
Metabolomic and Transcriptomic Insights into Anthocyanin Biosynthesis in ‘Ziyan’ Tea Plants under Varied Photoperiod and Temperature Conditions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Determination of Total Anthocyanin Content
2.3. Metabolome Analysis
2.4. RNA Extraction and Transcriptome Sequencing
2.5. Quality Assessment and Differentially Expressed Genes’ Identification
2.6. Quantitative Real-Time PCR Analysis
2.7. Data Analysis
3. Results
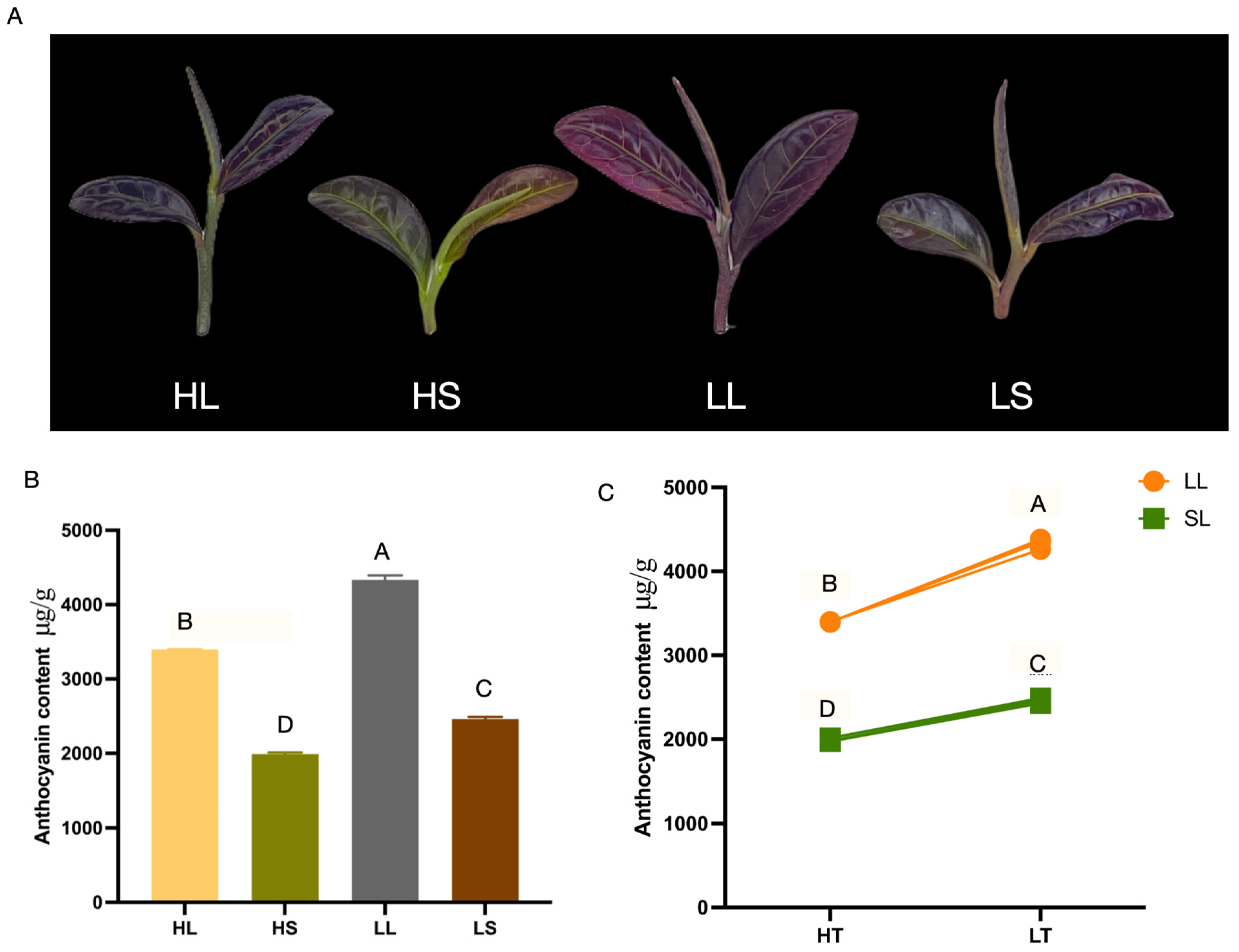
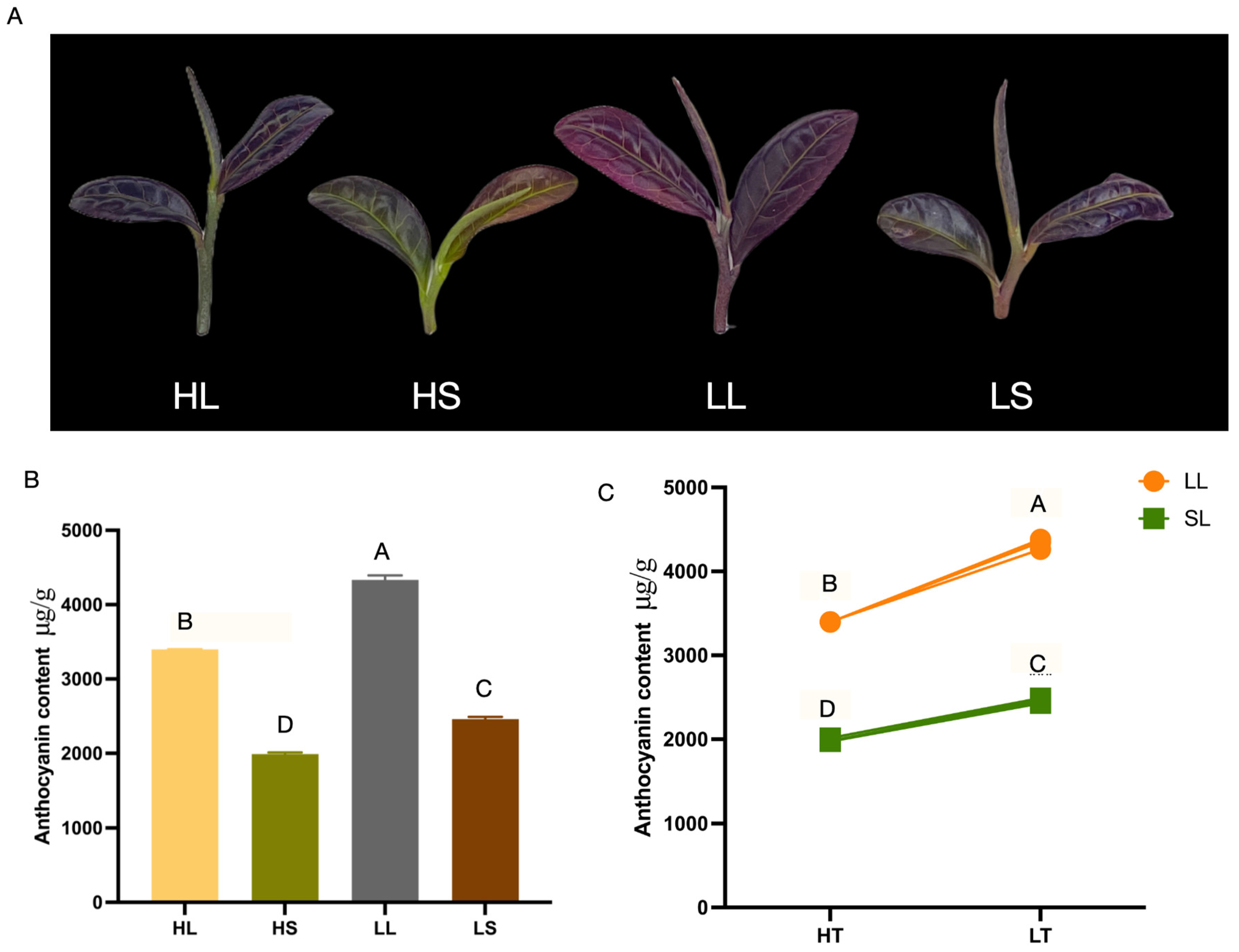
3.1. Low Temperatures and Extended Photoperiods Enhance the Total Anthocyanin Content of ‘Ziyan’ New Shoots
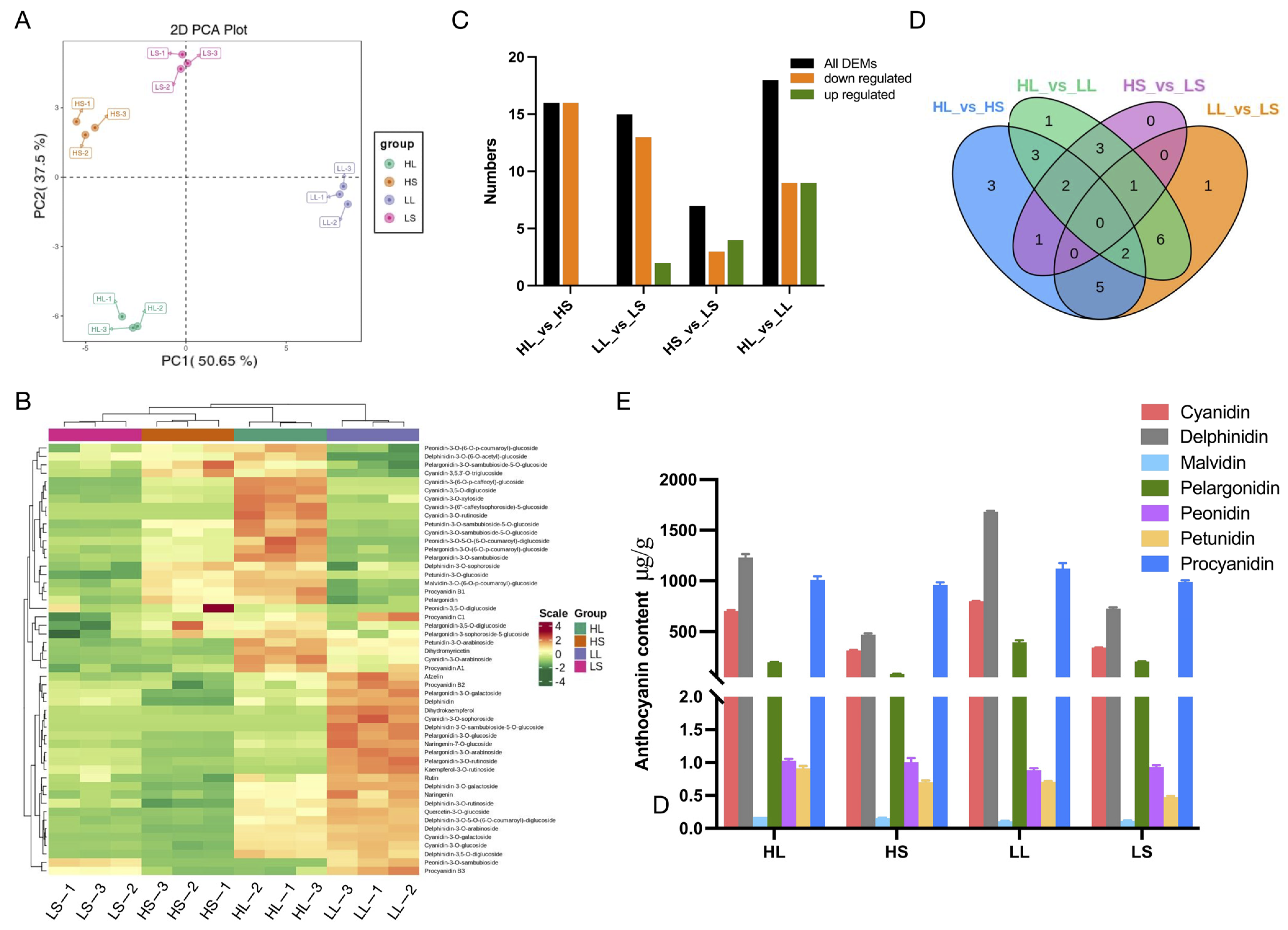
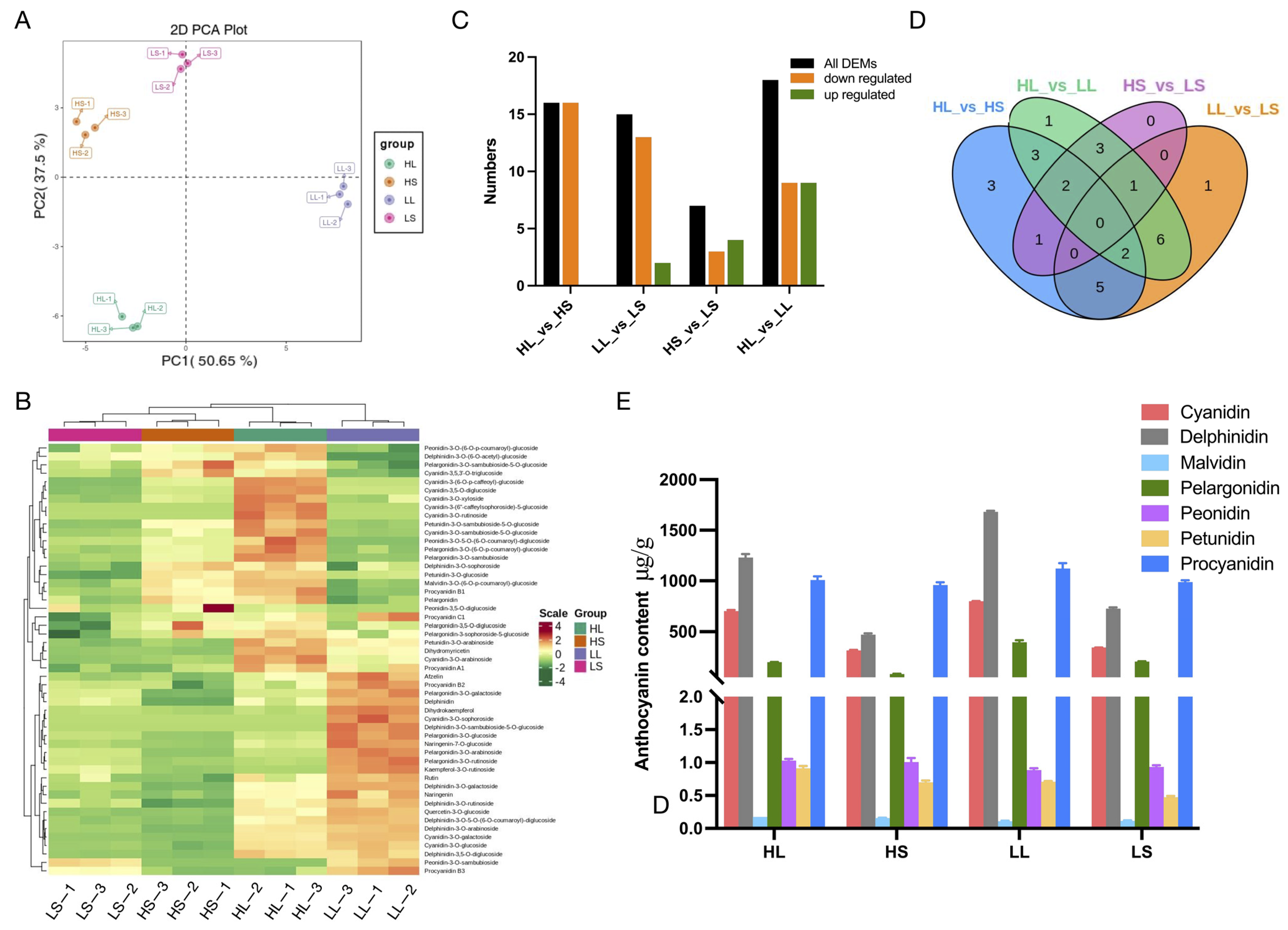
3.2. Anthocyanin Metabolites Analysis of ‘Ziyan’ New Shoots after Treatments with Different Photoperiods and Temperatures
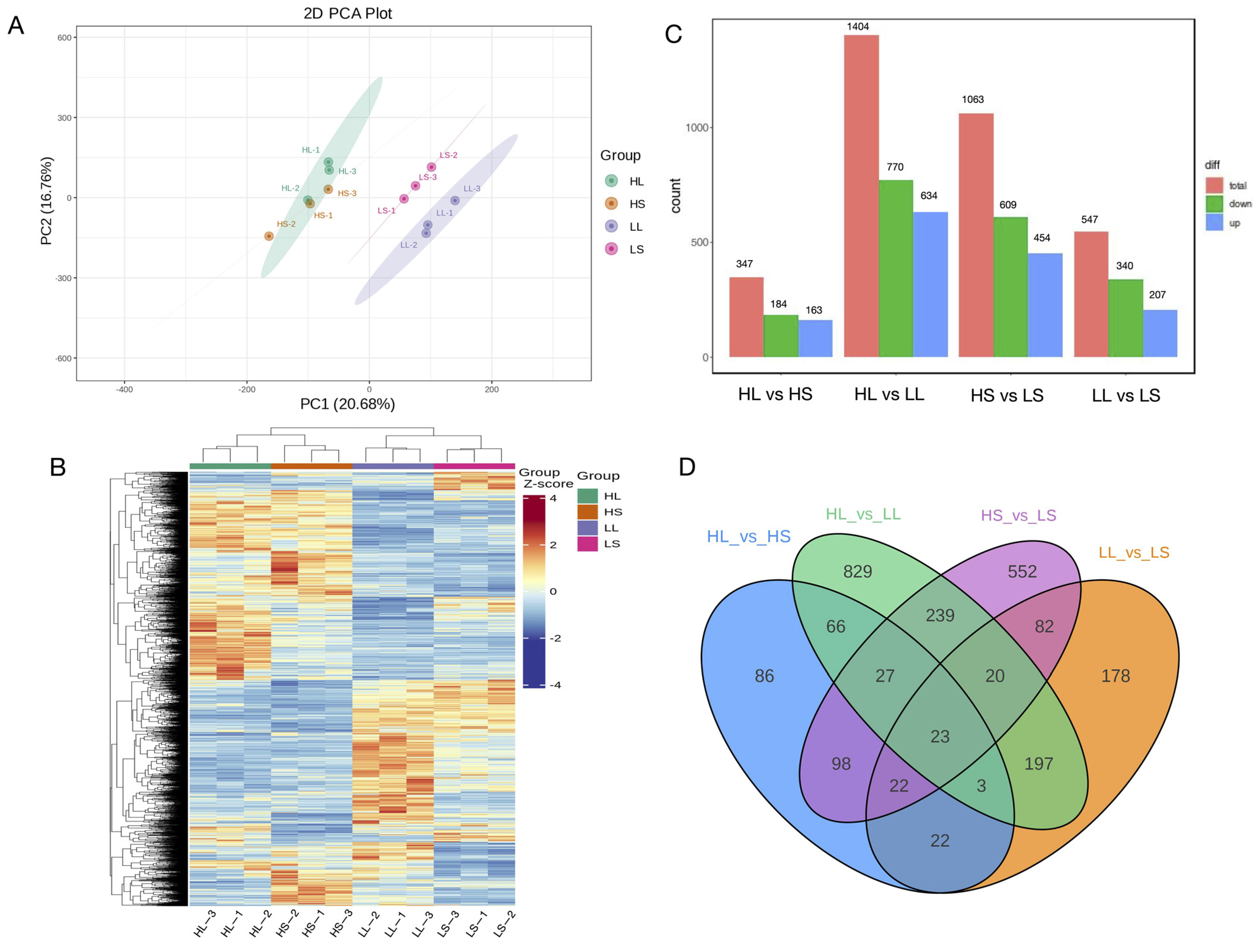
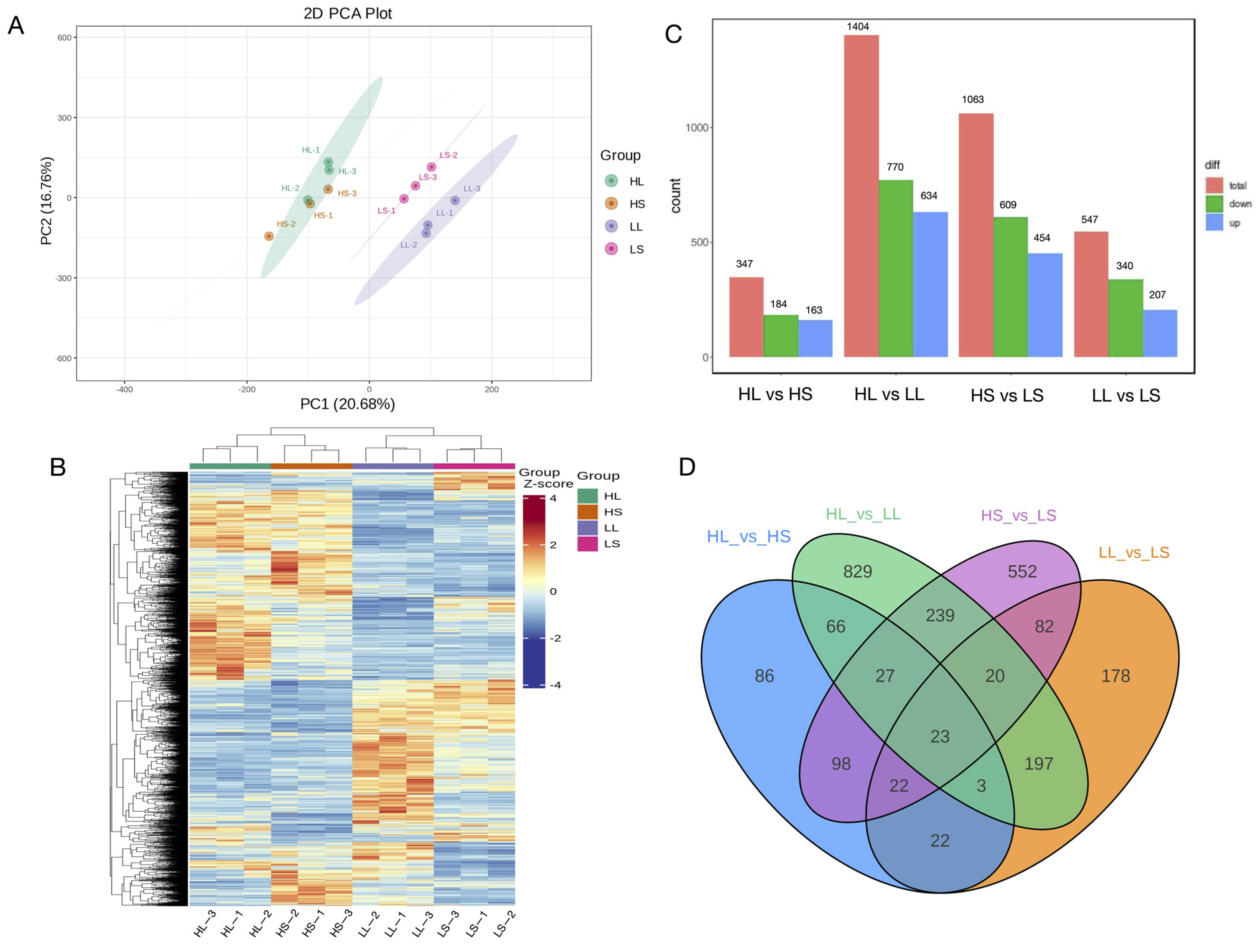
3.3. Differential Gene Expression Analysis under Different Photoperiod and Temperature Treatments
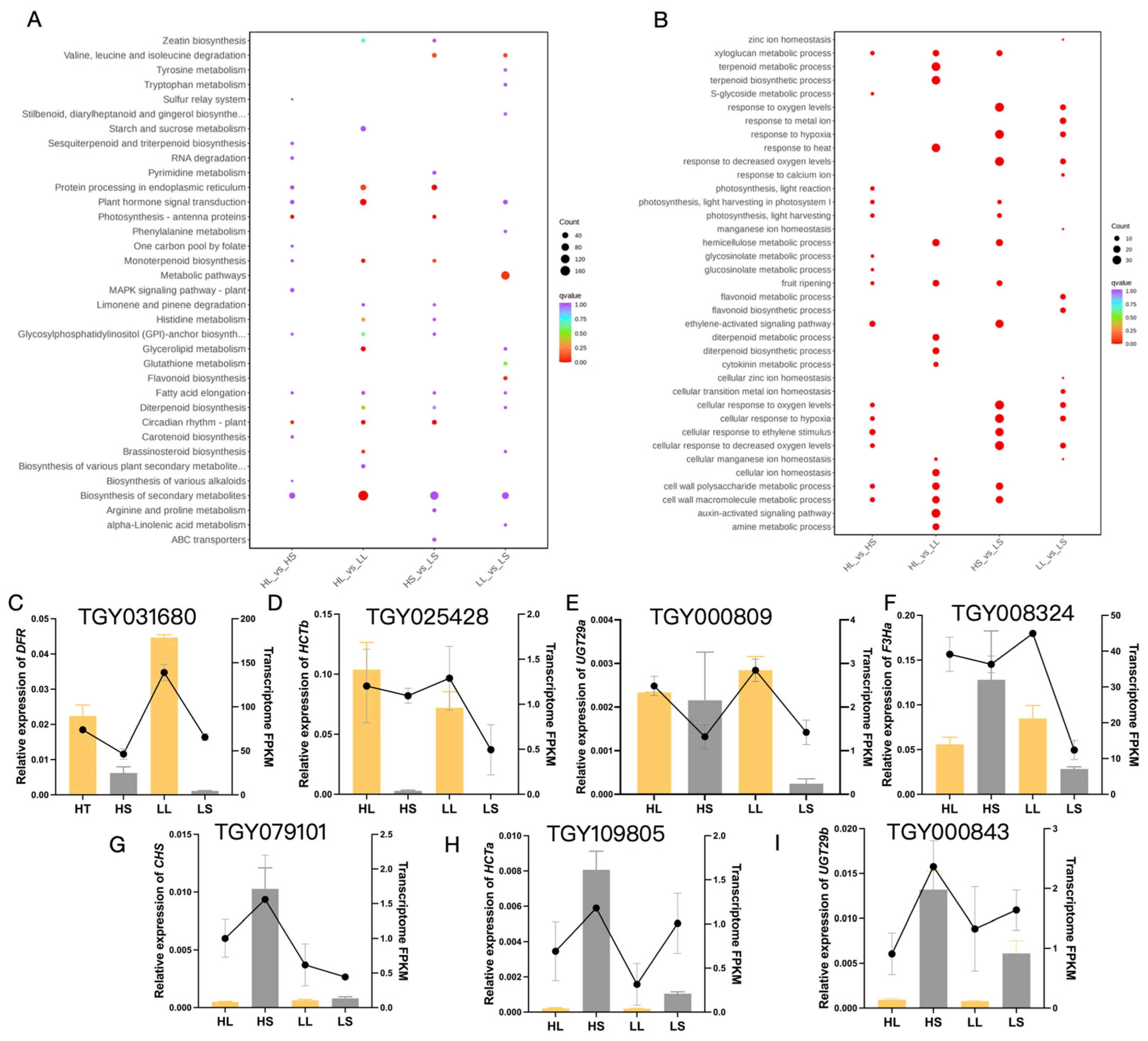
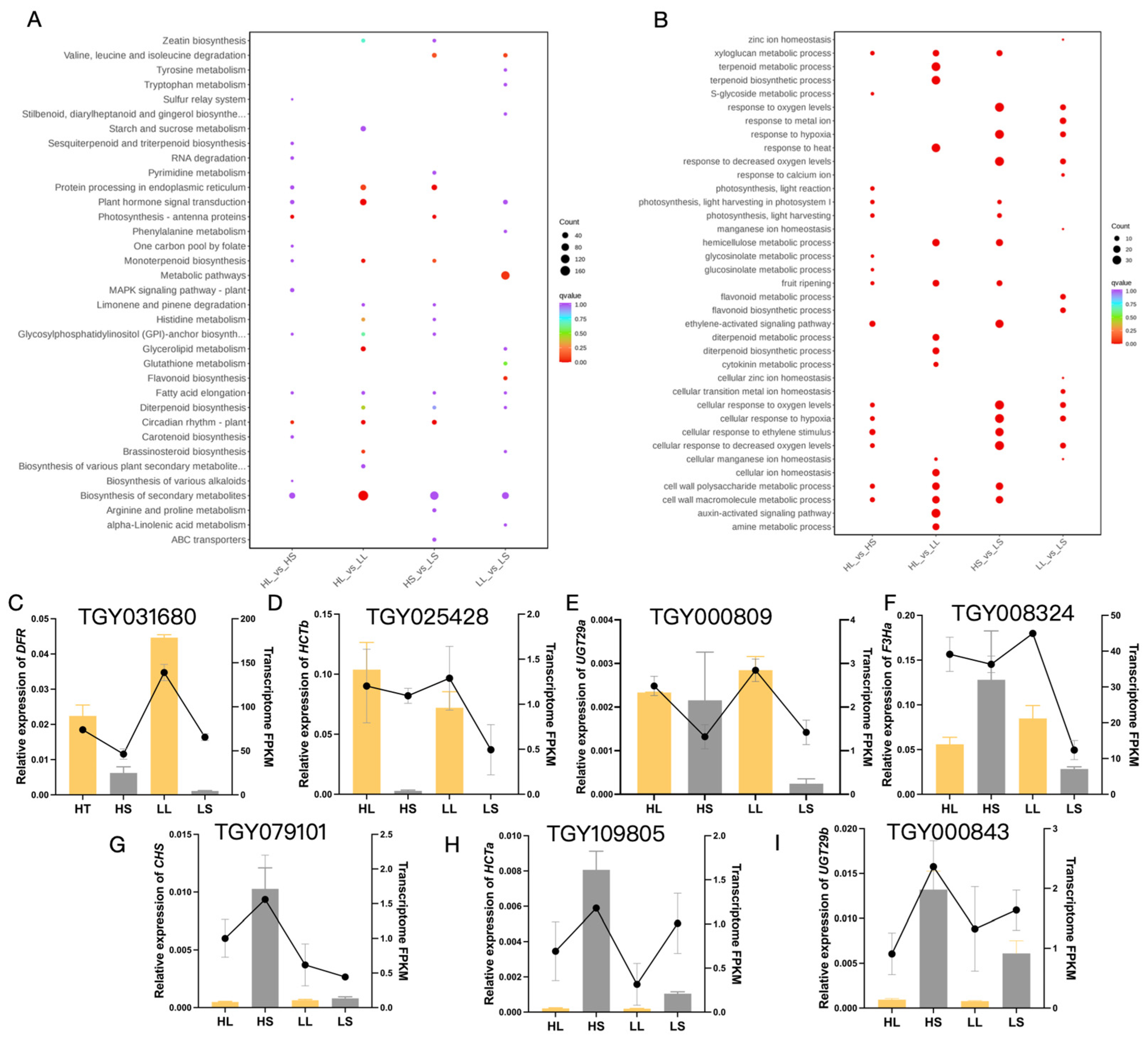
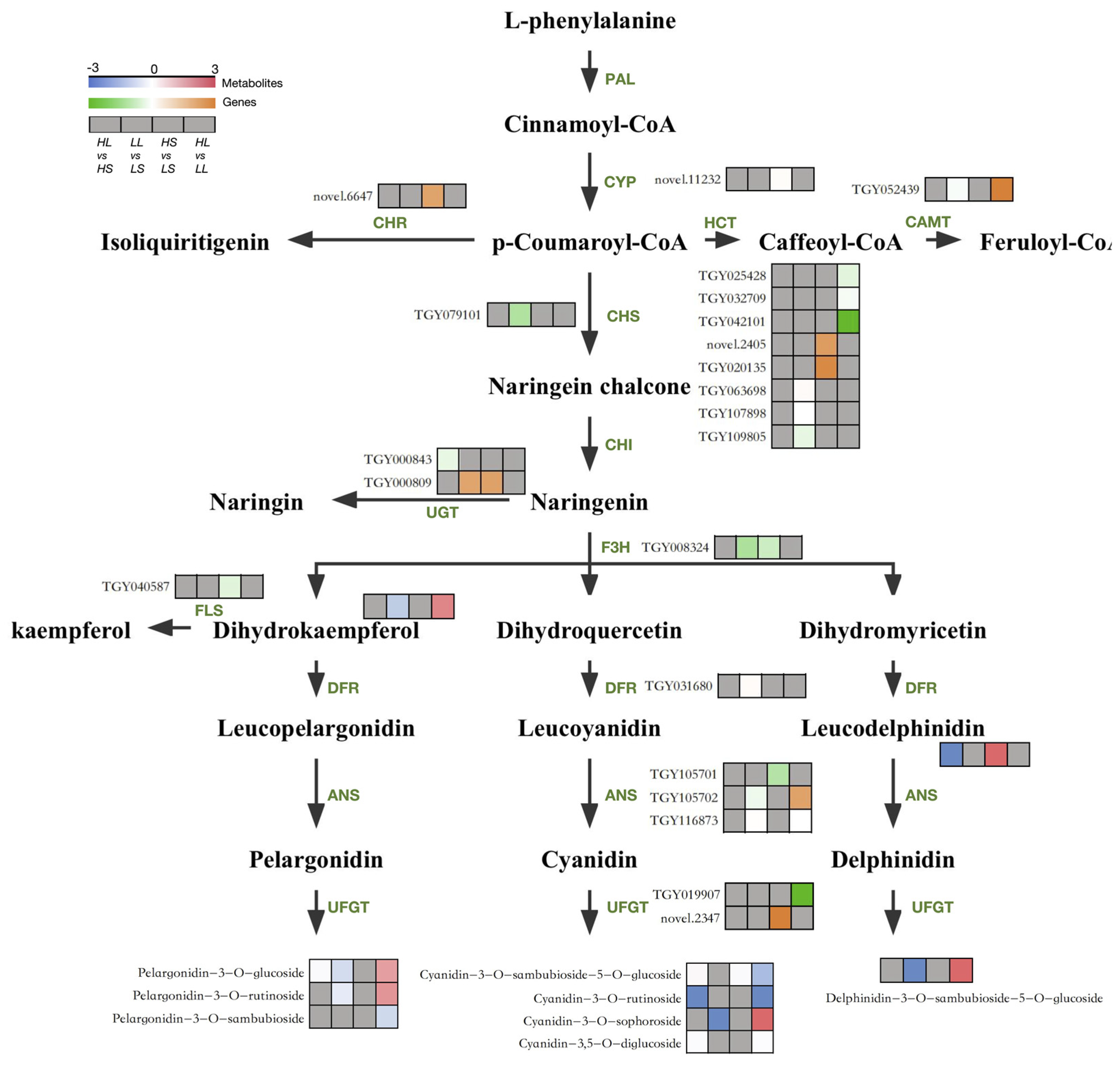
3.4. Analysis of DEGs Associated with the Anthocyanin Biosynthesis Pathway
3.5. Response of Anthocyanin Biosynthesis Metabolism in ‘Ziyan’ to Light and Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tan, L.; Zhang, P.; Cui, D.; Yang, X.; Zhang, D.; Yang, Y.; Chen, W.; Tang, D.; Tang, Q.; Li, P. Multi-omics analysis revealed anthocyanin accumulation differences in purple tea plants ‘Ziyan’, ‘Zijuan’ and their dark-purple hybrid. Sci. Hortic. 2023, 321, 112275. [Google Scholar] [CrossRef]
