
Microbiota Ecosystem Services in Vineyards and Wine: A Review

 ,
,  ,
,  and
and 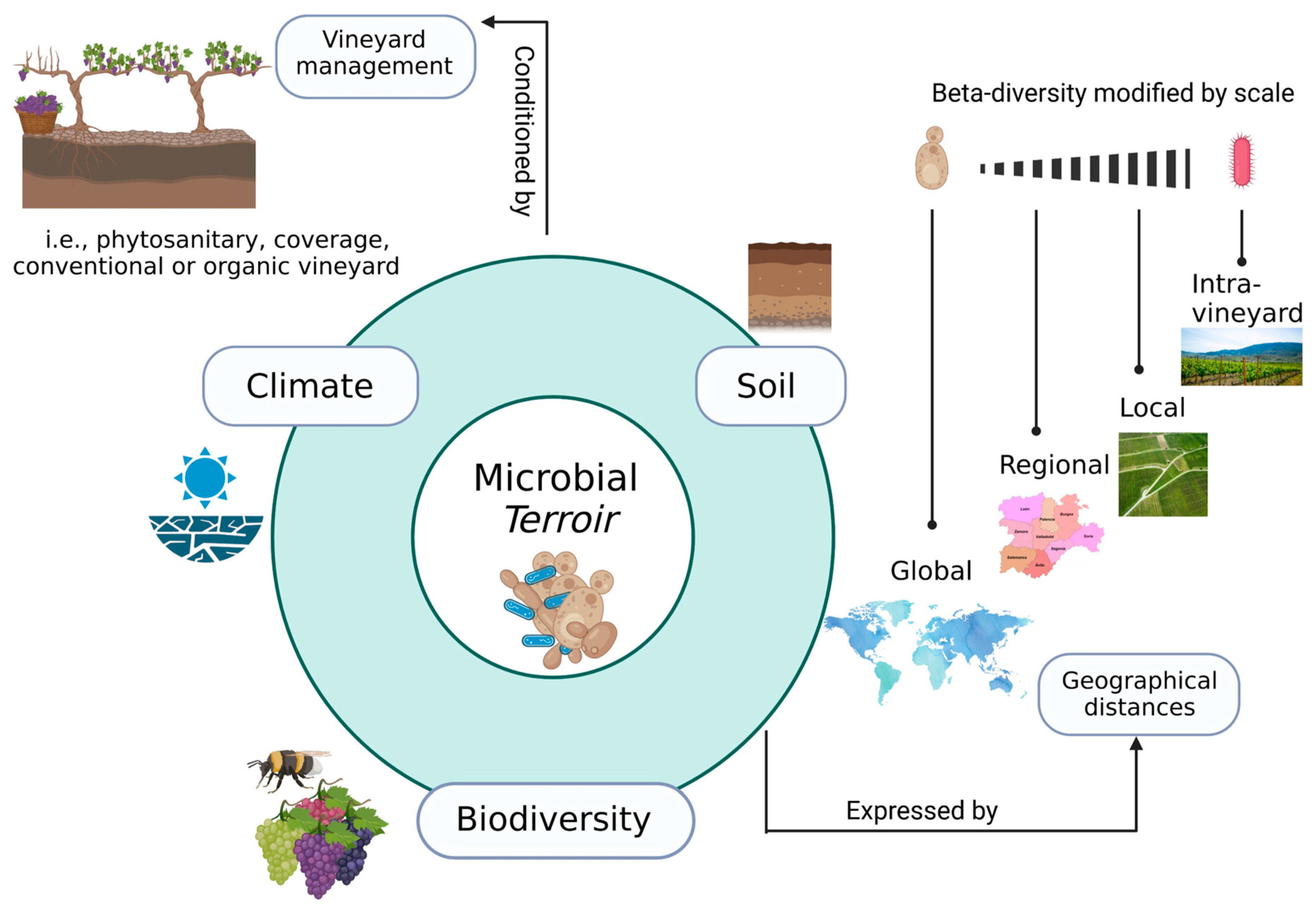{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Microbiota and Wine: The Evolution of Our Scientific Knowledge
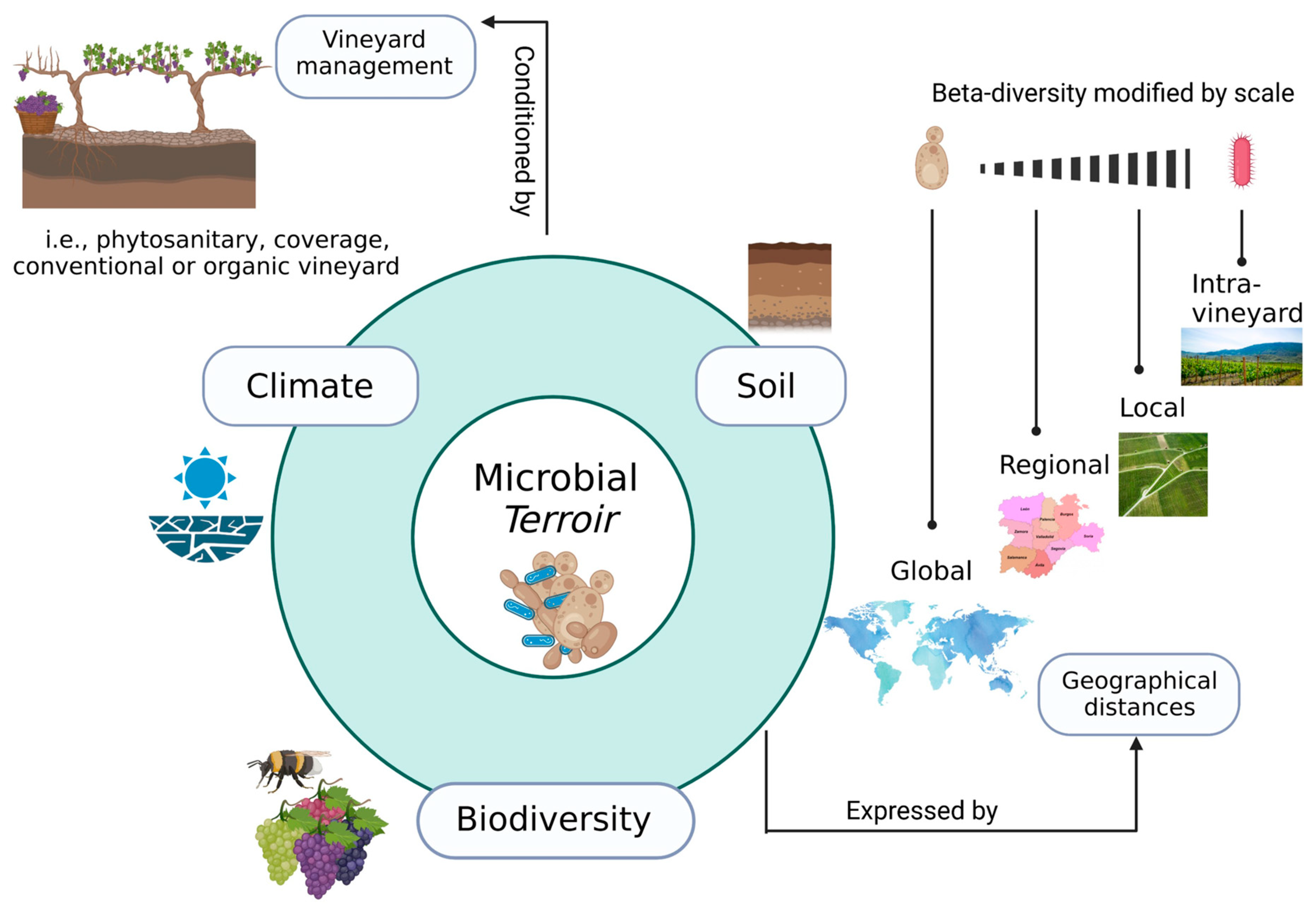
3. Terroir and Microbiota of Wine
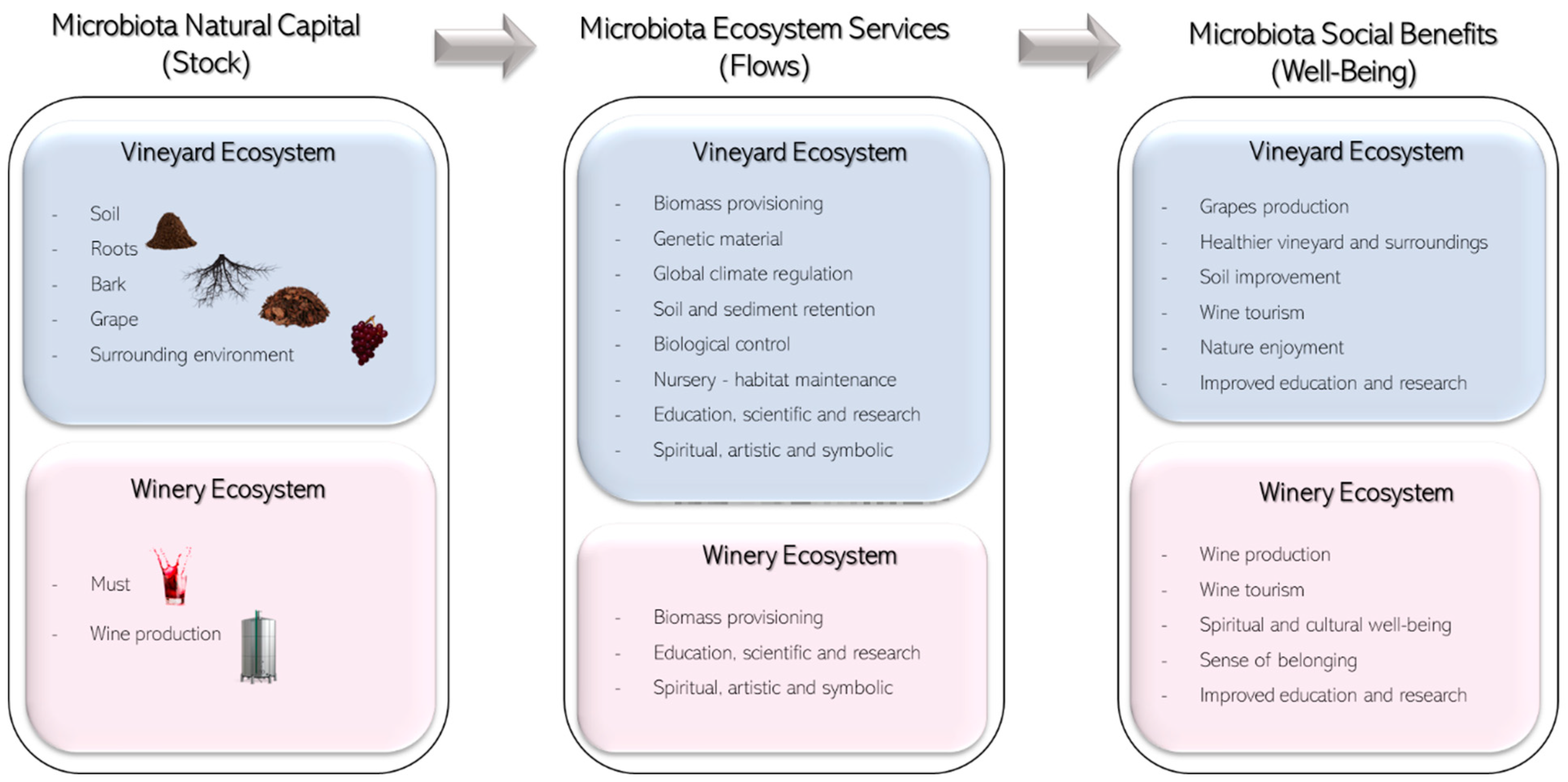
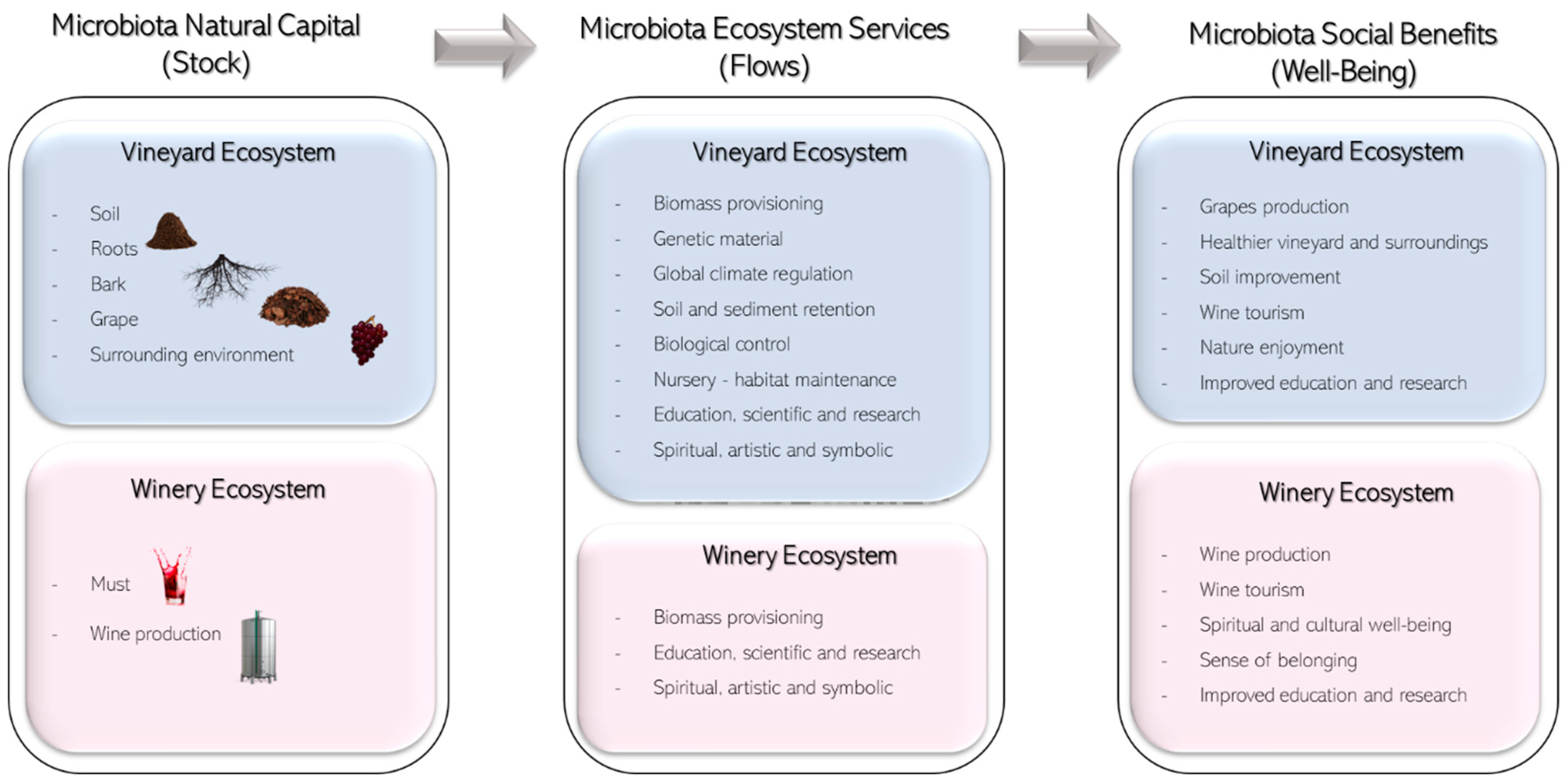
4. Ecosystem Services Provided by Microbiota in Vineyards and Wines
4.1. Provisioning Ecosystem Services
4.1.1. Biomass (Crop) Provisioning Services
4.1.2. Genetic Material Services
4.2. Regulating Ecosystem Services
4.2.1. Carbon Storage
4.2.2. Soil Quality Regulation Services
4.2.3. Soil and Sediment Retention Services
4.2.4. Biological Control Services
4.2.5. Nursery Population and Habitat Maintenance Services
4.3. Cultural Ecosystem Services
4.3.1. Recreation-Related Services
4.3.2. Visual Amenity Services
4.3.3. Education, Scientific, and Research Services
4.3.4. Spiritual, Artistic, and Symbolic Services
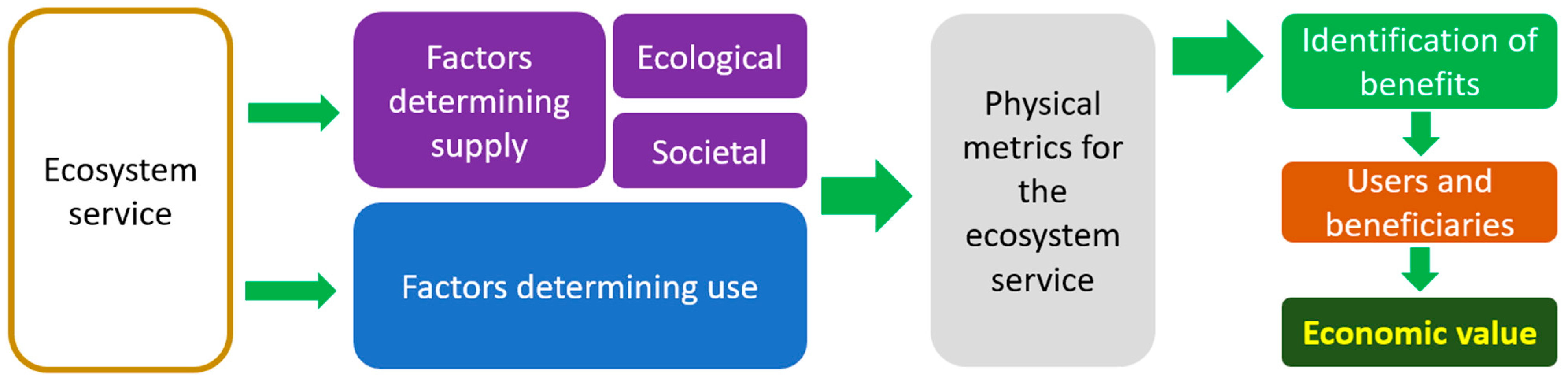
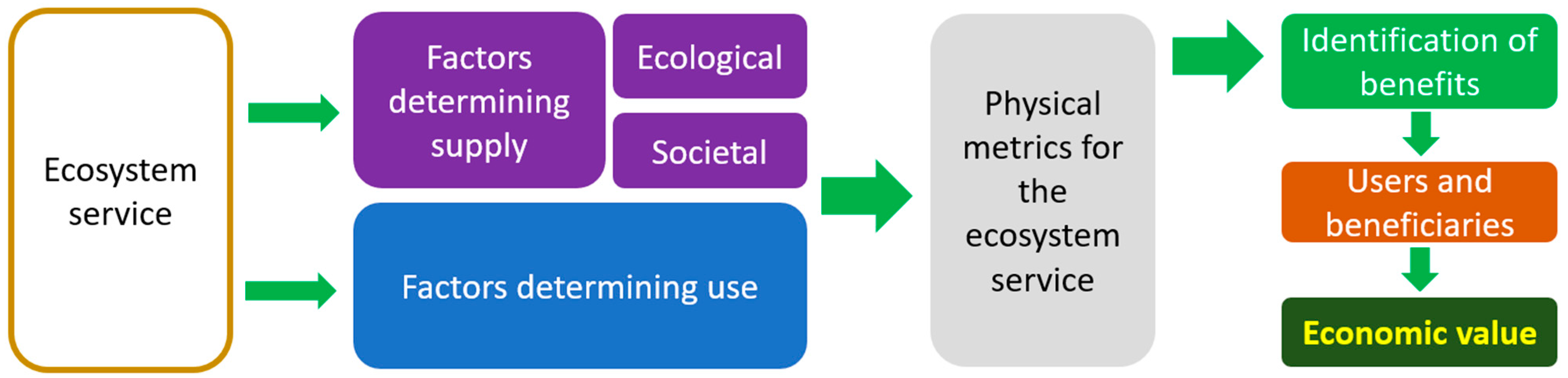
5. Economic Valuation of Microbiota Ecosystem Services in Vineyards and Wine
6. Vineyard Management and Microbiota Ecosystem Services
7. Towards Smart Farming: Microbiota as a Nature-Based Solution in Vineyards and Wineries
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPBES. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Díaz, S., Settele, J., Brondízio, E.S., Ngo, H.T., Guèze, M., Agard, J., Arneth, A., Balvanera, P., Brauman, K.A., Butchart, S.H.M., et al., Eds.; IPBES Secretariat: Bonn, Germany, 2019; 56p. [Google Scholar]
- Fu, B.; Wang, S.; Su, C.; Forsius, M. Linking ecosystem processes and ecosystem services. Curr. Opin. Environ. Sustain. 2013, 5, 4–10. [Google Scholar] [CrossRef]
- Daily, G.C. Nature’s Services. Societal Dependence on Natural Ecosystems; Island Press: Washington, DC, USA, 1997; 392p, ISBN 1-55963-475-8. [Google Scholar]
- Haines-Young, R.; Potschin, M.B. Common International Classification of Ecosystem Services (CICES) v5.1 and Guidance on the Application of the Revised Structure. 2018. Available online: www.cices.eu (accessed on 14 January 2023).
