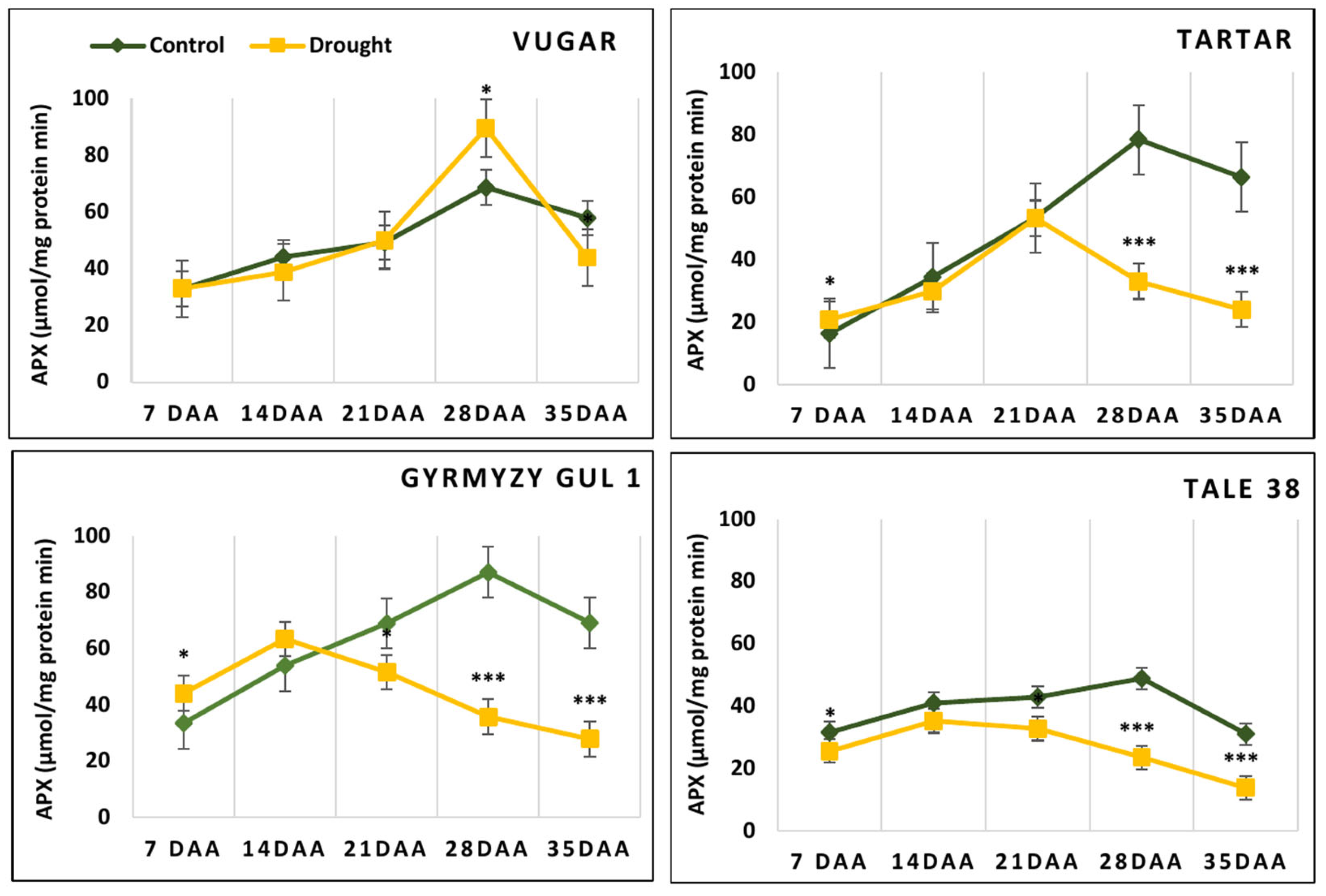
3.8. The Activity of Peroxidases
3.9. The Activity of CAT
3.10. Light Microscopy
3.11. Transmission Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Cheng, S.; Feng, C.; Wingen, L.U.; Cheng, H.; Riche, A.B.; Jiang, M.; Leverington-Waite, M.; Huang, Z.; Collier, S.; Orford, S.; et al. Harnessing landrace diversity empowers wheat breeding. Nature 2024, 632, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhao, H.; Wang, H.; He, Y. Research progress on the relationship between leaf senescence and quality, yield and stress resistance in horticultural plants. Front. Plant Sci. 2022, 13, 1044500. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.A.; Orford, S.; Lage, J.; Griffiths, S. Capturing and selecting senescence variation in wheat. Front. Plant Sci. 2021, 12, 638738. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Yang, M.; Rasheed, A.; Tian, X.; Reynolds, M.; Xia, X.; Xiao, Y.; He, Z. Quantifying senescence in bread wheat using multispectral imaging from an unmanned aerial vehicle and QTL mapping. Plant Physiol. 2021, 187, 2623–2636. [Google Scholar] [CrossRef]
- Li, C.; Wu, X.; Wang, P.; Wang, H.; Wang, L.; Sun, F.; Jing, H.C. Genome-wide association study of image-based trait reveals the genetic architecture of dark-induced leaf senescence in rice. J. Exp. Bot. 2024, erae391. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, H.J.; Lim, P.O.; Nam, H.G. Leaf senescence: Systems and dynamics aspects. Annu. Rev. Plant Biol. 2019, 70, 347–376. [Google Scholar] [CrossRef]
- Aloryi, K.D.; Jing, H.C.; Dijkwel, P.P. Comparison of leaf senescence regulation between distantly related plant species uncovers knowledge gaps and opportunities for plant improvement strategies. Environ. Exp. Bot. 2023, 214, 105474. [Google Scholar] [CrossRef]
- Guo, Y.; Ren, G.; Zhang, K.; Li, Z.; Miao, Y.; Guo, H. Leaf senescence: Progression, regulation, and application. Mol. Hortic. 2021, 1, 5. [Google Scholar] [CrossRef]
- Tan, S.; Sha, Y.; Sun, L.; Li, Z. Abiotic stress-induced leaf senescence: Regulatory mechanisms and application. Int. J. Mol. Sci. 2023, 24, 11996. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, Y.; Tan, S.; Yang, Q.; Wang, H.-L.; Xia, X.; Luo, J.; Guo, H.; Zhang, Z.; Li, Z. LSD 4.0: An improved database for comparative studies of leaf senescence. Mol. Hortic. 2022, 2, 24. [Google Scholar] [CrossRef]
- Sultana, N.; Islam, S.; Juhász, A.; Ma, W. Wheat leaf senescence and its regulatory gene network. Crop J. 2021, 9, 703–717. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Hajibarat, Z.; Saidi, A. Senescence-associated proteins and nitrogen remobilization in grain filling under drought stress condition. J. Genet. Eng. Biotechnol. 2022, 20, 101. [Google Scholar] [CrossRef]
- Joshi, S.; Choukimath, A.; Isenegger, D.; Panozzo, J.; Spangenberg, G.; Kant, S. Improved wheat growth and yield by delayed leaf senescence using developmentally regulated expression of a cytokinin biosynthesis gene. Front. Plant Sci. 2019, 10, 1285. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant senescence and crop productivity. Plant Mol. Biol. 2013, 81, 603–622. [Google Scholar] [CrossRef]