- Honma, D.; Tagashira, M.; Kanda, T.; Nesumi, A.; Maeda-Yamamoto, M. Anthocyanins from new red leaf tea ‘Sunrouge’. J. Agric. Food Chem. 2011, 59, 4779–4782. [Google Scholar] [CrossRef]
- Li, F.; Deng, X.; Huang, Z.; Zhao, Z.; Li, C.; Song, Q.; He, Y.; Niu, S. Integrated transcriptome and metabolome provide insights into flavonoid biosynthesis in ‘P113’, a new purple tea of Camellia tachangensis. Beverage Plant Res. 2023, 3, 3. [Google Scholar] [CrossRef]
- Cai, J.; Lv, L.; Zeng, X.; Zhang, F.; Chen, Y.; Tian, W.; Li, J.; Li, X.; Li, Y. Integrative Analysis of Metabolomics and Transcriptomics Reveals Molecular Mechanisms of Anthocyanin Metabolism in the Zikui Tea Plant (Camellia sinensis cv. Zikui). Int. J. Mol. Sci. 2022, 23, 4780. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shen, X.; Shoji, T.; Kanda, T.; Zhao, L. Characterization and Activity of Anthocyanins in Zijuan Tea (Camellia sinensis var. kitamura). J. Agric. Food Chem. 2013, 61, 3306–3310. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Wang, L.; Zhang, Y.; Ruan, L.; Li, H.; Wu, L.; Xu, L.; Zhang, C.; Zhou, X.; Cheng, H.; et al. A coupled role for CsMYB75 and CsGSTF1 in anthocyanin hyperaccumulation in purple tea. Plant J. 2019, 97, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Da silva, T.; Castilho, P.; SÁ-Nakanishi, A.; Vincente Seixas, F.; Dias, M.; Barros, L.; Ferreira, I.; Vracht, A.; Peralta, R. The inhibitory action of purple tea on in vivo starch digestion compared to other Camellia sinensis teas. Food Res. Int. 2021, 150, 110781. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Li, R.; Chen, J.; Wang, F.; Gao, Y.; Fu, Y.; Xu, Y.; Yin, J. Zijuan tea- based kombucha: Physicochemical, sensorial, and antioxidant profile. Food Chem. 2021, 30, 130322. [Google Scholar] [CrossRef]