- United Nations. System of Environmental-Economic Accounting—Ecosystem Accounting (SEEA EA). 2021. Available online: https://seea.un.org/ecosystem-accounting (accessed on 15 January 2023).
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, E.; Grasso, M.; Hannon, B.; Limburgo, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Ali, G.; Dahlhaus, P. Roles of Selective Agriculture Practices in sustainable agriculture performance: A systematic review. Sustainability 2022, 14, 3185. [Google Scholar] [CrossRef]
- Robertson, G.P.; Swinton, S.M. Reconciling agricultural productivity and environmental integrity: A grand challenge for agriculture. Front. Ecol. Environ. 2005, 3, 38–46. [Google Scholar] [CrossRef]
- Huang, J.; Tichit, M.; Poulot, M.; Darly, S.; Li, S.; Petit, C.; Aubry, C. Comparative review of multifunctionality and ecosystem services in sustainable agriculture. J. Environ. Manag. 2015, 149, 138–147. [Google Scholar] [CrossRef]
- TEEB. TEEB for Agriculture & Food: Scientific and Economic Foundations; UN Environment: Geneva, Switzerland, 2018. [Google Scholar]
- OIV-International Organisation of Vine and Wine. State of the World Vine and Wine Sector 2021; OIV-International Organisation of Vine and Wine: Dijon, France, 2022; 20p. [Google Scholar]
- Bindi, M.; Nunes, P.A.L.D. Vineyards and vineyard management related to ecosystem services: Experiences from a wide range of enological regions in the context of global climate change. J. Wine Econ. 2016, 11, 66–68. [Google Scholar] [CrossRef]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 2020, 706, 135839. [Google Scholar] [CrossRef]
- Winkler, K.J.; Viers, J.H.; Nicholas, K.A. Assessing ecosystem services and multifunctionality for vineyard systems. Front. Environ. Sci. 2017, 5, 15. [Google Scholar] [CrossRef]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef]
- Garcia, L.; Celette, F.; Gary, C.; Ripoche, A.; Valdes-Gomez, H.; Metay, A. Management of service crops for the provision of ecosystem services in vineyards: A review. Agric. Ecosyst. Environ. 2018, 251, 158–170. [Google Scholar] [CrossRef]
- Candiago, S.; Winkler, K.J.; Giombini, V.; Giupponi, C.; Vigl, L.E. An ecosystem service approach to the study of vineyard landscapes in the context of climate change: A review. Sustain. Sci. 2023, 18, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Han, K.Y.; Kröger, L.; Buchholz, F.; Dewan, I.; Quaas, M.; Schulenbur, H.; Reusch, T.B.H. The economics of microbiodiversity. Ecol. Econ. 2023, 204, 107664. [Google Scholar] [CrossRef]
- Dong, Y.; Duan, S.; Xia, Q.; Liang, Z.; Dong, X.; Margaryan, K.; Musayev, M.; Goryslavets, S.; Zdunić, G.; Bert, P.F.; et al. Dual domestications and origin of traits in grapevine evolution. Science 2023, 379, 892–901. [Google Scholar] [CrossRef] [PubMed]
- McGovern, P.E.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Zarraonaindia, I.; Perisin, M.; Palacios, A.; Acedo, A. From vineyard soil to wine fermentation: Microbiome approximations to explain the “terroir” concept. Front. Microbiol. 2017, 8, 821. [Google Scholar] [CrossRef] [PubMed]
- Guasch Jané, M.R.; Fonseca, S.; Ibrahim, M. “IREP EN KEMET” Wine of ancient Egypt: Documenting the viticulture and winemaking scenes in the Egyptian tombs. ISPRS J. Photogramm. Remote Sens. 2013, 2, 157–161. [Google Scholar] [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, H. Chapter 1: The history of Chinese wine. In Overview of Wine in China; EDP Sciences: Les Ulis, France, 2022; pp. 1–8. [Google Scholar] [CrossRef]
- Stevenson, A.C. Studies in the vegetational history of S. W. Spain II: Palynological investigations at Laguna de Las Madres, S.W. Spain. J. Biogeogr. 1985, 12, 293–314. [Google Scholar] [CrossRef]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef]
- Cavalieri, D.; McGovern, P.E.; Hartl, D.L.; Mortimer, R.; Polsinelli, M. Evidence for S. cerevisiae fermentation in ancient wine. J. Mol. Evol. 2003, 57, 226–232. [Google Scholar] [CrossRef]
- da Silva Fernandes, F.; de Souza, E.S.; Carneiro, L.M.; Alves Silva, J.P.; de Souza, J.V.B.; da Silva Batista, J. Current ethanol production requirements for the yeast Saccharomyces cerevisiae. Int. J. Microbiol. 2022, 2022, 7878830. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, L. Etudes sur le vin. Imprimeurs Imperials; BNF: Paris, France, 1866. [Google Scholar]
- Carrau, F.; Gaggero, C.; Aguilar, P.S. Yeast diversity and native vigor for flavor phenotypes. Trends Biotechnol. 2015, 33, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.M.; Legout, S. Louis Pasteur: Between myth and reality. Biomolecules 2022, 12, 596. [Google Scholar] [CrossRef] [PubMed]
- Marsit, S.; Dequin, S. Diversity and adaptative evolution of Saccharomyces wine yeast: A review. FEMS Yeast Res. 2015, 15, fov067. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderón, F.; Benito, S. Outlining the influence of non-conventional yeasts in wine ageing over lees. Yeast 2016, 33, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Englezos, V.; Jovial, N.P.; Gianvito, P.D.; Rantsiou, K.; Cocolín, L. Microbial interactions in winemaking: Ecological aspects and effect on wine quality. Trends Food Sci. Technol. 2022, 127, 99–113. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Bauer, F.F.; Pretorius, I.S. Yeast stress response and fermentation efficiency: How to survive the making of wine—A review. S. Afr. J. Enol. Vitic. 2000, 21, 27–51. [Google Scholar] [CrossRef]
- Querol, A.; Pérez-Torrado, R.; Alonso-Del-Real, J.; Minebois, R.; Stribny, J.; Oliveira, B.M.; Barrio, E. New trends in the uses of yeasts in oenology. Adv. Food Nutr. Res. 2018, 85, 177–210. [Google Scholar] [CrossRef]
- Ortiz-Álvarez, R.; Ortega-Arranz, H.; Ontiveros, V.J.; Celis, M.; Ravarani, C.; Acedo, A.; Belda, I. Network properties of local fungal communities reveal the anthropogenic disturbance consequences of farming practices in vineyard soils. Msystems 2021, 6, e00344-21. [Google Scholar] [CrossRef]
- Gonzalez, R.; Morales, P. Truth in wine yeast. Microb. Biotechnol. 2022, 15, 1339–1356. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.Y.; Han, D.Y.; Duan, S.F.; Wang, Q.M. The ecology and evolution of the baker’s yeast Saccharomyces cerevisiae. Genes 2022, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torrado, R.; Barrio, E.; Querol, A. Alternative yeasts for winemaking: Saccharomyces non-cerevisiae and its hybrids. Crit. Rev. Food Sci. Nutr. 2017, 58, 1780–1790. [Google Scholar] [CrossRef]
- Sidari, R.; Ženišová, K.; Tobolková, B.; Belajová, E.; Cabicarová, T.; Bučková, M.; Puškárová, A.; Planý, M.; Kuchta, T.; Pangallo, D. Wine yeasts selection: Laboratory characterization and protocol review. Microorganisms 2021, 9, 2223. [Google Scholar] [CrossRef] [PubMed]
- Mateo, J.J.; Maicas, S. Application of non-Saccharomyces yeasts to wine-making process. Fermentation 2016, 2, 14. [Google Scholar] [CrossRef]
- Chalvantzi, I.; Banilas, G.; Tassou, C.; Nisiotou, A. Patterns of genetic diversity and the invasion of commercial starters in Saccharomyces cerevisiae vineyard populations of Santorini Island. Foods 2020, 9, 561. [Google Scholar] [CrossRef]
- Engel, S.R.; Dietrich, F.S.; Fisk, D.G.; Binkley, G.; Balakrishnan, R.; Costanzo, M.C.; Dwight, S.S.; Hitz, B.C.; Karra, K.; Nash, R.S.; et al. The reference genome sequence of Saccharomyces cerevisiae: Then and now. G3 Genes Genomes Genet. 2014, 4, 389–398. [Google Scholar] [CrossRef]
- Belda, I.; Gobbi, A.; Ruiz, J.; de Celis, M.; Ortiz-Álvarez, R.; Acedo, A.; Santos, A. Microbiomics to define wine terroir. In Comprehensive Foodomics; Cifuentes, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Belda, I.; Palacios, A.; Fresno, J.; Ortega, H.; Acedo, A. WineSeq: A new tool for the study of the functional biodiversity of soils, and its use as a biomarker and guide for vitiviniculture practices. BIO Web Conf. 2017, 9, 01012. [Google Scholar] [CrossRef]
- Li, R.; Yang, S.; Lin, M.; Guo, S.; Han, X.; Ren, M.; Du, L.; Song, Y.; You, Y.; Zhan, J.; et al. The biogeography of fungal communities across different Chinese wine-producing regions associated with environmental factors and spontaneous fermentation performance. Front. Microbiol. 2022, 12, 636639. [Google Scholar] [CrossRef]
- Portillo, M.C.; Mas, A. Analysis of microbial diversity and dynamics during wine fermentation of Grenache grape variety by high-throughput barcoding sequencing. LWT Food Sci. Technol. 2016, 72, 317–321. [Google Scholar] [CrossRef]