- Camargo Rodriguez, A.V. Integrative modelling of gene expression and digital phenotypes to describe senescence in wheat. Genes 2021, 12, 909. [Google Scholar] [CrossRef]
- Cao, J.; Liu, H.; Tan, S.; Li, Z. Transcription Factors-Regulated Leaf Senescence: Current Knowledge, Challenges and Approaches. Int. J. Mol. Sci. 2023, 24, 9245. [Google Scholar] [CrossRef]
- Borrill, P.; Harrington, S.A.; Simmonds, J.; Uauy, C. Identification of transcription factors regulating senescence in wheat through gene regulatory network modelling. Plant Physiol. 2019, 180, 1740–1755. [Google Scholar] [CrossRef]
- Yolcu, S.; Li, X.; Li, S.; Kim, Y.J. Beyond the genetic code in leaf senescence. J. Exp. Bot. 2018, 69, 801–810. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Guo, P.; Xia, X.; Guo, H.; Li, Z. Multiple layers of regulation on leaf senescence: New advances and perspectives. Front. Plant Sci. 2021, 12, 788996. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.H.; Lyu, J.I.; Woo, H.R.; Lim, P.O. New insights into the regulation of leaf senescence in Arabidopsis. J. Exp. Bot. 2018, 69, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Wang, J. Mitochondrial functions in leaf senescence: Insights from metabolic and cellular dynamics. J. Plant Physiol. 2023, 271, 153681. [Google Scholar]
- Nousis, L.; Kanavaros, P.; Barbouti, A. Oxidative Stress-Induced Cellular Senescence: Is Labile Iron the Connecting Link? Antioxidants 2023, 12, 1250. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, W.; Pang, J.; Zhou, M.; Liu, J.; Zhao, J.; Yang, M. Integrated physiological and metabolomic analyses reveal changes during the natural senescence of Quercus mongolica leaves. PLoS ONE 2023, 18, e0289272. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Shekoofa, A.; Pessarakli, M. Reactive Oxygen Species (ROS) generation and detoxifying in plants. J. Plant Nutr. 2014, 37, 1573–1585. [Google Scholar] [CrossRef]
- Li, W.; Yang, Y.; Yu, L. The role of chloroplasts in leaf senescence and related signaling pathways. Int. J. Mol. Sci. 2023, 24, 1147. [Google Scholar]
- Buet, A.; Costa, M.L.; Martínez, D.E.; Guiamet, J.J. Chloroplast protein degradation in senescing leaves: Proteases and lytic compartments. Front. Plant Sci. 2019, 10, 747. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Hönig, M.; Plíhalová, L.; Husičková, A.; Nisler, J.; Doležal, K. Role of Cytokinins in Senescence, Antioxidant Defence and Photosynthesis. Int. J. Mol. Sci. 2018, 19, 4045. [Google Scholar] [CrossRef]
- Chen, C.; Galon, Y.; Rahmati Ishka, M.; Malihi, S.; Shimanovsky, V.; Twito, S.; Miller, G. Ascorbate peroxidase 6 delays the onset of age-dependent leaf senescence. Plant Physiol. 2021, 185, 441–456. [Google Scholar]
- Ďúranová, H.; Šimora, V.; Ďurišová, Ľ.; Olexiková, L.; Kovár, M.; Požgajová, M. Modifications in ultrastructural characteristics and redox status of plants under environmental stress: A review. Plants 2023, 12, 1666. [Google Scholar] [CrossRef] [PubMed]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [PubMed]