- Ahammed, G.; Li, X. Hormonal regulation of health-promoting compounds in tea (Camellia sinensis L.). Plant Physiol. Biochem. 2022, 15, 390–400. [Google Scholar] [CrossRef]
- Jia, W.B.; Tang, Q.; Zou, Y.; Yang, Y.; Wu, W.; Xu, W. Investigating the antidepressant effect of Ziyan green tea on chronic unpredictable mild stress mice through fecal metabolomics. Front. Microbiol. 2023, 14, 1256142. [Google Scholar] [CrossRef]
- Lai, Y.; Li, S.; Tang, Q.; Li, H.; Chen, S. The dark-purple tea cultivar ‘Ziyan’ accumulates a large amount of delphinidin-related anthocyanins. J. Agric. Food Chem. 2016, 64, 2719–2726. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Yin, Z.; Tang, Q. Water extraction process of anthocyanins from ‘Ziyan’ tea and the antitumor activity of its extracts. Journal of Anhui agricultural university. Environ. Sci. 2019, 186, 115273. [Google Scholar] [CrossRef]
- Tan, L.; Yang, C.; Zhou, B.; Wang, L.; Zou, Y.; Chen, W.; Xia, T.; Tang, Q. Inheritance and quantitative trait loci analyses of the anthocyanins and catechins of Camellia sinensis cultivar ‘Ziyan’ with dark-purple leaves. Physiol. Plant 2020, 170, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Winkelshirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Ma, P.; Bian, X.; Yang, Q.; Guo, X.; Xie, Y. Biosynthesis Metabolic Pathway and Molecular Regulation of Plants Anthocyanin. Northwest Bot. J. 2014, 34, 1496–1506. [Google Scholar]
- Wang, L.; Dai, S.; Jin, X.; Huang, H.; Hong, Y. Advances in plant anthocyanin transport mechanism. J. Biotechnol. 2014, 30, 848–863. [Google Scholar] [CrossRef]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef]