- Stefanini, I.; Albanese, D.; Cavazza, A.; Franciosi, E.; De Filippo, C.; Donati, C.; Cavalieri, D. Dynamic changes in microbiota and mycobiota during spontaneous ‘Vino Santo Trentino’ fermentation. Microb. Biotechnol. 2016, 9, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Sirén, K.; Mak, S.S.T.; Fischer, U.; Hansen, L.H.; Gilbert, M.T.P. Multi-omics and potential applications in wine production. Curr. Opin. Biotechnol. 2019, 56, 172–178. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Joseph, C.L.; Allen, G.; Benson, A.K.; Mills, D.A. Next-generation sequencing reveals significant bacterial diversity of botrytized wine. PLoS ONE 2012, 7, e36357. [Google Scholar] [CrossRef]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [PubMed]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, B.; Freeborough, M.H.; Maree, H.J.; Celton, J.M.; Rees, D.J.G.; Burger, J.T. Deep sequencing analysis of viruses infecting grapevines: Virome of a vineyard. Virology 2010, 400, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Setati, M.E.; Jacobson, D.; Bauer, F.F. Sequence-based analysis of the Vitis vinifera L. cv Cabernet sauvignon grape must mycobiome in three South African Vineyards employing distinct agronomic systems. Front. Microbiol. 2015, 6, 1358. [Google Scholar] [CrossRef]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S.; et al. The soil microbiome influences grapevine-associated microbiota. mBio 2015, 6, e02527-14. [Google Scholar] [CrossRef]
- Cobos, R.; Ibáñez, A.; Diez-Galán, A.; Calvo-Peña, C.; Ghoreshizadeh, S.; Coque, J.J.R. The grapevine microbiome to the rescue: Implications for the biocontrol of trunk diseases. Plants 2022, 11, 840. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Nityagovsky, N.N.; Suprun, A.R.; Ananev, A.A.; Dubrovina, A.S.; Kiselev, K.V. The diversity of fungal endophytes from wild grape Vitis amurensis Rupr. Plants 2022, 11, 2897. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.; Albanese, D.; Stegen, J.; Franceschi, P.; Coller, E.; Zanzotti, R.; Ioriatti, C.; Stefani, E.; Pindo, M.; Cestaro, A.; et al. Distinct and temporally stable assembly mechanisms shape bacterial and fungal communities in vineyard soils. Microb. Ecol. 2023, 86, 337–349. [Google Scholar] [CrossRef]
- Fabres, P.J.; Collins, C.; Cavagnaro, T.R.; Rodríguez-López, C.M. A concise review on multi-omics data integration for terroir analysis in Vitis vinifera. Front. Plant Sci. 2017, 8, 1065. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, A.; Acedo, A.; Imam, N.; Santini, R.G.; Ortiz-Álvarez, R.; Ellegaard-Jensen, L.; Belda, I.; Hansen, L.H. A global microbiome survey of vineyard soils highlights the microbial dimension of viticultural terroirs. Commun. Biol. 2022, 5, 241. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Afonso, I.M.; Pereira, J.; Rocha, R.; Rodrigues, A.S. Epiphitic microbiome of Alvarinho wine grapes from different geographic regions in Portugal. Biology 2023, 12, 146. [Google Scholar] [CrossRef]
- Chiarello, M.; McCauley, M.; Villéger, S.; Jackson, C.R. Ranking the biases: The choice of OTUs vs. ASVs in 16S rRNA amplicon data analysis has stronger effects on diversity measures than rarefaction and OUT identity threshold. PLoS ONE 2022, 17, e0264443. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- García-López, R.; Cornejo-Granados, F.; Lopez-Zavala, A.A.; Cota-Huízar, A.; Sotelo-Mundo, R.; Gómez-Gil, B.; Ochoa-Leyva, A. OTUs and ASVs produce comparable taxonomic and diversity from shrimp microbiota 16S profiles using tailored abundance filters. Genes 2021, 12, 564. [Google Scholar] [CrossRef]
- Glassman, S.I.; Martiny, J.B.H. Broadscale ecological patterns are robust to use of exact sequence variants versus Operational Taxonomic Units. mSphere 2018, 3, e00148-18. [Google Scholar] [CrossRef]
- Venter, J.C.; Glass, J.I.; Hutchison, C.A.; Vashee, S. Synthetic chromosomes, genomes, viruses and cells. Cell 2022, 185, 2708–2724. [Google Scholar] [CrossRef]
- Pretorius, I.S. Synthetic genome engineering forging new frontiers for wine yeast. Crit. Rev. Biotechol. 2017, 37, 112–136. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S.; Boeke, J.D. Yeast 2.0-connecting the dots in the construction of the world’s first functional synthetic eukaryotic genome. FEMS Yeast Res. 2018, 18, foy032. [Google Scholar] [CrossRef] [PubMed]
- Kutyna, D.R.; Onetto, C.A.; Williams, T.C.; Goold, H.D.; Paulsen, I.T.; Pretorius, I.S.; Johnson, D.L.; Borneman, A.R. Construction of a synthetic Saccharomyces cerevisiae pan-genome neo-chromosome. Nat. Commun. 2022, 13, 3628. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S. Tasting the terroir of wine yeast innovation. FEMS Yeast Res. 2020, 20, foz084. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S. Visualizing the new frontiers in wine yeast research. FEMS Yeast Res. 2022, 22, foac010. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, P.; Chen, D.; Howell, K. From the vineyard to the winery: How microbial ecology drives regional distinctiveness of wine. Front. Microbiol. 2019, 10, 2679. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Pio Rossetti, A.; Battistelli, N.; Zulli, C.; Cichelli, A.; Arfelli, A.; Arfelli, G.; Tofalo, R. Impact of vineyard management on grape fungal community and Montepulciano d’Abruzzo wine quality. Food Res. Int. 2022, 158, 111577. [Google Scholar] [CrossRef] [PubMed]
- Griggs, R.G.; Steenwerth, K.L.; Mills, D.A.; Cantu, D.; Bokulich, N.A. Sources and assembly of microbial communities in vineyards as a functional component of winegrowing. Front. Microbiol. 2021, 12, 673810. [Google Scholar] [CrossRef]
- Giffard, B.; Winter, S.; Guidoni, S.; Nicolai, A.; Castaldini, M.; Cluzeau, D.; Coll, P.; Cortet, J.; Le Cadre, E.; d’Errico, G.; et al. Vineyard management and its impacts on soil biodiversity, functions, and ecosystem services. Front. Ecol. Evol. 2022, 10, 850272. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Seguin, G. The concept of Terroir in viticulture. J. Wine Res. 2006, 17, 1–10. [Google Scholar] [CrossRef]
- Lewin, B. Wine Myths and Reality; Wine Appreciation Guild: San Francisco, CA, USA, 2010; 636p, ISBN 1934259519. [Google Scholar]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial terroir for wine grapes. Proc. Natl. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Lauga, B.; Miot-Sertier, C.; Mercier, A.; Lonvaud, A.; Soulas, M.L.; Soulas, G.; Masneuf-Pomarède, I. Characterization of epiphytic bacterial communities from grapes, leaves, bark and soil of grapevine plants grown, and their relations. PLoS ONE 2013, 8, e73013. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Silva, A.; Laureano, G.; Pereira, M.; Días, R.; Moreira da Silva, J.; Oliveira, N.; Gouveia, C.; Cruz, C.; Gama-Carvalho, M.; Alagna, F.; et al. A new perspective for vineyard terroir identity: Looking for microbial indicator species by long read nanopore sequencing. Microorganisms 2023, 11, 672. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage and climate. Proc. Nail. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [PubMed]
- Rivas, G.A.; Guillade, A.C.; Semorile, L.C.; Delfederico, L. Influence of climate on soil and wine bacterial diversity on a vineyard in a non-traditional wine region in Argentina. Front. Microbiol. 2021, 12, 726384. [Google Scholar] [CrossRef]
- Liu, D.; Chen, Q.; Zhang, P.; Chen, D.; Howell, K.S. The fungal microbiome is an important component of vineyard ecosystems and correlates with regional distinctiveness of wine. mSphere 2020, 5, e00534-20. [Google Scholar] [CrossRef]
- Burns, K.N.; Kluepfel, D.A.; Strauss, S.L.; Bokulich, N.A.; Cantu, D.; Steenwerth, K.L. Vineyard soil bacterial diversity and composition revealed by 16S rRNA genes: Differentiation by geographic features. Soil Biol. Biochem. 2015, 91, 232–247. [Google Scholar] [CrossRef]
- Vadour, E.; Costantini, E.; Jones, G.V.; Mocali, S. An overview of the recent approaches for terroir functional modelling, footprinting and zoning. Soil Discuss. 2015, 1, 827–906. [Google Scholar] [CrossRef]
- Gayevski, V.; Goddard, M.R. Geographic delineations of yeast communities and populations associated with vines and wines in New Zealand. ISME J. 2012, 6, 1281–1290. [Google Scholar] [CrossRef]
- Mezzasalma, V.; Sandionigi, A.; Guzzetti, L.; Galimberti, A.; Grando, M.S.; Tardaguila, J.; Labra, M. Geographical and cultivar features differentiate grape microbiota in Northern Italy and Spain vineyards. Front. Microbiol. 2018, 9, 946. [Google Scholar] [CrossRef] [PubMed]
- Tronchoni, J.; Setati, M.E.; Fracassetti, D.; Valdetara, F.; Maghradze, D.; Foschino, R.; Curiel, J.A.; Morales, P.; Gonzalez, R.; Vigentini, I.; et al. Identifying the main drivers in microbial diversity for cabernet sauvignon cultivars from Europe to South Africa: Evidence for a cultivar-specific microbial fingerprint. J. Fungi 2022, 8, 1034. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Tsai, P.; Anfang, N.; Ross, H.A.; Goddard, M.R. Pyrosequencing reveals regional differences in fruit-associated fungal communities. Environ. Microbiol. 2014, 16, 2848–2858. [Google Scholar] [CrossRef] [PubMed]
- Morrison-Whittle, P.; Goddard, M.R. From vineyard to winery: A source map of microbial diversity driving wine fermentation. Environ. Microbiol. 2018, 20, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kioroglou, D.; Kraeva-Deloire, E.; Leigh, M.; Schmidtke, L.M.; Mas, A.; Portillo, M.C. Geographical origin has a greater impact on grape berry fungal community than grape variety and maturation state. Microorganisms 2019, 7, 669. [Google Scholar] [CrossRef] [PubMed]
