
A CRISPR-Cas9-Derived Male Sterility System for Tomato Breeding

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. CRISPR-Derived Dyt1Gstaa Mutants Were Generated in Elite Inbred Lines
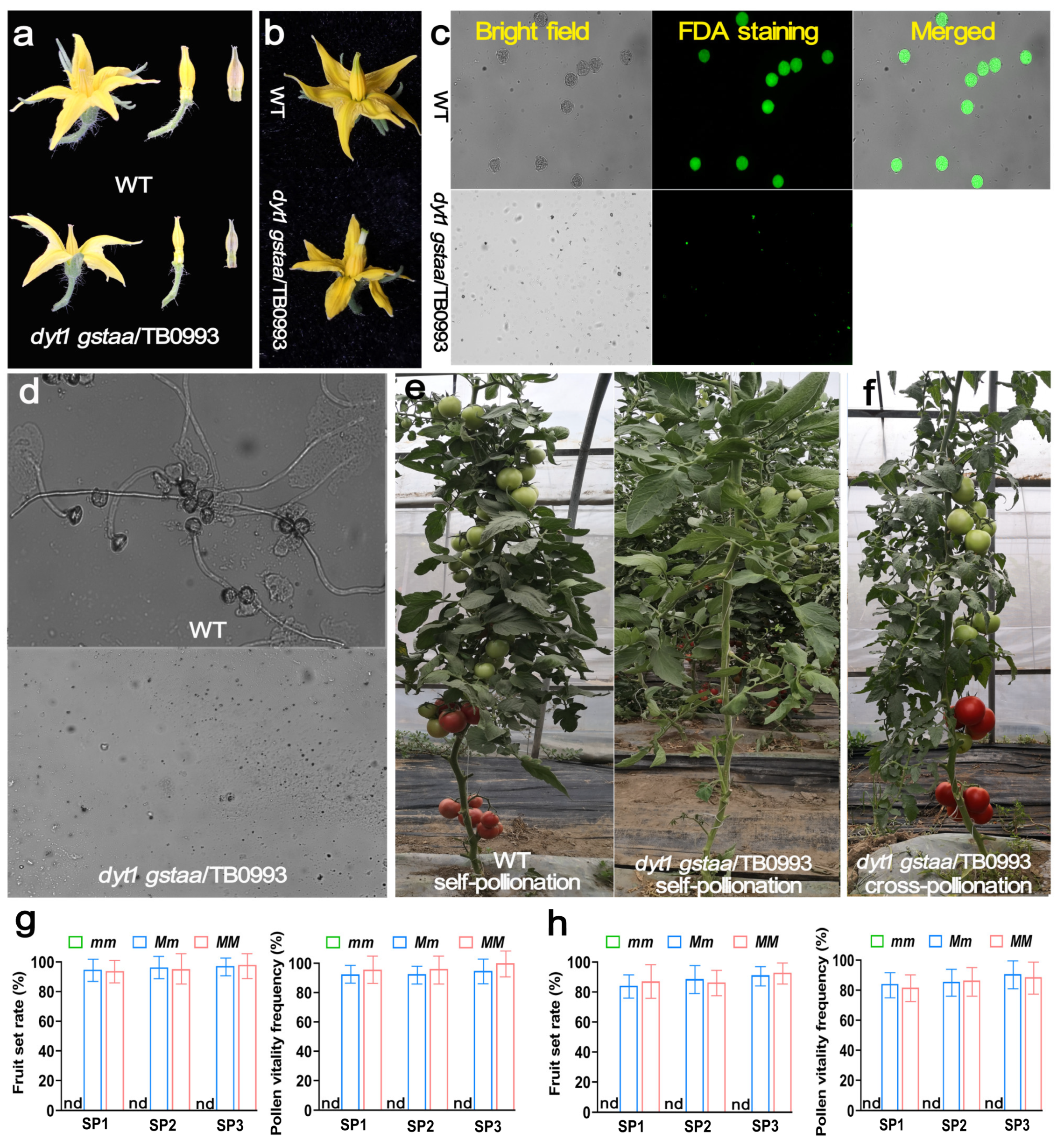
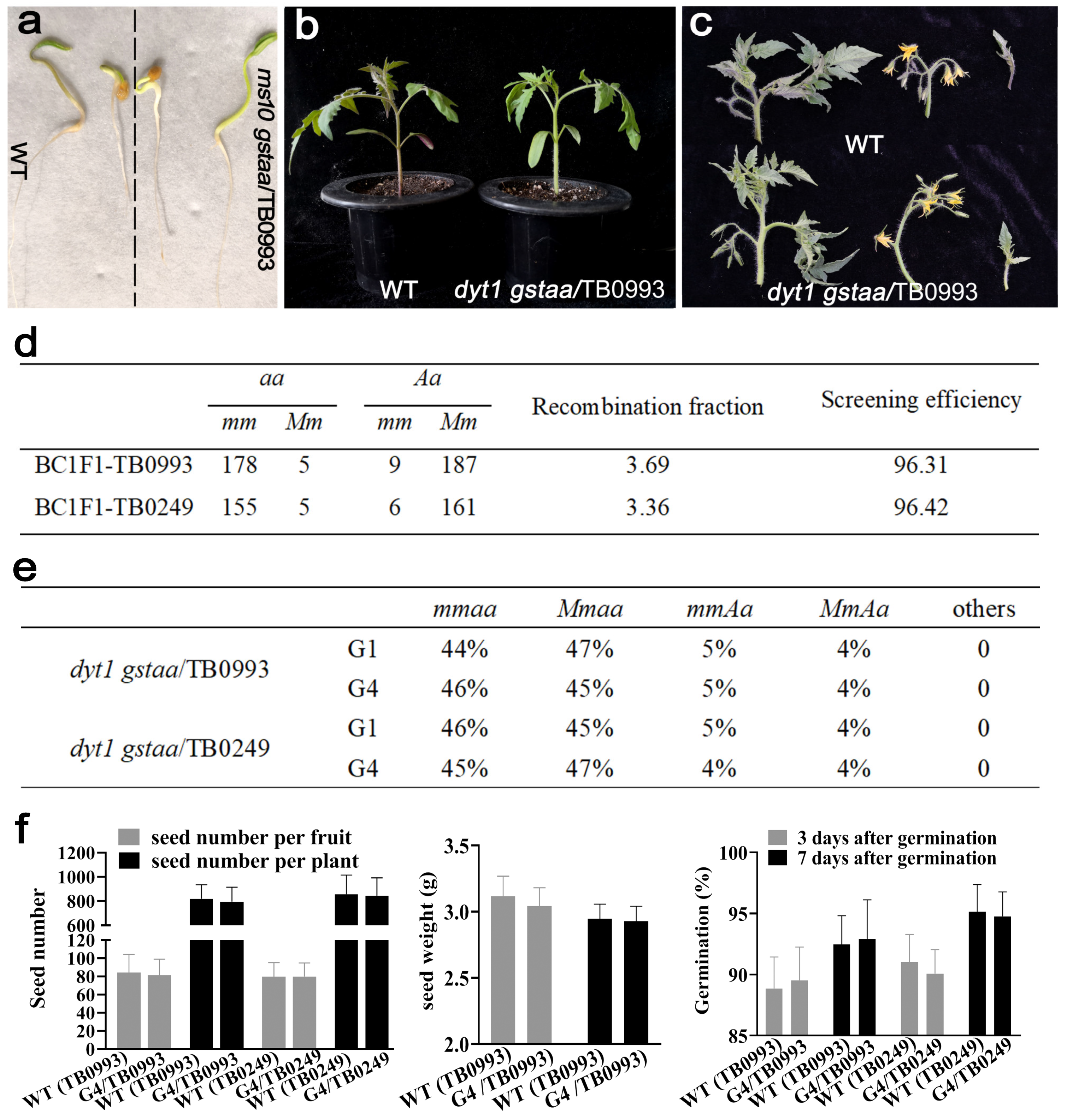
2.2. CRISPR-Derived Dyt1Gstaa Mutants Exhibited Complete and Stable Male Sterility
2.3. Propagation of the Male Sterility Lines
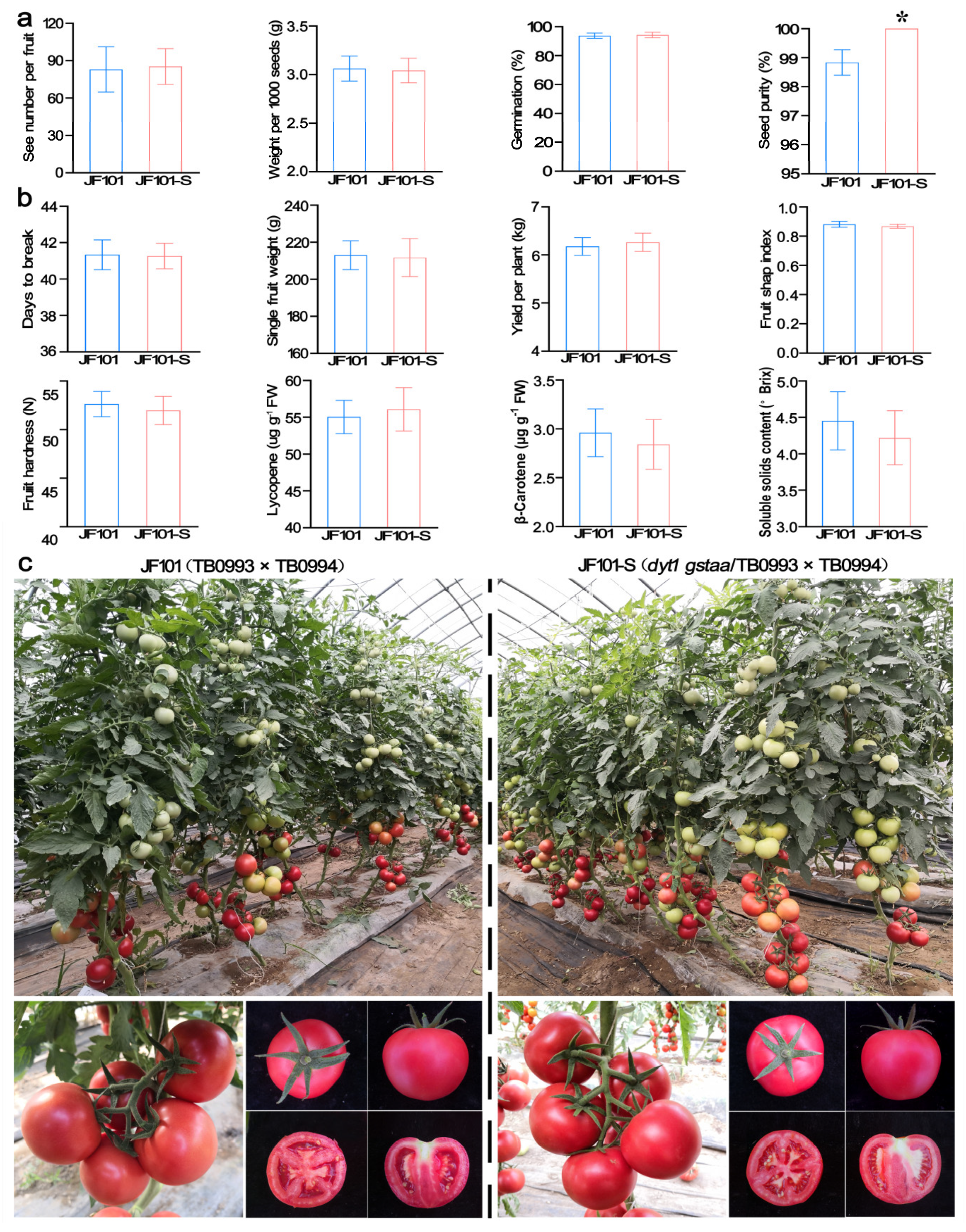
2.4. The Created Dyt1Gstaa System Had No Side Effects on Main Agronomic Traits When Used for Hybrid Production
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. CRISPR/Cas9-Mediated Mutagenesis
4.3. Pollen Vitality Analysis
4.4. Linkage Analysis
4.5. Assessment of Side Effects on Seed Yield, Seed Quality, and Agronomic Traits
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tamta, S.; Singh, J.P. Heterosis in tomato for growth and yield traits. Int. J. Veg. Sci. 2018, 24, 169–179. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant. Biol. 2014, 65, 1473–1475. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Zhang, D. Molecular Control of male fertility for crop hybrid breeding. Trends Plant. Sci. 2018, 23, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.H.; Song, W.Y.; Zhang, X.L. Genic male and female sterility in vegetable crops. Hortic. Res. 2022, 10, uhac232. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, B. Functional male sterility ps-2 in tomato (Lycopesicon esculentum Mill.) and its application in breeding and hybrid seed production. Euphytica 1999, 107, 13–21. [Google Scholar] [CrossRef]