- David, W. Regulation of flower pigmentation and growth: Multiple signaling pathways control anthocyanin synthesis in expanding petals. Plant Physiol. 2000, 110, 152–157. [Google Scholar] [CrossRef]
- Hu, K.; Han, K.; Dai, S. Regulation of Plant Anthocyanin Synthesis and Pigmentation by Enviromental Factors. Chin. Bull. Bot. 2010, 45, 307–317. [Google Scholar]
- Sheehan, M.; Farmer, P.; Brutnell, T. Structure and expression of maize phytochrome family homeologs. Genetics 2004, 167, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Albert, N.; Lewis, D.; Zhang, H.; Irving, L.; Jameson, P.; Davies, K. Light-induced vegetative anthocyanin pigmentation in Petunia. J. Exp. Bot. 2009, 60, 2191–2202. [Google Scholar] [CrossRef]
- Luo, Q.; Li, H.; Bai, B.; Yu, H.; You, J. Effect of Light on the Anthocyanin Biosynthesis and Expression of CHS and DFR in Rosa Chinensis ‘Spectra’. Mol. Plant Breed. 2013, 1, 126–131. [Google Scholar]
- Li, W.; Tan, L.; Zou, Y.; Tan, X.; Huang, J.; Chen, W.; Tang, Q. The Effects of Ultraviolet A/B Treatments on Anthocyanin Accumulation and Gene Expression in Dark-Purple Tea Cultivar ‘Ziyan’ (Camellia sinensis). Molecules 2020, 25, 354. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, M. A novel R2R3-MYB transcription factor regulates light-mediated floral and vegetative anthocyanin pigmentation patterns in Lilium regale. Mol. Breed. 2016, 36, 3. [Google Scholar] [CrossRef]
- Sun, B.; Zhu, Z.; Cao, P.; Chen, H.; Chen, C.; Zhou, X.; Mao, Y.; Lei, J.; Jiang, Y.; Meng, W.; et al. Purple foliage coloration in tea (Camellia sinensis L.) arises from activation of the R2R3-MYB transcription factor CsAN1. Sci. Rep. 2016, 6, 32534. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Honda, C.; Hatsuyama, Y.; Igarashi, M.; Bessho, H.; Moriguchi, T. Isolation and functional analysis of a MYB transcription factor gene that is a key regulator for the development of red coloration in apple skin. Plant Cell Physiol. 2007, 48, 958–970. [Google Scholar] [CrossRef]