- Tefera, A.; Kebede, M.; Tadesse, K.; Getahun, T. Morphological, physiological, and biochemical characterization of drought-tolerant wheat (Triticum spp.) varieties. Int. J. Agron. 2021, 2021, 8811749. [Google Scholar] [CrossRef]
- Gan, S.S. Hypothesis: The subcellular senescence sequence of a mesophyll cell mirrors the cell origin and evolution. Mol. Hortic. 2022, 2, 27. [Google Scholar] [CrossRef]
- Tamary, E.; Nevo, R.; Naveh, L.; Levin-Zaidman, S.; Kiss, V.; Savidor, A.; Adam, Z. Chlorophyll catabolism precedes changes in chloroplast structure and proteome during leaf senescence. Plant Direct 2019, 3, e00127. [Google Scholar] [CrossRef]
- Yumoto, G.; Nishio, H.; Muranaka, T.; Sugisaka, J.; Honjo, M.N.; Kudoh, H. Seasonal switching of integrated leaf senescence controls in an evergreen perennial Arabidopsis. Nat. Commun. 2024, 15, 4719. [Google Scholar] [CrossRef]
- Xie, Y.; Ma, M.; Liu, Y.; Wang, B.; Wei, H.; Kong, D.; Wang, H. Arabidopsis FHY3 and FAR1 function in age gating of leaf senescence. Front. Plant Sci. 2021, 12, 770060. [Google Scholar] [CrossRef]
- Zhao, X.; Zhuang, W.; Zhang, Y. The relationship between leaf senescence and stress tolerance in plants. J. Plant Growth Regul. 2023, 42, 1–15. [Google Scholar]
- Yan, F.; Yu, Z.; Shi, Y. Optimized border irrigation delays winter wheat flag leaf senescence and promotes grain filling. Front. Plant Sci. 2023, 14, 1051323. [Google Scholar] [CrossRef]
- Allahverdiyev, T.I. Physiological traits of durum wheat (Triticum durum Desf.) and bread wheat (Triticum aestivum L.) genotypes under drought stress. Agric. Sci. 2015, 6, 848–859. [Google Scholar]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Deshmukh, P.; Sairam, R.; Shukla, D. Measurement of ion leakage as a screening technique for drought resistance in wheat genotypes. Indian J. Plant Physiol. 1991, 35, 89–91. [Google Scholar]
- Wang, H.F.; Zhong, X.H.; Shi, W.Y.; Guo, B. Study of malondialdehyde (MDA) content, superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) activities in chickens infected with avian infectious bronchitis virus. Afr. J. Biotechnol. 2011, 10, 9213–9217. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Zelinova, V.; Mistrik, I.; Palove-Balang, P.; Tamas, L. Peroxidase activity against guaiacol, NADH, chlorogenic acid, ferulic acid and coniferyl alcohol in root tips of Lotus japonicus and L. corniculatus grown under low pH and aluminium stress. Biologia 2010, 65, 279–283. [Google Scholar] [CrossRef]
- Gechev, T.; Gadjiev, I.; Van Breusagem, F.; Inze, D.; Dukiandjiev, S.; Toneva, V.; Minkov, I. Hydrogen peroxide protects tobacco from oxidative stress by inducing a set of antioxidant enzymes. Cell Mol. Life Sci. 2002, 59, 708–714. [Google Scholar] [CrossRef]
- Kumar, G.M.; Knowles, N.R. Changes in lipid peroxidation and lipolytic and free-radical scavenging enzyme activities during aging and sprouting of potato (Solanum tuberosum) seed-tubers. Plant Physiol. 1993, 102, 115–124. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kuo, J. Processing plant tissues for ultrastructural study. Electron Microsc. Methods Protoc. 2014, 1117, 39–55. [Google Scholar]