- Gorguet, B.; Schipper, D.; van Lammeren, A.; Visser, R.G.F.; van Heusden, A.W. ps-2, the gene responsible for functional sterility in tomato, due to non-dehiscent anthers, is the result of a mutation in a novel polygalacturonase gene. Theor. Appl. Genet. 2009, 118, 1199–1209. [Google Scholar] [CrossRef]
- Liu, X.Y.; Yang, M.X.; Liu, X.L.; Wei, K.; Cao, X.; Wang, X.T.; Wang, X.X.; Guo, Y.M.; Du, Y.C.; Li, J.M.; et al. A putative bHLH transcription factor is a candidate gene for male sterile32, a locus affecting pollen and tapetum development in tomato. Hortic. Res. 2019, 6, 88. [Google Scholar] [CrossRef]
- Cao, X.; Liu, X.Y.; Wang, X.T.; Giang, T.V.; Wang, J.; Liu, X.L.; Sun, S.; Wei, K.; Wang, X.X.; Gao, J.C.; et al. B-class MADS-box TM6 is a candidate gene for tomato male sterile-1526. Theor. Appl. Genet. 2019, 132, 2125–2135. [Google Scholar] [CrossRef]
- Pucci, A.; Picarella, M.E.; Mazzucato, A. Phenotypic, genetic and molecular characterization of 7B-1, a conditional male-sterile mutant in tomato. Theor. Appl. Genet. 2017, 130, 2361–2374. [Google Scholar] [CrossRef]
- Perez-Prat, E.; van Lookeren Campagne, M.M. Hybrid seed production and the challenge of propagating male-sterile plants. Trends Plant. Sci. 2002, 7, 199–203. [Google Scholar] [CrossRef]
- Atanassova, B. Genic male sterility and its application in tomato (Lycopersicon esculentum mill.) hybrid breeding and hybrid seed production. In Proceedings of the 3rd Balkan Symposium on Vegetable and Potatoes, Bursa, Turkey, 4–6 September 2004; International Society for Horticultural Science: Korbeek-Lo, Belgium, 2004; pp. 45–51. [Google Scholar]