- Fang, Z.; Wang, L.; Zhou, D.; Lin, Y.; Pan, S.; Espley, R.; Ye, X. Postharvest temperature and light treatments induce anthocyanin accumulation in peel of ‘Akihime’ plum (Prunus salicina Lindl.) via transcription factor PsMYB10.1. Postharvest Biol. Technol. 2021, 179, 111592. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, Z.; Jiang, S.; Xu, H.; Wang, Y.; Feng, S.; Chen, X. Synergistic effects of light and temperature on anthocyanin biosynthesis in callus cultures of red-fleshed apple (Malus sieversii f. niedzwetzkyana). Plant Cell Tissue Organ Cult. 2016, 127, 217–227. [Google Scholar] [CrossRef]
- Rowan, D.; Cao, M.; Lin, W.; Cooney, J.; Jensen, D.; Austin, P.; Hunt, M.; Norling, C.; Hellens, R.; Schaffer, R.; et al. Environmental regulation of leaf colour in red 35S:PAP1 Arabidopsis thaliana. New Phytol. 2009, 182, 102–115. [Google Scholar] [CrossRef]
- Shaked, L.; Weiss, D.; Reuveini, M.; Nissim, A.; Oren, M. Increased anthocyanin accumulation in aster flowers at elevated temperatures due to magnesium treatment. Physiol Plant. 2002, 114, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, D.; Zhou, L.; Zhang, X.; Liao, J.; Duan, Y.; Wen, B.; Ma, Y.; Wang, Y.; Fang, W.; et al. Transcriptomic and metabolomic profiling of Camellia sinensis L. cv. ‘Suchazao’ exposed to temperature stresses reveals modification in protein synthesis and photosynthetic and anthocyanin biosynthetic pathways. Tree Physiol. 