- Morikawa, S.; Sato, A.; Ezaki, T. A simple, one-step polychromatic staining method for epoxy-embedded semithin tissue sections. Microscopy 2018, 67, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Španić, V.; Šunić, K.; Duvnjak, J.; Hu, Y.G.; Katanić, Z. Chlorophyll a fluorescence during flag leaf senescence of field-grown winter wheat plants under drought conditions. Ann. Appl. Biol. 2023, 183, 80–92. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A.; Krupinska, K.; Shimakawa, G. The impact of photosynthesis on initiation of leaf senescence. Physiol. Plant. 2019, 166, 148–164. [Google Scholar] [CrossRef]
- Pic, E.; de La Serve, B.T.; Tardieu, F.; Turc, O. Leaf senescence induced by mild water deficit follows the same sequence of macroscopic, biochemical, and molecular events as monocarpic senescence in pea. Plant Physiol. 2002, 128, 236–246. [Google Scholar] [CrossRef]
- Liu, Y.N.; Xu, Q.Z.; Li, W.C.; Yang, X.H.; Zheng, Q.; Li, B.; Li, H.W. Long-term high light stress induces leaf senescence in wheat (Triticum aestivum L.). Photosynthetica 2019, 57, 3. [Google Scholar] [CrossRef]
- Huang, Q.N.; Shi, Y.F.; Zhang, X.B.; Song, L.X.; Feng, B.H.; Wang, H.M.; Xu, X.; Li, X.H.; Guo, D.; Wu, J.L. Single base substitution in OsCDC48 is responsible for premature senescence and death phenotype in rice. J. Integr. Plant Biol. 2016, 58, 12–28. [Google Scholar] [CrossRef]
- Maqsood, M.F.; Shahbaz, M.; Kanwal, S.; Kaleem, M.; Shah, S.M.R.; Luqman, M.; Farhat, F. Methionine promotes the growth and yield of wheat under water deficit conditions by regulating the antioxidant enzymes, reactive oxygen species, and ions. Life 2022, 12, 969. [Google Scholar] [CrossRef]
- Díaz-Mendoza, M.; Velasco-Arroyo, B.; Santamaría, M.E.; González-Melendi, P.; Martínez, M.; Díaz, I. Plant senescence and proteolysis: Two processes with one destiny. Genet. Mol. Biol. 2016, 39, 329–338. [Google Scholar] [CrossRef]
- Christiansen, M.W.; Gregersen, P.L. Members of the barley NAC transcription factor gene family show differential co-regulation with senescence-associated genes during senescence of flag leaves. J. Exp. Bot. 2014, 65, 4009–4022. [Google Scholar] [CrossRef]
- Hollmann, J.; Gregersen, P.L.; Krupinska, K. Identification of predominant genes involved in regulation and execution of senescence-associated nitrogen remobilization in wheat. J. Exp. Bot. 2014, 65, 4101–4113. [Google Scholar] [CrossRef]
- Roberts, I.N.; Caputo, C.; Criado, M.V.; Funk, C. Senescence-associated proteases in plants. Physiol. Plant. 2012, 145, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Hongwei, L.; Gui, W.; Shudong, L.; Qiang, A.; Zheng, Q.; Bin, L.; Zhensheng, L. Comparative changes in the antioxidant system in the flag leaf of early and normally senescing near-isogenic lines of wheat (Triticum aestivum L.). Plant Cell Rep. 2014, 33, 1109–1120. [Google Scholar]
- Karatas, I.; Ozturk, L.; Ersahin, Y.; Okatan, Y. Effects of auxin on photosynthetic pigments and some enzyme activities during dark-induced senescence of Tropaeolum leaves. Pak. J. Bot. 2010, 42, 1881–1888. [Google Scholar]