- Jeong, H.J.; Kang, J.H.; Zhao, M.; Kwon, J.K.; Choi, H.S.; Bae, J.H.; Lee, H.; Joung, Y.H.; Choi, D.; Kang, B.C. Tomato male sterile 1035 is essential for pollen development and meiosis in anthers. J. Exp. Bot. 2014, 65, 6693–6709. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Huang, Z.J.; Wang, X.X.; Gao, J.C.; Guo, Y.M.; Du, Y.C.; Hu, H. Fine mapping and molecular marker development of anthocyanin absent, a seedling morphological marker for the selection of male sterile 10 in tomato. Mol. Breed. 2016, 36, 107. [Google Scholar] [CrossRef]
- Aglawe, S.B.; Barbadikar, K.M.; Mangrauthia, S.K.; Madhav, M.S. New breeding technique “genome editing” for crop improvement: Applications, potentials and challenges. 3 Biotech. 2018, 8, 336. [Google Scholar] [CrossRef]
- Gao, C.X. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Xia, X.H.; Cheng, X.H.; Li, R.; Yao, J.N.; Li, Z.G.; Cheng, Y.L. Advances in application of genome editing in tomato and recent development of genome editing technolog. Theor. Appl. Genet. 2021, 134, 2727–2747. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Singh, A.K.; Tusar, K.B. CRISPR/Cas genome editing in tomato improment: Advances and applications. Front. Plant. Sci. 2023, 14, 1121209. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Kim, D.H.; Lee, H.J.; Nam, K.H.; Bae, S.; Nou, I.S.; Cho, Y.G.; Kim, Y.K.; Kang, O.K. Knockout of SlMS10 gene (Solyc02g079810) encoding bHLH transcription factor using CRISPR/Cas9 system confers male sterility phenotype in tomato. Plants 2020, 9, 1189. [Google Scholar] [CrossRef]