2019, 1, 1583–1599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, S.; Shi, L.; Gong, D.; Zhang, S.; Zhao, Q.; Zhan, D.; Vasseur, L.; Wang, Y.; Yu, J.; et al. Haplotype-resolved genome assembly provides insights into evolutionary history of the tea plant Camellia sinensis. Nat Genet. 2021, 53, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, U.; Valentine, W.; Christie, J.; Hays, J.; Jenkins, G.; Weisshaar, B. Identification of UV/blue light-response elements in the Arabidopsis thaliana chalcone synthase promoter using a homologous protoplast transient expression system. Plant Mol. Biol. 1998, 36, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, U.; Sagasser, M.; Mehrtens, F.; Stracke, R.; Weisshaar, B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol. Biol. 2005, 57, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Zoratti, L.; Karppinen, K.; Escobar, A.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Ma, H.; Zhang, Y.; Zhang, J.; Wu, T.; Song, T.; Yao, Y.; Tian, J. Apple MPK4 mediates phosphorylation of MYB1 to enhance light-induced anthocyanin accumulation. Plant J. 2021, 106, 1728–1745. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads | Clean Base (Gb) | Q30 (%) | GC Content (%) | Reads Mapped (%) | Unique Mapped (%) |
|---|---|---|---|---|---|---|
| HL-1 | 43,832,718 | 6.57 | 90.65 | 42.81 | 86.34 | 81.33 |
| HL-2 | 47,538,358 | 7.13 | 90.15 | 43.8 | 86.61 | 81.93 |
| HL-3 | 43,692,182 | 6.55 | 89.87 | 42.91 | 86.24 | 81.56 |
| HS-1 | 44,636,012 | 6.7 | 90.2 | 43.65 | 86.37 | 81.65 |
| HS-2 | 46,091,272 | 6.91 | 91.56 | 43.94 | 87.78 | 82.66 |
| HS-3 | 44,820,114 | 6.72 | 90.49 | 43.41 | 86.32 | 81.33 |
| LL-1 | 44,589,648 | 6.69 | 90.77 | 43.43 | 87.62 | 82.52 |