- Lambert, R.; Quiles, F.A.; Gálvez-Valdivieso, G.; Piedras, P. Nucleases activities during French bean leaf aging and dark-induced senescence. J. Plant Physiol. 2017, 218, 235–242. [Google Scholar] [CrossRef]
- Laus, M.N.; De Santis, M.A.; Flagella, Z.; Soccio, M. Changes in Antioxidant Defence System in Durum Wheat under Hyperosmotic Stress: A Concise Overview. Plants 2022, 11, 98. [Google Scholar] [CrossRef]
- Huseynova, I.M. Photosynthetic characteristics and enzymatic antioxidant capacity of leaves from wheat cultivars exposed to drought. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 1516–1523. [Google Scholar] [CrossRef]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Abd_Allah, E.F.; Ahmad, P.; Corpas, F.J. Assessment of subcellular ROS and NO metabolism in higher plants: Multifunctional signaling molecules. Antioxidants 2019, 8, 641. [Google Scholar] [CrossRef]
- Mao, H.; Chen, M.; Su, Y.; Wu, N.; Yuan, M.; Yuan, S.; Brestic, M.; Zivcak, M.; Zhang, H.; Chen, Y. Comparison on photosynthesis and antioxidant defense Systems in Wheat with different Ploidy levels and Octoploid Triticale. Int. J. Mol. Sci. 2018, 19, 3006. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Prochazkova, D.; Wilhelmova, N. Leaf senescence and activities of the antioxidant enzymes. Biol. Plant. 2007, 51, 401–406. [Google Scholar] [CrossRef]
- Jiménez, A.; Hernández, J.A.; Pastori, G.; del Río, L.A.; Sevilla, F. Role of the ascorbate-glutathione cycle of mitochondria and peroxisomes in the senescence of pea leaves. Plant Physiol. 1998, 118, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Aliyeva, D.R.; Gurbanova, U.A.; Rzayev, F.H.; Gasimov, E.K.; Huseynova, I.M. Biochemical and ultrastructural changes in wheat plants during drought stress. Biochemistry 2023, 88, 1944–1955. [Google Scholar] [CrossRef] [PubMed]
- Saed-Moucheshi, A.; Sohrabi, F.; Fasihfar, E.; Baniasadi, F.; Riasat, M.; Mozafari, A.A. Superoxide dismutase (SOD) as a selection criterion for triticale grain yield under drought stress: A comprehensive study on genomics and expression profiling, bioinformatics, heritability, and phenotypic variability. BMC Plant Biol. 2021, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhou, L.; Liu, J.; Du, X.; Huang, F.; Pan, G.; Cheng, F. Relationship of ROS accumulation and superoxide dismutase isozymes in developing anther with floret fertility of rice under heat stress. Plant Physiol. Biochem. 2018, 122, 90–101. [Google Scholar] [CrossRef]
- Mohammadi, M.; Tavakoli, A.; Pouryousef, M.; Fard, E.M. Study the effect of 24-epibrassinolide application on the Cu/Zn-SOD expression and tolerance to drought stress in common bean. Physiol. Mol. Biol. Plants 2020, 23, 459–474. [Google Scholar] [CrossRef]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The roles of environmental factors in regulation of oxidative stress in plant. Biomed Res. Int. 2019, 2019, 435–455. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Pakniyat, H.; Pirasteh-Anosheh, H.; Azooz, M.M. Role of ROS as signaling molecules in plants. In Oxidative Damage to Plants; Academic Press: Cambridge, MA, USA, 2014; pp. 585–620. [Google Scholar]