- Bao, H.; Ding, Y.; Yang, F.; Zhang, J.; Xie, J.; Zhao, C.; Du, K.H.; Zeng, Y.W.; Zhao, K.; Li, Z.S.; et al. Gene silencing, knockout and over-expression of a transcription factor ABORTED MICROSPORES (SlAMS) strongly affects pollen viability in tomato (Solanum lycopersicum). BMC Genom. 2022, 23, 346. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Han, H.; Huang, W.; Zhao, L.; Song, M.; Cao, X.; Liu, C.L.; Niu, X.M.; Lang, Z.B.; Ma, C.L.; et al. Generating novel male sterile tomatoes by editing respiratory burst oxidase homolog genes. Front. Plant. Sci. 2022, 12, 817101. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.H.; Zhao, J.G.; Chen, Z.W.; Qiao, J.; Zhang, Y.F.; Shen, H.; Hu, Z.L. CRISPR/Cas9-targeted mutagenesis of SlCMT4 causes changes in plant architecture and reproductive organs in tomato. Hortic. Res. 2022, 9, uhac081. [Google Scholar] [CrossRef]
- Du, M.M.; Zhou, K.; Liu, Y.Y.; Deng, L.; Zhang, X.Y.; Lin, L.H.; Zhou, M.; Zhao, W.; Wen, C.L.; Xing, J.Y.; et al. A biotechnology-based male-sterility system for hybrid seed production in tomato. Plant. J. 2020, 102, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.W.; Wang, S.F.; Wang, H.; Luo, B.; Cai, Y.Y.; Li, X.D.; Zhang, Y.F.; Wang, X.F. Rapid generation of tomato male-sterile lines with a marker use for hybrid seed production by CRISPR/Cas9 system. Mol. Breed. 2021, 41, 25. [Google Scholar] [CrossRef] [PubMed]
- Mutschler, M.A.; Rick, C.M.; Tanksley, S.D. Linkage maps of the tomato (Lycopersicon esculentum). Rep. Tomato Genet. Coop. 1987, 37, 5–34. [Google Scholar]
- Cui, J.; You, C.; Zhu, E.; Huang, Q.; Ma, H.; Chang, F. Feedback regulation of DYT1 by interactions with downstream bHLH factors promotes DYT1 nuclear localization and anther development. Plant. Cell. 2016, 28, 1078–1093. [Google Scholar] [CrossRef]