| LL-2 | 45,562,798 | 6.83 | 90.51 | 43.4 | 87.13 | 82.24 |
| LL-3 | 44,766,816 | 6.72 | 91.2 | 43.16 | 86.96 | 81.86 |
| LS-1 | 44,646,814 | 6.7 | 90.92 | 43.37 | 87.29 | 82.37 |
| LS-2 | 45,546,030 | 6.83 | 90.51 | 42.76 | 85.85 | 80.94 |
| LS-3 | 45,172,040 | 6.78 | 89.98 | 42.87 | 86.35 | 81.63 |
| Gene ID | Gene Name | Description | FPKM Expression Mean | |||
|---|---|---|---|---|---|---|
| HS | HL | LS | LL | |||
| TGY116873 | ANSa | Anthocyanin synthase | 133.58 | 134.32 | 87.19 | 194.49 |
| TGY105701 | ANSb | Anthocyanin synthase | 9.18 | 5.32 | 2.72 | 5.35 |
| TGY031680 | DFR | Dihydroflavonol 4-reductase | 46.36 | 73.96 | 65.53 | 138.97 |
| TGY079101 | CHS | Chalcone synthase 3 | 0.62 | 1.00 | 0.44 | 1.56 |
| TGY008324 | F3Ha | Flavanone-3-hydroxylase | 36.32 | 39.12 | 12.37 | 44.94 |
| TGY040587 | F3Hb | Flavanone-3-hydroxylase | 1.56 | 0.65 | 0.60 | 0.79 |
| TGY109805 | HCTa | shikimate hydroxycinnamoyltransferase | 1.09 | 1.20 | 0.49 | 1.29 |
| TGY025428 | HCTb | shikimate hydroxycinnamoyltransferase | 17.00 | 31.75 | 14.06 | 11.96 |
| TGY042101 | HCTc | shikimate hydroxycinnamoyltransferase | 0.29 | 0.75 | 0.06 | 0.11 |
| TGY063698 | HCTd | shikimate hydroxycinnamoyltransferase | 8.35 | 7.42 | 7.08 | 14.81 |
| TGY107898 | HCTe | shikimate hydroxycinnamoyltransferase | 3.50 | 5.34 | 2.56 | 5.86 |
| TGY032709 | HCTf | shikimate hydroxycinnamoyltransferase | 25.24 | 33.74 | 18.33 | 14.23 |
| novel.2405 | HCTg | shikimate hydroxycinnamoyltransferase | 0.60 | 0.87 | 1.41 | 1.69 |
| TGY020135 | HCTh | shikimate hydroxycinnamoyltransferase | 0.32 | 0.69 | 1.01 | 1.18 |
| TGY052439 | CCoAOMT | Caffeoyl-CoA-O-methyltransferase | 0.98 | 0.87 | 1.34 | 3.16 |