- Awan, S.A.; Khan, I.; Rizwan, M.; Zhang, X.; Brestic, M.; Khan, A.; El-Sheikh, M.A.; Alyemeni, M.N.; Ali, S.; Huang, L. Exogenous abscisic acid and jasmonic acid restrain polyethylene glycol-induced drought by improving the growth and antioxidative enzyme activities in pearl millet. Physiol. Plant. 2020, 147, 125–136. [Google Scholar] [CrossRef]
- Sheoran, S.; Thakur, V.; Narwal, S.; Turan, R.; Mamrutha, H.; Singh, V.; Tiwari, V.; Sharma, I. Differential activity and expression profile of antioxidant enzymes and physiological changes in wheat (Triticum aestivum L.) under drought. Appl. Biochem. Biotechnol. 2015, 177, 1282–1298. [Google Scholar] [CrossRef]
- Wang, W.; Xia, M.; Chen, J.; Yuan, R.; Deng, F.; Shen, F. Gene expression characteristics and regulation mechanisms of superoxide dismutase and its physiological roles in plants under stress. Biochemistry 2018, 81, 465–480. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, L.; He, Y.; Zhang, H.; Li, W.; Chen, H.; Yin, J. Genome-wide identification and transcriptional expression analysis of superoxide dismutase (SOD) family in wheat (Triticum aestivum). PeerJ 2019, 7, e8062. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Huseynova, I.; Aliyeva, D.; Aliyev, J. Subcellular localization and responses of superoxide dismutase isoforms in local wheat varieties subjected to continuous soil drought. Plant Physiol. Biochem. 2014, 81, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Huseynova, I.; Aliyeva, D.; Mammadov, A.; Aliyev, J. Hydrogen peroxide generation and antioxidant enzyme activities in the leaves and roots of wheat cultivars subjected to long-term soil drought stress. Photosynth. Res. 2015, 1125, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Huseynova, I.; Rustamova, S.; Suleymanov, S.; Aliyeva, D.; Mammadov, A.; Aliyev, J. Drought-induced changes in photosynthetic apparatus and antioxidant components of wheat (Triticum durum Desf.) varieties. Photosynth. Res. 2016, 130, 215–223. [Google Scholar] [CrossRef]
- Scebba, F.; Sebastiani, L.; Vitagliano, C. Changes in activity of antioxidative enzymes in wheat (Triticum aestivum) seedlings under cold acclimation. Physiol. Plant. 1998, 104, 747–752. [Google Scholar] [CrossRef]
- Manoharan, K.; Karuppanapandian, T.; Sinha, P.B.; Prasad, R. Membrane degradation, accumulation of phosphatidic acid, stimulation of catalase activity and nuclear DNA fragmentation during 2,4-D-induced leaf senescence in mustard. J. Plant Biol. 2005, 48, 394–403. [Google Scholar] [CrossRef]
- Prochazkova, D.; Sairam, R.K.; Srivastava, G.C.; Singh, D.V. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Paluch-Lubawa, E.; Stolarska, E.; Sobieszczuk-Nowicka, E. Dark-induced barley leaf senescence—A crop system for studying senescence and autophagy mechanisms. Front. Plant Sci. 2021, 12, 635619. [Google Scholar] [CrossRef]
- Smart, C.M. Gene expression during leaf senescence. New Phytol. 1994, 126, 419–448. [Google Scholar] [CrossRef]