- Jung, K.H.; Han, M.J.; Lee, Y.S.; Kim, Y.W.; Hwang, I.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; An, G. Rice Undeveloped Tapetum1 is a major regulator of early tapetum development. Plant. Cell. 2005, 17, 2705–2722. [Google Scholar] [CrossRef]
- Hao, S.; Ariizumi, T.; Ezura, H. Sexual sterility is essential for both male and female gametogenesis in tomato. Plant. Cell. Physiol. 2017, 58, 22–34. [Google Scholar]
- Okabe, Y.; Yamaoka, T.; Ariizumi, T.; Ushijima, K.; Kojima, M.; Takebayashi, Y.; Sakakibara, H.; Kusano, M.; Shinozaki, Y.; Pulungan, S.I.; et al. Aberrant stamen development is associated with parthenocarpic fruit set through up-regulation of gibberellin biosynthesis in tomato. Plant. Cell. Physiol. 2019, 60, 38–51. [Google Scholar] [CrossRef]
- Rojas-Gracia, P.; Roque, E.; Medina, M.; Rochina, M.; Hamza, R.; Angarita-Díaz, M.P.; Moreno, V.; Perez-Martín, F.; Lozano, R.; Canas, L.; et al. The parthenocarpic hydra mutant reveals a new function for a SPOROCYTELESS-like gene in the control of fruit set in tomato. New. Phytol. 2017, 214, 1198–1212. [Google Scholar] [CrossRef]
- Salazar-Sarasua, B.; López-Martín, M.J.; Roque, E.; Hamza, R.; Cañas, L.A.; Beltrán, J.P.; Gomez-Mena, C. The tapetal tissue is essential for the maintenance of redox homeostasis during microgametogenesis in tomato. Plant. J. 2022, 1, 1281–1297. [Google Scholar] [CrossRef]
- Deng, L.; Wang, H.; Sun, C.L.; Li, Q.; Jiang, H.L.; Du, M.M.; Li, C.B.; Li, C.Y. Efficient generation of pink-fruited tomatoes using CRISPR/Cas9 system. J. Genet. Genomics 2018, 45, 51–54. [Google Scholar] [CrossRef]