- Kamilari, E.; Mina, M.; Karallis, C.; Tsaltas, D. Metataxonomic analysis of grape microbiota during wine fermentation reveals the distinction of Cyprus regional terroirs. Front. Microbiol. 2021, 12, 726483. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.F.; Migicovsky, Z.; Trello, G.E.; Miller, A.J. Grapevine bacterial communities display compartment-specific dynamics over space and time within the Central Valley of California. Environ. Microbiome 2023, 18, 84. [Google Scholar] [CrossRef] [PubMed]
- Portillo, M.C.; Franquès, J.; Araque, I.; Reguant, C.; Bordons, A. Bacterial diversity of grenache and carignan grape surface from different vineyards at Priorat wine region (Catalonia, Spain). Int. J. Food Microbiol. 2016, 219, 56–63. [Google Scholar] [CrossRef]
- Knight, S.J.; Karon, O.; Goddard, M.R. Small scale fungal community differentiation in a vineyard system. Food Microbiol. 2020, 87, 103358. [Google Scholar] [CrossRef]
- Yan, H.; Ge, C.; Zhou, J.; Li, J. Diversity of soil fungi in the vineyards of Changli región in China. Can. J. Microbiol. 2022, 68, 341–352. [Google Scholar] [CrossRef]
- Setati, M.E.; Jacobson, D.; Andong, U.; Bauer, F. The vineyard yeast microbiome, a mixed model microbial map. PLoS ONE 2012, 7, e52609. [Google Scholar] [CrossRef]
- Chalvantzi, I.; Banilas, G.; Tassou, C.; Nisiotou, A. Biogeographical regionalization of wine yeast communities in Greece and environmental drivers of species distribution at a local scale. Front. Microbiol. 2021, 12, 705001. [Google Scholar] [CrossRef]
- Zhou, J.; Cavagnaro, T.R.; De Bei, R.; Nelson, T.M.; Stephen, J.R.; Metcalfe, A.; Gilliham, M.; Breen, J.; Collins, C.; Rodríguez López, C.M. Wine terroir and the soil bacteria: An amplicon sequencing-based assessment of the Barossa Valley and its sub-Regions. Front. Microbiol. 2021, 11, 597944. [Google Scholar] [CrossRef]
- Wei, R.T.; Chen, N.; Ding, Y.T.; Wang, L.; Gao, F.F.; Zhang, L.; Liu, Y.H.; Li, H.; Wang, H. Diversity and dynamics of epidermal microbes during grape development of cabernet sauvignon (Vitis vinifera L.) in the ecological viticulture model in Wuhai, China. Front. Microbiol. 2022, 13, 935647. [Google Scholar] [CrossRef]
- Kecskeméti, E.; Berkelmann-Löhnertz, B.; Reineke, A. Are epiphytic microbial communities in the carposphere of ripening grape clusters (Vitis vinifera L.) different between conventional, organic, and biodynamic grapes? PLoS ONE 2016, 11, e0160852. [Google Scholar] [CrossRef]
- Grangeteau, C.; Roullier-Gall, C.; Rousseaux, S.; Gougeon, R.D.; Schmitt-Kopplin, P.; Alexandre, H.; Guilloux-Benatier, M. Wine microbiology is driven by vineyard and winery anthropogenic factors. Microb. Biotechnol. 2017, 10, 354–370. [Google Scholar] [CrossRef]
- Morrison-Whittle, P.; Lee, S.A.; Goddard, M.R. Fungal communities are differentially affected by conventional and biodynamic agricultural management approaches in vineyard ecosystems. Agric. Ecosyst. Environ. 2017, 246, 306–313. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Chen, X.; Cui, B.; Bai, Z.; Zhuang, G. Variety features differentiate microbiota in the grape leaves. Can. J. Microbiol. 2020, 66, 653–663. [Google Scholar] [CrossRef]
- Martins, G.; Casini, C.; Da Costa, J.P.; Geny, L.; Lonvaud, A.; Masneuf-Pomarède, I. Correlation between water activity (aw) and microbial epiphytic communities associated with grape berries. OENO One 2020, 54, 49–61. [Google Scholar] [CrossRef]
- Mozzachiodi, S.; Bai, F.Y.; Baldrian, P.; Bell, G.; Boundy-Mills, K.; Buzzini, P.; Čadež, N.; Cubillos, F.A.; Dashko, S.; Dimitrov, R.; et al. Yeasts from temperate forests. Yeast 2022, 39, 4–24. [Google Scholar] [CrossRef]
- Valentini, B.; Barbero, F.; Casacci, L.P.; Luganini, A.; Stefanini, I. Forest influence yeast populations vectored by insects into vineyards. Front. Microbiol. 2022, 13, 1039939. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Sánchez, R.; Castañeda, L.E.; Godoy, K.; Barbosa, O. Is microbial terroir related to geographic distance between vineyards? Environ. Microbiol. Rep. 2017, 9, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, J.; Franco, L.M.; Cools, T.L.; De Meester, L.; Michiels, J.; Wenseleers, T.; Hassan, B.A.; Yaksi, E.; Verstrepen, K.J. The fungal aroma gene ATF1 promotes dispersal of yeast cells through insect vectors. Cell Rep. 2014, 9, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Andreolli, M.; Lampis, S.; Vallini, G. Diversity, distribution and functional role of bacterial endophytes in Vitis vinifera. In Endophytes: Biology and Biotechnology; Maheshwari, D.K., Ed.; Springer: Cham, Switzerland, 2017; Volume 5, pp. 233–266. [Google Scholar] [CrossRef]
- Martínez-Diz, M.P.; Andrés-Sodupe, M.; Bujanda, R.; Díaz-Losada, E.; Eichmeier, A.; Gramaje, D. Soil-plant compartments affect fungal microbiome diversity and composition in grapevine. Fungal Ecol. 2019, 41, 234–244. [Google Scholar] [CrossRef]
- Bao, L.; Sun, B.; Wei, Y.; Xu, N.; Zhang, S.; Gu, L.; Bai, Z. Grape cultivar features differentiate the grape rhizosphere microbiota. Plants 2022, 11, 1111. [Google Scholar] [CrossRef]
- Cureau, N.; Threlfall, R.; Marasini, D.; Lavefve, L.; Carbonero, F. Year, location, and variety impact on grape-associated mycobiota of Arkansas-grown wine grapes for wine production. Microb. Ecol. 2021, 82, 845–858. [Google Scholar] [CrossRef]
- Paolinelli, M.; Martinez, L.E.; García-Lampasona, S.; Diaz-Quirós, C.; Belmonte, M.; Ahumada, G.; Pirrone, M.A.; Farber, M.D.; Escoriaza, G.; Longone, V.; et al. Microbiome in soils of Mendoza: Microbial resources for the development of agroecological management in viticulture. OENO One 2023, 57, 191–205. [Google Scholar] [CrossRef]
- Ghiță, S.; Hnatiuc, M.; Ranca, A.; Artem, V.; Mădălina-Andreea, C. Studies on the short-term effects of the cease of pesticides use on vineyard microbiome. In Vegetation Dynamics, Changing Ecosystems and Human Responsibility; Hufnagel, L., El-Esawi, M.A., Eds.; Intechopen: London, UK, 2023; pp. 1–10. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef]
- Knight, S.J.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional microbial signatures positively correlate with differential wine phenotypes: Evidence for a microbial aspect to terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Pölme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Villarreal Ruiz, L.; Vasco-Palacios, A.M.; Quang Thu, P.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 6213. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Smets, W.; Moretti, S.; Denys, S.; Lebeer, S. Airborne bacteria in the atmosphere: Presence, purpose, and potential. Atmos. Environ. 2016, 139, 214–221. [Google Scholar] [CrossRef]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Coller, E.; Cestaro, A.; Zanzotti, R.; Bertoldi, D.; Pindo, M.; Larger, S.; Albanese, D.; Mescalchin, E.; Donati, C. Microbiome of vineyard soils is shaped by geography and management. Microbiome 2019, 7, 140. [Google Scholar] [CrossRef] [PubMed]
- Steenwerth, K.L.; Morelan, I.; Stahel, R.; Figueroa-Balderas, R.; Cantu, D.; Lee, J.; Runnebaum, R.C.; Poret-Peterson, A.T. Fungal and bacterial communities of “Pinot noir” must: Effects of vintage, growing region, climate, and basic must chemistry. PeerJ 2021, 9, e10836. [Google Scholar] [CrossRef]
- Sabate, J.; Cano, J.; Esteve-Zarzoso, B.; Guillamon, J.M. Isolation and identification of yeasts associated with vineyard and winery by RFLP analysis of ribosomal genes and mitochondrial DNA. Microbiol. Res. 2002, 157, 267–274. [Google Scholar] [CrossRef]
- Vigentini, I.; De Lorenzis, G.; Fabrizio, V.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Imazio, S.; Failla, O.; Foschino, R. The vintage effect overcomes the terroir effect: A three year survey on the wine yeast biodiversity in Franciacorta and Oltrepò Pavese, two northern Italian vine-growing areas. Microbiology 2015, 161, 362–373. [Google Scholar] [CrossRef]
- Huang, R.; Shen, L.; Yu, H.; Jiang, J.; Qin, Y.; Liu, Y.; Zhang, J.; Song, Y. Evaluation of rain-shelter cultivation mode effects on microbial diversity during Cabernet Sauvignon (Vitis vinifera L.) maturation in Jingyang, Shaanxi, China. Food Res. Int. 2022, 156, 111165. [Google Scholar] [CrossRef]
- Conacher, C.G.; Luyt, N.A.; Naidoo-Blassoples, R.K.; Rossouw, D.; Setati, M.E.; Bauer, F.F. The ecology of wine fermentation: A model for the study of complex microbial ecosystems. Appl. Microbiol. Biotechnol. 2021, 105, 3027–3043. [Google Scholar] [CrossRef]
- Alonso, A.; de Celis, M.; Ruiz, J.; Vicente, J.; Navascués, E.; Acedo, A.; Ortiz-Álvarez, R.; Belda, I.; Santos, A.; Gómez-Flechoso, M.A.; et al. Looking at the origin: Some insights into the general and fermentative microbiota of vineyard soils. Fermentation 2019, 5, 78. [Google Scholar] [CrossRef]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Ohta, M.; Richardson, P.M.; Mills, D.A. Monitoring seasonal changes in winery-resident microbiota. PLoS ONE 2013, 8, e66437. [Google Scholar] [CrossRef] [PubMed]
- Piao, H.; Hawley, E.; Kopf, S.; DeScenzo, R.; Sealock, S.; Henick-Kling, T.; Hess, M. Insights into the bacterial community and its temporal succession during the fermentation of wine grapes. Front. Microbiol. 2015, 6, 809. [Google Scholar] [CrossRef] [PubMed]