- Austin, J.R.; Frost, E.; Vidi, P.A.; Kessler, F.; Staehelin, L.A. Plastoglobules are lipoprotein subcompartments of the chloroplast that are permanently coupled to thylakoid membranes and contain biosynthetic enzymes. Plant Cell 2006, 18, 1693–1703. [Google Scholar] [CrossRef]
- Challabathula, D.; Analin, B.; Mohanan, A.; Bakka, K. Differential modulation of photosynthesis, ROS and antioxidant enzyme activities in stress-sensitive and -tolerant rice cultivars during salinity and drought upon restriction of COX and AOX pathways of mitochondrial oxidative electron transport. J. Plant Physiol. 2022, 268, 153583. [Google Scholar] [CrossRef]
- Vollenweider, P.; Ottiger, M.; Günthardt-Goerg, M.S. Validation of leaf ozone symptoms in natural vegetation using microscopical methods. Environ. Pollut. 2003, 124, 101–118. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Direction | Sequences |
|---|---|---|
| MnSOD | F | AACATCTGGAAGGTGGTGAACT |
| R | AACTCAAGAGCGAGCGAAGTA | |
| CuZnSOD | F | CTCCATGAGTTCGGTGACAT |
| R | GACGGACTTCATCTTCTGGT | |
| FeSOD | F | GAATTCCACTGGGGAAGCATC |
| R | GTAAGCGTGCTCCCAAACGTC | |
| Elf1-α | F | CAGATTGGCAACGGCTACG |
| R | CGGACAGCAAAACGACCAAG |
| Trait | Source of Variance | NumDF | DenDF | F Value | ProbF |
|---|---|---|---|---|---|
| Cla | Treatment | 4 | 5 | 7.009887 | 0.027812 |
| Genotype | 3 | 95 | 76.88884 | 2.55 × 10−25 | |
| Treatment × Genotype | 12 | 95 | 2.972035 | 0.001478 | |
| Clb | Treatment | 4 | 5 | 2.55842 | 0.165371 |
| Genotype | 3 | 95 | 7.240645 | 0.000199 | |
| Treatment × Genotype | 12 | 95 | 0.988862 | 0.465342 | |
| MDA | Treatment | 4 | 5 | 1.717407 | 0.281804 |
| Genotype | 3 | 95 | 18.55237 | 1.49 × 10−9 | |
| Treatment × Genotype | 12 | 95 | 1.495899 | 0.139189 | |
| MSI | Treatment | 4 | 5 | 2.939509 | 0.13392 |
| Genotype | 3 | 95 | 16.68884 | 8.69 × 10−9 | |
| Treatment × Genotype | 12 | 95 | 2.65968 | 0.004059 | |
| Total protein | Treatment | 4 | 5 | 5.457763 | 0.045485 |
| Genotype | 3 | 95 | 14.97228 | 4.67 × 10−8 | |
| Treatment × Genotype | 12 | 95 | 13.69248 | 5.72 × 10−16 | |
| SOD activity | Treatment | 4 | 5 | 5.998624 | 0.037898 |
| Genotype | 3 | 95 | 14.36139 | 8.63 × 10−8 | |
| Treatment × Genotype | 12 | 95 | 11.57891 | 5.56 × 10−14 | |
| APX activity | Treatment | 4 | 5 | 1.595833 | 0.307119 |
| Genotype | 3 | 95 | 33.84412 | 5.74 × 10−15 | |
| Treatment × Genotype | 12 | 95 | 6.462609 | 2.79 × 10−8 | |
| BPX activity | Treatment | 4 | 5 | 6.381422 | 0.033558 |
| Genotype | 3 | 95 | 80.75748 | 4.86 × 10−26 | |
| Treatment × Genotype | 12 | 95 | 13.97545 | 3.2 × 10−16 | |
| GPX activity | Treatment | 4 | 5 | 27.33506 | 0.001356 |
| Genotype | 3 | 95 | 191.9561 | 3.45 × 10−40 | |
| Treatment × Genotype | 12 | 95 | 13.50056 | 8.52 × 10−16 | |
| CAT activity | Treatment | 4 | 5 | 9.505715 | 0.014778 |
| Genotype | 3 | 95 | 20.89745 | 1.76 × 10−10 | |
| Treatment × Genotype | 12 | 95 | 3.890105 | 7.53 × 10−5 | |
| CuZnSOD expression | Treatment | 4 | 5 | 8.020674 | 0.021115 |
| Genotype | 3 | 95 | 19.27962 | 7.61 × 10−10 | |