| TGY000809 | UGT29a | Flavanone 7-O-glucoside 2″-O-beta-L-rhamnosyltransferase | 1.32 | 2.48 | 2.84 | 1.42 |
| TGY000843 | UGT29b | 0.91 | 2.36 | 1.64 | 1.32 | |
| novel.2347 | UFGTa | Anthocyanidin 3-O-glucosyltransferase | 1.14 | 1.23 | 2.53 | 2.41 |
| TGY019907 | UFGTb | Anthocyanidin 3-O-glucosyltransferase | 22.61 | 24.73 | 38.84 | 51.77 |
| novel.15478 | F6H | Flavonoid 6-hydroxylase | 3.89 | 2.78 | 4.73 | 6.11 |
| TGY009888 | BAHD | Isoflavone 7-O-glucoside-6″-O-malonyltransferase | 0.66 | 1.28 | 0.39 | 0.51 |
| TGY076670 | MYB1 | Transcription factor MYB | 15.81 | 10.40 | 17.59 | 25.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Chen, W.; Tang, D.; Tan, X.; Tan, L.; Tang, Q. Metabolomic and Transcriptomic Insights into Anthocyanin Biosynthesis in ‘Ziyan’ Tea Plants under Varied Photoperiod and Temperature Conditions. Agronomy 2024, 14, 56. https://doi.org/10.3390/agronomy14010056
Yang C, Chen W, Tang D, Tan X, Tan L, Tang Q. Metabolomic and Transcriptomic Insights into Anthocyanin Biosynthesis in ‘Ziyan’ Tea Plants under Varied Photoperiod and Temperature Conditions. Agronomy. 2024; 14(1):56. https://doi.org/10.3390/agronomy14010056
Chicago/Turabian StyleYang, Chunjing, Wei Chen, Dandan Tang, Xiaoqin Tan, Liqiang Tan, and Qian Tang. 2024. "Metabolomic and Transcriptomic Insights into Anthocyanin Biosynthesis in ‘Ziyan’ Tea Plants under Varied Photoperiod and Temperature Conditions" Agronomy 14, no. 1: 56. https://doi.org/10.3390/agronomy14010056
APA StyleYang, C., Chen, W., Tang, D., Tan, X., Tan, L., & Tang, Q. (2024). Metabolomic and Transcriptomic Insights into Anthocyanin Biosynthesis in ‘Ziyan’ Tea Plants under Varied Photoperiod and Temperature Conditions. Agronomy, 14(1), 56. https://doi.org/10.3390/agronomy14010056