- Querol, A.; Fernández-Espinar, M.T.; del Olmo, M.; Barrio, E. Adaptative evolution of wine yeast. Int. J. Food Microbiol. 2003, 86, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Tello, J.; Cordero-Bueso, G.; Aporta, I.; Cabellos, J.M.; Arroyo, T. Genetic diversity in commercial wineries: Effects of the farming system and vinification management on wine yeasts. J. Appl. Microbiol. 2011, 112, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Fleet, G.H.; Heard, G.M. Lactic acid bacteria associated with wine grapes from several Australian vineyards. J. Appl. Microbiol. 2006, 100, 712–727. [Google Scholar] [CrossRef] [PubMed]
- Henick-Kling, T. Modification of wine flavour by malolactic fermentation. In Proceedings of the 10th International Oenological Symposium, Breisach, Germany, 3–5 May 1993; International Association for Winery Technology and Management: London, UK, 1993; pp. 290–306. [Google Scholar]
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie Leeuwenhoek 1999, 76, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 43, 111–128. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Yang, H.; Sun, L.; Xia, H.; Sun, W.; Wang, Z.; Zhang, J. Bacterial communities related to aroma formation during spontaneous fermentation of “cabernet sauvignon” wine in Ningxia, China. Foods 2022, 11, 2775. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Du, H.; Zhang, H.; Fang, C.; Jin, G.; Chen, S.; Wu, Q.; Zhang, Y.; Zhang, M.; Xu, Y. Geographically associated fungus-bacterium interactions contribute to the formation of geography-dependent flavor during high-complexity spontaneous fermentation. Microbiol. Spectr. 2022, 10, e0184422. [Google Scholar] [CrossRef] [PubMed]
- Tempère, S.; Marchal, A.; Barbe, J.C.; Bely, M.; Masneuf-Pomarede, I.; Marullo, P.; Albertin, W. The complexity of wine: Clarifying the role of microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3995–4007. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S. Nature and origin of wine quality. In Wine Tasting: A Professional Handbook, 2nd ed.; Jackson, R.S., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 387–426. [Google Scholar]
- Guillamón, J.M.; Barrio, E. Genetic polymorphism in wine yeasts: Mechanisms and methods for its detection. Front. Microbiol. 2017, 8, 806. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, S.; Yang, Q.; Han, X.; Zhou, Z.; Mao, J. Saccharomyces cerevisiae strains with low-yield higher alcohols and high-yield acetate esters improve the quality, drinking comfort and safety of huangjiu. Food Res. Int. 2022, 161, 111763. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Peng, S.; Sam, F.E.; Zhu, Y.; Liang, L.; Li, M.; Wang, J. Indigenous non-Saccharomyces yeast with β-Glucosidase activity in sequential fermentation with Saccharomyces cerevisiae: A strategy to improve the volatile composition and sensory characteristics of wines. Front. Microbiol. 2022, 13, 845837. [Google Scholar] [CrossRef]
- Amato, M.; Ballco, P.; López-Galán, B.; De Magistris, T.; Verneau, F. Exploring consumers’ perception and willingness to pay for “Non-Added Sulphite” wines through experimental auctions: A case study in Italy and Spain. Wine Econ. Policy 2017, 6, 146–154. [Google Scholar] [CrossRef]
- Carrau, F.; Henschke, P. Hanseniaspora vineae and the concept of friendly yeasts to increase autochthonous wine flavor diversity. Front. Microbiol. 2021, 12, 702093. [Google Scholar] [CrossRef]
- Scannell, D.R.; Zill, O.A.; Rokas, A.; Payen, C.; Dunham, M.J.; Eisen, M.B.; Rine, J.; Johnston, M.; Hittinger, C.T. The awesome power of yeast evolutionary genetics: New genome sequences and strain resources for the Saccharomyces sensu stricto genus. G3 Genes Genomes Genet. 2011, 1, 11–25. [Google Scholar] [CrossRef]
- Borneman, A.R.; Pretorius, I.S. Genomic insights into the Saccharomyces sensu stricto complex. Genetics 2015, 199, 281–291. [Google Scholar] [CrossRef]
- Tapia, S.M.; Pérez-Torrado, R.; Adam, A.C.; Macías, L.G.; Barrio, E.; Querol, A. Adaptative evolution in the Saccharomyces kudriavzevii Aro4p promoted a reduced production of higher alcohols. Microb. Biotechnol. 2022, 15, 2958–2969. [Google Scholar] [CrossRef]
- Karabegović, I.; Malićanin, M.; Popović, N.; Stamenković Stojanović, S.; Lazić, M.; Stanojević, J.; Danilović, B. Native non-Saccharomyces yeasts as a tool to produce distinctive and diverse Tamjanika grape wines. Foods 2022, 11, 1935. [Google Scholar] [CrossRef]
- Alsammar, H.; Delneri, D. An update on the diversity, ecology and biogeography of the Saccharomyces genus. FEMS Yeast Res. 2020, 20, foaa013. [Google Scholar] [CrossRef]
- Pontes, A.; Hutzler, M.; Brito, P.H.; Sampaio, J.P. Revisiting the taxonomic synonyms and populations of Saccharomyces cerevisiae—Phylogeny, phenotypes, ecology and domestication. Microorganisms 2020, 8, 903. [Google Scholar] [CrossRef]
- Boynton, P.J.; Greig, D. The ecology and evolution of non-domesticated Saccharomyces species. Yeast 2014, 31, 449–462. [Google Scholar] [CrossRef]
- Zahid, M.S.; Hussain, M.; Song, Y.; Li, J.; Guo, D.; Li, X.; Song, S.; Wang, L.; Xu, W.; Wang, S. Root-zone restriction regulates soil factors and bacterial community assembly of grapevine. Int. J. Mol. Sci. 2022, 23, 15628. [Google Scholar] [CrossRef]
- Pulleman, M.; Creamer, R.; Hamer, U.; Helder, J.; Pelosi, C.; Pérès, G.; Rutgers, M. Soil biodiversity, biological indicators and soil ecosystem services—An overview of European approaches. Curr. Opin. Environ. Sustain. 2012, 4, 529–538. [Google Scholar] [CrossRef]
- Burns, K.N.; Bokulich, N.A.; Cantu, D.; Greenhut, R.F.; Kluepfel, D.A.; O’Geen, A.T.; Strauss, S.L.; Steenwerth, K.L. Vineyard soil bacterial diversity and composition revealed by 16S rRNA genes: Differentiation by vineyard management. Soil Biol. Biochem. 2016, 103, 337–348. [Google Scholar] [CrossRef]
- Darriaut, R.; Tran, J.; Martins, G.; Ollat, N.; Masneuf-Pomarède, I.M.; Lauvergeat, V. In grapevine decline, microbiomes are affected differently in symptomatic and asymptomatic soils. Appl. Soil Ecol. 2023, 183, 104767. [Google Scholar] [CrossRef]
- Williams, J.N.; Morandé, J.A.; Vaghti, M.G.; Medellín-Azuara, J.; Viers, J.H. Ecosystem services in vineyard landscapes: A focus on aboveground carbon storage and accumulation. Carbon Balance Manag. 2020, 15, 23. [Google Scholar] [CrossRef]
- Song, R.; Zhu, Z.; Zhang, L.; Li, H.; Wang, H. A simple method using an allometric model to quantify the carbon sequestration capacity in vineyards. Plants 2023, 12, 997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xue, T.; Gao, F.; Wei, R.; Wang, Z.; Li, H.; Wang, H. Carbon storage distribution characteristics of vineyard ecosystems in Hongsibu, Ningxia. Plants 2021, 10, 119. [Google Scholar] [CrossRef] [PubMed]
- Sun, q.; Guoa, S.; Wang, R.; Song, J. Responses of bacterial communities and their carbon dynamics to subsoil exposure on the Loess Plateau. Sci. Total Environ. 2021, 756, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yuan, J.; Yue, F.-J.; Li, S.; Wang, B.; Mohinuzzaman, M.; Liu, Y.; Senesi, N.; Lao, X.; Li, L.; et al. New insights into mechanisms of sunlight- and dark-mediated high-temperature can accelerate diurnal production-degradation transformation of lake fluorescent DOM. Sci. Total Environ. 2021, 760, 143377. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Nail. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—Current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Scandellari, F. Arbuscular mycorrhizal contribution to nitrogen uptake of grapevines. Vitis 2017, 56, 147–154. [Google Scholar] [CrossRef]
- Le Bissonnais, Y.; Montier, C.; Jamague, M.; Daroussin, J.; King, D. Mapping erosion risk for cultivated soil in France. Catena 2001, 46, 207–220. [Google Scholar] [CrossRef]
- Brenot, J.; Quiquerez, A.; Petit, C.; Garcia, J.P. Erosion rates and sediment budgets in vineyards at 1-m resolution based on stock unearthing (Burgundy, France). Geomorphology 2008, 100, 345–355. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Kurtural, S.K. Inoculation with mycorrhizal fungi and irrigation management shape the bacterial and fungal communities and networks in vineyard soils. Microorganisms 2021, 9, 1273. [Google Scholar] [CrossRef] [PubMed]
- Oehl, F.; Koch, B. Diversity of arbuscular mycorrhizal fungi in no-till and conventionally tilled vineyards. J. Appl. Bot. Food Qual. 2018, 91, 56–60. [Google Scholar] [CrossRef]
- Schreiner, R.P. Effects of native and nonnative arbuscular mycorrhizal fungi on growth and nutrient uptake of “Pinot noir” (Vitis vinifera L.) in two soils with contrasting levels of phosphorous. Appl. Soil Ecol. 2007, 36, 205–215. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mumey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Petgen, M.; Schropp, A.; Marschner, H.; Roemheld, V. Investigations on the Occurrence of Arbuscular Mycorrhizae in Some Grape Vine Nurseries and the Practical Management of Field Inoculation with Arbuscular Mycorrhizae; Mitteilungen-Biologische Bundesanstalt für Land-und Forstwirtschaft: Berlin, Germany, 1997; pp. 32–46. [Google Scholar]