| Treatment × Genotype | 12 | 95 | 7.145308 | 3.98 × 10−9 | |
| FeSOD expression | Treatment | 4 | 5 | 29.16198 | 0.001164 |
| Genotype | 3 | 95 | 10.08343 | 7.88 × 10−6 | |
| Treatment × Genotype | 12 | 95 | 4.693792 | 5.83 × 10−6 | |
| MnSOD expression | Treatment | 4 | 5 | 21.59548 | 0.002355 |
| Genotype | 3 | 95 | 10.99319 | 2.92 × 10−6 | |
| Treatment × Genotype | 12 | 95 | 1.659175 | 0.088501 |
| Chlorophyll a (mg/g FW) | ||||||
|---|---|---|---|---|---|---|
| 7 DAAs | 14 DAAs | 21 DAAs | 28 DAAs | 35 DAAs | ||
| Vugar | control | 6.89 ± 0.56 | 7.18 ± 0.55 | 6.4 ± 0.53 | 5.8 ± 0.44 | 3.5 ± 0.26 |
| stress | 6.51 ± 0.52 | 6.18 ± 0.49 | 4.89 ± 0.39 | 3.5 ± 0.28 | 2.3 ± 0.18 | |
| Tartar | control | 7.43 ± 0.61 | 7.09 ± 0.56 | 6.07 ± 0.49 | 4.8 ± 0.37 | 3.3 ± 0.26 |
| stress | 6.82 ± 0.55 | 6.06 ± 0.48 | 3.23 ± 0.26 | 2.2 ± 0.18 | 1.3 ± 0.1 | |
| Gyrmyzy gul 1 | control | 5.57 ± 0.44 | 5.04 ± 0.41 | 4.35 ± 0.35 | 3.5 ± 0.28 | 2.1 ± 0.17 |
| stress | 5.09 ± 0.41 | 4.45 ± 0.18 | 3.02 ± 0.19 | 2.5 ± 0.2 | 1.8 ± 0.14 | |
| Tale 38 | control | 5.12 ± 0.41 | 5.06 ± 0.4 | 4.6 ± 0.37 | 2.9 ± 0.23 | 1.9 ± 0.15 |
| stress | 4.49 ± 0.36 | 4.04 ± 0.32 | 2.21 ± 0.18 | 1.69 ± 0.14 | 1.05 ± 0.08 | |
| Chlorophyll b (mg/g FW) | ||||||
| 7 DAAs | 14 DAAs | 21 DAAs | 28 DAAs | 35 DAAs | ||
| Vugar | control | 6.51 ± 0.52 | 1.91 ± 0.16 | 1.5 ± 0.12 | 1.2 ± 0.09 | 1.1 ± 0.08 |
| stress | 1.87 ± 0.16 | 1.65 ± 0.12 | 1.03 ± 0.08 | 0.7 ± 0.06 | 0.5 ± 0.04 | |
| Tartar | control | 2.25 ± 0.18 | 2.15 ± 0.17 | 2.02 ± 0.16 | 1.6 ± 0.14 | 1.1 ± 0.07 |
| stress | 1.81 ± 0.14 | 0.98 ± 0.08 | 0.6 ± 0.05 | 0.4 ± 0.03 | 0.19 ± 0.02 | |
| Gyrmyzy gul 1 | control | 1.98 ± 0.16 | 1.82 ± 0.15 | 1.5 ± 0.12 | 1.2 ± 0.1 | 0.8 ± 0.06 |
| stress | 1.66 ± 0.13 | 1.61 ± 0.13 | 0.9 ± 0.07 | 0.7 ± 0.06 | 0.4 ± 0.03 | |
| Tale 38 | control | 2.18 ± 0.17 | 2.04 ± 0.16 | 1.65 ± 0.13 | 0.88 ± 0.1 | 0.6 ± 0.05 |
| stress | 1.56 ± 0.12 | 0.85 ± 0.07 | 0.61 ± 0.05 | 0.5 ± 0.04 | 0.2 ± 0.02 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isgandarova, T.Y.; Rustamova, S.M.; Aliyeva, D.R.; Rzayev, F.H.; Gasimov, E.K.; Huseynova, I.M. Antioxidant and Ultrastructural Alterations in Wheat During Drought-Induced Leaf Senescence. Agronomy 2024, 14, 2924. https://doi.org/10.3390/agronomy14122924
Isgandarova TY, Rustamova SM, Aliyeva DR, Rzayev FH, Gasimov EK, Huseynova IM. Antioxidant and Ultrastructural Alterations in Wheat During Drought-Induced Leaf Senescence. Agronomy. 2024; 14(12):2924. https://doi.org/10.3390/agronomy14122924
Chicago/Turabian StyleIsgandarova, Turana Y., Samira M. Rustamova, Durna R. Aliyeva, Fuad H. Rzayev, Eldar K. Gasimov, and Irada M. Huseynova. 2024. "Antioxidant and Ultrastructural Alterations in Wheat During Drought-Induced Leaf Senescence" Agronomy 14, no. 12: 2924. https://doi.org/10.3390/agronomy14122924
APA StyleIsgandarova, T. Y., Rustamova, S. M., Aliyeva, D. R., Rzayev, F. H., Gasimov, E. K., & Huseynova, I. M. (2024). Antioxidant and Ultrastructural Alterations in Wheat During Drought-Induced Leaf Senescence. Agronomy, 14(12), 2924. https://doi.org/10.3390/agronomy14122924