- Cavagnaro, T.R.; Smith, F.A.; Smith, S.E.; Jakobsen, I. Functional diversity in arbuscular mycorrhizas: Exploitation of soil patches with different phosphate enrichment differs among fungal species. Plant. Cell Environ. 2005, 28, 642–650. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Eviner, V.T. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: Comparing effects of five plant species. Plant Soil 2002, 238, 325–333. [Google Scholar] [CrossRef]
- Caravaca, F.; Alguacil, M.M.; Azcón, R.; Roldán, A. Formation of stable aggregates in rhizosphere soil of Juniperus oxycedrus: Effect of AM fungi and organic amendments. Appl. Soil Ecol. 2006, 33, 30–38. [Google Scholar] [CrossRef]
- Bedini, S.; Pellegrino, E.; Avio, L.; Pellegrini, S.; Bazzoffi, P.; Argese, E.; Giovannetti, M. Changes in soil aggregation and glomalin-related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol. Biochem. 2009, 41, 1491–1496. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Bergmann, J.; Verbruggen, E.; Veresoglou, S.D.; Lehmann, A. (2014) Plant root and mycorrhizal fungal traits for understanding soil aggregation. New Phytol. 2014, 205, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Trouvelot, S.; Bonneau, L.; Redecker, D.; van Tuinen, D.; Adrian, M.; Wipf, D. Arbuscular mycorrhiza symbiosis in viticulture: A review. Agron. Sustain. Dev. 2015, 35, 1449–1467. [Google Scholar] [CrossRef]
- Bettenfeld, P.; Cadena I Canals, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.E.; Trouvelot, S. The microbiota of the grapevine holobiont: A key component of plant health. J. Adv. Res. 2022, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hanada, R.E.; Pomelia, A.W.V.; Soberanis, W.; Loguercio, L.L.; Pereira, J.O. Biocontrol potential of Trichoderma martiale against the black-pod disease (Phytophthora palmivora) of cacao. Biol. Control 2009, 50, 143–149. [Google Scholar] [CrossRef]
- Gadoury, D.M.; Seem, R.C.; Pearson, R.C.; Wilcox, W.F.; Dunst, R.M. Effects of powdery mildew on vine growth, yield, and quality of concord grapes. Plant Dis. 2001, 85, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Steel, C.C.; Blackman, J.W.; Schmidtke, L.M. Grapevine bunch rots: Impacts on wine composition, quality, and potential procedures for the removal of wine faults. J. Agric. Food Chem. 2013, 61, 5189–5206. [Google Scholar] [CrossRef] [PubMed]
- Darriaut, R.; Lailheugue, V.; Masneuf-Pomarède, I.; Marguerit, E.; Martins, G.; Company, S.; Ballestra, P.; Upton, S.; Ollat, N.; Lauvergeat, V. Grapevine rootstock and soil microbiome interactions: Keys for a resilient viticulture. Hortic. Res. 2022, 9, uhac019. [Google Scholar] [CrossRef]
- Mutawila, C.; Halleen, F.; Mostert, L. Optimisation of time of application of Trichoderma biocontrol agents for protection of grapevine pruning wounds. Aust. J. Grape Wine Res. 2016, 22, 279–287. [Google Scholar] [CrossRef]
- Perazzolli, M.; Antonielli, L.; Storari, M.; Puopolo, G.; Pancher, M.; Giovannini, O.; Pindo, M.; Pertot, I. Resilience of the natural phyllosphere microbiota of the grapevine to chemical and biological pesticides. Appl. Environ. Microbiol. 2014, 80, 3585–3596. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionst. Nat. Rev. Microbiol. 2004, 2, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Carro-Huerga, G.; Mayo-Prieto, S.; Rodríguez-González, Á.; Cardoza, R.E.; Gutiérrez, S.; Casquero, P.A. Vineyard management and physicochemical parameters of soil affect native Trichoderma populations, sources of biocontrol agents against Phaeoacremonium minimum. Plants 2023, 12, 887. [Google Scholar] [CrossRef] [PubMed]
- Döring, J.; Collins, C.; Frisch, M.; Kauer, R. Organic and biodynamic viticulture affect biodiversity and properties of vine and wine: A systematic quantitative review. Am. J. Enol. Vitic. 2019, 70, 3. [Google Scholar] [CrossRef]
- Pancher, M.; Coel, M.; Corneo, P.E.; Longa, C.M.O.; Yousaf, S.; Pertot, I.; Campisano, A. Fungal endophytic communities in grapevines (Vitis vinifera L.) respond to crop management. Appl. Environ. Microbiol. 2012, 78, 4308–4317. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.; Custódio, V.; Nunes, M.; Songy, A.; Rabenoelina, F.; Courteaux, B.; Clément, C.; Gomes, A.C.; Fontaine, F. Understand the potential role of Aureobasidium pullulans, a resident microorganism from grapevine, to prevent the infection caused by Diplodia seriata. Front. Microbiol. 2018, 9, 3047. [Google Scholar] [CrossRef] [PubMed]
- Kernaghan, G.; Mayerhofer, M.; Griffin, A. Fungal endophytes of wild and hybrid Vitis leaves and their potential for vineyard biocontrol. Can. J. Microbiol. 2017, 63, 583–8595. [Google Scholar] [CrossRef] [PubMed]
- Point, E.; Tyedmers, P.; Naugler, C. Life cycle environmental impacts of wine production and consumption in Nova Scotia, Canada. J. Clean. Prod. 2012, 27, 11–20. [Google Scholar] [CrossRef]
- De la Huerta-Bengoechea, P.; Gil-Serna, J.; Melguizo, C.; Ramos, A.J.; Prim, M.; Vázquez, C.; Patiño, B. Biocontrol of mycotoxigenic fungi using bacteria isolated from ecological vineyard soils. J. Fungi 2022, 8, 1136. [Google Scholar] [CrossRef]
- Maluleke, E.; Jolly, N.P.; Patterton, H.G.; Setati, M.E. Antifungal activity of non-conventional yeasts against Botrytis cinerea and non-Botrytis grape bunch rot fungi. Front. Microbiol. 2022, 13, 986229. [Google Scholar] [CrossRef]
- Tempesta, T.; Giancristofaro, R.A.; Corain, L.; Salmaso, L.; Tomasi, D.; Boatto, V. The importance of landscape in wine quality perception: An integrated approach using choice-based conjoint analysis and combination-based permutation tests. Food Qual. Prefer. 2010, 21, 827–836. [Google Scholar] [CrossRef]
- Montella, M.M. Wine tourism and sustainability: A review. Sustainability 2017, 9, 113. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Friant, P.; Chone, X.; Tregoat, O.; Koundouras, S.; Dubourdieu, D. Influence of climate, soil and cultivar on terroir. Am. J. Enol. Vitic. 2004, 55, 207–217. [Google Scholar] [CrossRef]
- Rosso, A.M. Beer and wine in antiquity: Beneficial remedy or punishment imposed by the Gods? Acta Med.-Hist. Adriat. 2012, 10, 237–262. [Google Scholar] [PubMed]
- Stanislawski, D. Dionysus westward: Early religion and the economic geography of wine. Geogr. Rev. 1975, 65, 4. [Google Scholar] [CrossRef]
- Kirkpatrick, J. The jews and their god of wine. Arch. Für Relig. 2014, 15, 167–186. [Google Scholar] [CrossRef]
- Onofri, L.; Boatto, V. On the economic valuation of cultural ecosystem services: A tale of myths, vine and wine. Ecosyst. Serv. 2020, 46, 101215. [Google Scholar] [CrossRef]
- Chou, M.Y.; Vanden Heuvel, J.; Bell, T.H.; Panke-Buisse, K.; Kao-Kniffin, J. Vineyard under-vine floor management alters soil microbial composition, while the fruit microbiome shows no corresponding shifts. Sci. Rep. 2018, 8, 11039. [Google Scholar] [CrossRef]
- Quiquerez, A.; García, J.P.; Dequiedt, S.; Djemiel, C.; Terrat, S.; Mathieu, O.; Sassi, A.; Ranjard, L. Legacy of land-cover changes on soil microbiology in Burgundy vineyards (Pernand-Vergelesses, France). OENO One 2022, 56, 2. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Arroyo, T.; Serrano, A.; Valero, E. Influence of different floor management strategies of the vineyard on the natural yeast population associated with grape berries. Int. J. Food Microbiol. 2011, 148, 23–29. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef]
- Bagheri, B.; Bauer, F.; Setati, M. The diversity and dynamics of indigenous yeast communities in grape must from vineyards employing different agronomic practices and their influence on wine fermentation. S. Afr. J. Enol. Vitic. 2015, 36, 243–251. [Google Scholar] [CrossRef]
- Capó-Bauçà, S.; Marqués, A.; Llopis-Vidal, N.; Bota, J.; Baraza, E. Long-term establishment of natural green cover provides agroecosystem services by improving soil quality in a Mediterranean vineyard. Ecol. Eng. 2019, 127, 285–291. [Google Scholar] [CrossRef]
- Daane, K.M.; Hogg, B.N.; Wilson, H.; Yokota, G.Y. Native grass ground covers provide multiple ecosystem services in Californian vineyards. J. Appl. Ecol. 2018, 55, 2473–2483. [Google Scholar] [CrossRef]
- Rodríguez-Entrena, M.; Colombo, S.; Arriaza, M. The landscape of olive groves as a driver of the rural economy. Land Use Policy 2017, 65, 164–175. [Google Scholar] [CrossRef]
- Chapela-Oliva, C.; Winter, S.; Ochoa-Hueso, R. Edaphoclimatic drivers of the effect of extensive vegetation management on ecosystem services and biodiversity in vineyards. Agric. Ecosyst. Environ. 2022, 339, 108115. [Google Scholar] [CrossRef]
- Bünemann, E.; Schwenke, G.; Zwieten, L. Impact of agricultural inputs on soil organisms—A review. Aust. J. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef]
- Okur, N.; Altindİşlİ, A.; Çengel, M.; Göçmez, S.; Kayikçioğlu, H.H. Microbial biomass and enzyme activity in vineyard soils under organic and conventional farming systems. Turk. J. Agric. For. 2009, 33, 413–423. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- García-Orenes, F.; Roldán, A.; Morugán-Coronado, A.; Linares, C.; Cerdà, A.; Caravaca, F. Organic fertilization in traditional Mediterranean grapevine orchards mediates changes in soil microbial community structure and enhances soil fertility. Land Degrad. Dev. 2016, 27, 1622–1628. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Yeoh, Y.K.; Kasinadhuni, N.R.; Lonhienne, T.G.; Robinson, N.; Hugenholtz, P.; Ragan, M.A.; Schmidt, S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Sci. Rep. 2015, 5, 8678. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Ivan, D.; Zottini, M. Management intensity and topography determined plant diversity in vineyards. PLoS ONE 2013, 8, e76167. [Google Scholar] [CrossRef] [PubMed]
- Amaral, H.F.; Schwan-Estrada, K.R.F.; de Sena, J.O.A.; Colozzi-Filho, A.; Andrade, D.S. Seasonal variations in soil chemical and microbial indicators under conventional and organic vineyards. Acta Sci.-Agron. 2023, 45, e56158. [Google Scholar] [CrossRef]
- Karimi, B.; Masson, V.; Guilland, C.; Leroy, E.; Pellegrinelli, S.; Giboulot, E.; Maron, P.A.; Ranjard, L. Ecotoxicity of copper input and accumulation for soil biodiversity in vineyards. Environ. Chem. Lett. 2021, 19, 2013–2030. [Google Scholar] [CrossRef]
- Ostandie, N.; Giffard, B.; Bonnard, O.; Joubard, B.; Richart-Cervera, S.; Thiéry, D.; Rusch, A. Multi-community effects of organic and conventional farming practices in vineyards. Sci. Rep. 2021, 11, 11979. [Google Scholar] [CrossRef] [PubMed]
- Milanovic, V.; Comitini, F.; Ciani, M. Grape berry yeast communities: Influence of fungicide treatments. Int. J. Food. Microbiol. 2013, 161, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Mandl, K.; Cantelmo, C.; Gruber, E.; Faber, F.; Friedrich, B.; Zaller, J.G. Effects of glyphosate-, glufosinate- and flazasulfuron-based herbicides on soil microorganisms in a vineyard. Bull. Env. Contam. Toxic. 2018, 101, 562–569. [Google Scholar] [CrossRef]
- Probst, B.; Schüler, C.; Joergensen, R.G. Vineyard soils under organic and conventional management—Microbial biomass and activity indices and their relation to soil chemical properties. Biol. Fert. Soils 2008, 44, 443–450. [Google Scholar] [CrossRef]
- Vega-Avila, A.D.; Gumiere, T.; Andrade, P.A.M.; Lima-Perim, J.E.; Durrer, A.; Baigori, M.; Vazquez, F.; Andreote, F.D. Bacterial communities in the rhizosphere of Vitis vinifera L. cultivated under distinct agricultural practices in Argentina. Antonie Leeuwenhoek J. Microb. 2015, 107, 575–588. [Google Scholar] [CrossRef]
- Patrignani, F.; Montanari, C.; Serrazanetti, D.I.; Braschi, G.; Vernocchi, P.; Tabanelli, G.; Parpinello, G.P.; Versari, A.; Gardini, F.; Lanciotti, R. Characterisation of yeast microbiota, chemical and sensory properties of organic and biodynamic Sangiovese red wines. Ann. Microbiol. 2016, 67, 99–109. [Google Scholar] [CrossRef]
- Schmid, F.; Moser, G.; Müller, H.; Berg, G. Functional and structural microbial diversity in organic and conventional viticulture: Organic farming benefits natural biocontrol agents. Appl. Environ. Microb. 2011, 77, 2188–2191. [Google Scholar] [CrossRef]
- Power, A.G. Ecosystem services and agriculture: Trade-offs and synergies. Phil. Trans. R. Soc. B 2010, 365, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Viers, J.H.; Williams, J.N.; Nicholas, K.A.; Barbosa, O.; Kotzé, I.; Spence, L.; Webb, L.B.; Merenlender, A.; Reynolds, M. Vinecology: Pairing wine with nature. Conserv. Lett. 2013, 6, 287–299. [Google Scholar] [CrossRef]
- Cerdan, O.; Govers, G.; Le Bissonnais, Y.; Van Oost, K.; Poesen, J.; Saby, N.; Gobin, A.; Vacca, A.; Quinton, J.; Auerswald, K.; et al. Rates and spatial variations of soil erosion in Europe: A study based on erosion plot data. Geomorphology 2010, 122, 167–177. [Google Scholar] [CrossRef]
- Biddoccu, M.; Ferraris, S.; Opsi, F.; Cavallo, E. Long-term monitoring of soil management effects on runoff and soil erosion in sloping vineyards in Alto Monferrato (North–West Italy). Soil Till. Res. 2016, 155, 176–189. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Litskas, V.; Mandoulaki, A.; Antoniou, D.; Boyias, T.; Stavrinides, M.; Tzortzakis, N. Drought stress and soil management practices in grapevines in Cyprus under the threat of climate change. J. Water Clim. Chang. 2018, 9, 703–714. [Google Scholar] [CrossRef]
- Wolfert, S.; Ge, L.; Verdouw, C.; Bogaardt, M.J. Big data in smart farming—A review. Agric. Syst. 2017, 153, 69–80. [Google Scholar] [CrossRef]
- Hartman, K.; van der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.C.; Schlaeppi, K. Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 2018, 6, 14. [Google Scholar] [CrossRef]
- Guerry, A.D.; Polasky, S.; Lubchenco, J.; Chaplin-Kramer, R.; Daily, G.C.; Griffin, R.; Ruckelshaus, M.; Bateman, I.J.; Duraiappah, A.; Elmqvist, T.; et al. Natural capital and ecosystem services informing decisions: From promise to practice. Proc. Natl. Acad. Sci. USA 2015, 112, 7348–7355. [Google Scholar] [CrossRef]
- Chen, H.; Ma, K.; Lu, C.; Fu, Q.; Qiu, Y.; Zhao, J.; Huang, Y.; Yang, Y.; Schadt, C.W.; Chen, H. Functional redundancy in soil microbial community based on metagenomics across the globe. Front. Microbiol. 2022, 13, 878978. [Google Scholar] [CrossRef]
- Grosskopf, T.; Soyer, O.S. Synthetic microbial communities. Curr. Opin. Microbiol. 2014, 18, 72–77. [Google Scholar] [CrossRef]
- Gonçalves, O.S.; Creevey, C.J.; Santana, M.F. Designing a synthetic microbial community through genome metabolic modeling to enhance plant-microbe interaction. Environ. Microbiome 2023, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Suman, A.; Govindasamy, V.; Ramakrishnan, B.; Aswini, K.; SaiPrasad, J.; Sharma, P.; Pathak, D.; Annapurna, K. Microbial Community and Function-Based Synthetic Bioinoculants: A Perspective for Sustainable Agriculture. Front. Microbiol. 2022, 12, 805498. [Google Scholar] [CrossRef] [PubMed]
- Carlström, C.I.; Field, C.M.; Bortfeld-Miller, M.; Müller, B.; Sunagawa, S.; Vorholt, J.A. Synthetic microbiota reveal priority effects and keystone strains in the Arabidopsis phyllosphere. Nat. Ecol. Evol. 2019, 3, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, T.; Devers-Lamrani, M.; Spor, A.; Blouin, M. Community diversity determines the evolution of synthetic bacterial communities under artificial selection. Evolution 2022, 76, 1883–1895. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.S.K.; Pretorius, I.S. Synthetic biology for the engineering of complex wine yeast communities. Nat. Food 2022, 3, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Dixon, T.A.; Walker, R.S.K.; Pretorius, I.S. Visioning synthetic futures for yeast research within the context of current global technolopolitical trends. Yeast 2023, 40, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Suman, J.; Rakshit, A.; Ogireddy, S.D.; Singh, S.; Gupta, C.; Chandrakala, J. Microbiome as a key player in sustainable agriculture and human health. Front. Soil Sci. 2022, 2, 821589. [Google Scholar] [CrossRef]
- Traeger, C.P. On option values in environmental and resource economics. Resour. Energy Econ. 2014, 37, 242–252. [Google Scholar] [CrossRef]
- Pascual, U.; Termansen, M.; Hedlund, K.; Brussaard, L.; Faber, J.H.; Foudi, S.; Lemanceau, P.; Jørgensen, S.L. On the value of soil biodiversity and ecosystem services. Ecosyst. Serv. 2015, 15, 11–18. [Google Scholar] [CrossRef]
- Baumgärtner, S.; Strunz, S. The economic insurance value of ecosystem resilience. Ecol. Econ. 2014, 101, 21–32. [Google Scholar] [CrossRef]
- Dallimer, M.; Martin-Ortega, J.; Rendon, O.; Afionis, S.; Bark, R.; Gordon, I.J.; Paavola, J. Taking stock of the empirical evidence on the insurance value of ecosystems. Ecol. Econ. 2020, 167, 106451. [Google Scholar] [CrossRef]
- Paul, C.; Hanley, N.; Meyer, S.T.; Fürst, C.; Weisser, W.W.; Knoke, T. On the functional relationship between biodiversity and economic value. Sci. Adv. 2020, 6, 7712. [Google Scholar] [CrossRef] [PubMed]
- Primmer, E.; Paavola, J. Insurance value of ecosystems: An introduction. Ecol. Econ. 2021, 184, 107001. [Google Scholar] [CrossRef]
- Baumgärtner, S. The insurance value of biodiversity in the provision of ecosystem services. Nat. Resour. Model. 2007, 20, 87–127. [Google Scholar] [CrossRef]
- Colwell, R.R. Microbial diversity: The importance of exploration and conservation. J. Ind. Microbiol. Biotechnol. 1997, 18, 302–307. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Izquierdo, I.; Colino-Rabanal, V.J.; Tamame, M.; Rodríguez-López, F. Microbiota Ecosystem Services in Vineyards and Wine: A Review. Agronomy 2024, 14, 131. https://doi.org/10.3390/agronomy14010131
García-Izquierdo I, Colino-Rabanal VJ, Tamame M, Rodríguez-López F. Microbiota Ecosystem Services in Vineyards and Wine: A Review. Agronomy. 2024; 14(1):131. https://doi.org/10.3390/agronomy14010131
Chicago/Turabian StyleGarcía-Izquierdo, Isabel, Victor J. Colino-Rabanal, Mercedes Tamame, and Fernando Rodríguez-López. 2024. "Microbiota Ecosystem Services in Vineyards and Wine: A Review" Agronomy 14, no. 1: 131. https://doi.org/10.3390/agronomy14010131
APA StyleGarcía-Izquierdo, I., Colino-Rabanal, V. J., Tamame, M., & Rodríguez-López, F. (2024). Microbiota Ecosystem Services in Vineyards and Wine: A Review. Agronomy, 14(1), 131. https://doi.org/10.3390/agronomy14010131