- Li, X. Pollen fertility/viability assay using FDA staining. Bio Protoc. 2011, 1, e75. [Google Scholar] [CrossRef]
- Song, J.; Nada, K.; Tachibana, S. Ameliorative effect of polyamines on the high temperature inhibition of in vitro pollen germination in tomato (Lycopersicon esculentum Mill.). Sci. Hortic. 1999, 80, 203–212. [Google Scholar] [CrossRef]
- Zhou, M.; Deng, L.; Guo, S.G.; Yuan, G.L.; Li, C.Y.; Li, C.B. Alternative transcription and feedback regulation suggest that SlIDI1 is involved in tomato carotenoid synthesis in a complex way. Hortic. Res. 2022, 9, Uhab045. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Deng, L.; Yuan, G.; Zhao, W.; Ma, M.; Sun, C.; Du, M.; Li, C.; Li, C. A CRISPR-Cas9-Derived Male Sterility System for Tomato Breeding. Agronomy 2023, 13, 1785. https://doi.org/10.3390/agronomy13071785
Zhou M, Deng L, Yuan G, Zhao W, Ma M, Sun C, Du M, Li C, Li C. A CRISPR-Cas9-Derived Male Sterility System for Tomato Breeding. Agronomy. 2023; 13(7):1785. https://doi.org/10.3390/agronomy13071785
Chicago/Turabian StyleZhou, Ming, Lei Deng, Guoliang Yuan, Wei Zhao, Mingyang Ma, Chuanlong Sun, Minmin Du, Chuanyou Li, and Changbao Li. 2023. "A CRISPR-Cas9-Derived Male Sterility System for Tomato Breeding" Agronomy 13, no. 7: 1785. https://doi.org/10.3390/agronomy13071785
APA StyleZhou, M., Deng, L., Yuan, G., Zhao, W., Ma, M., Sun, C., Du, M., Li, C., & Li, C. (2023). A CRISPR-Cas9-Derived Male Sterility System for Tomato Breeding. Agronomy, 13(7), 1785. https://doi.org/10.3390/agronomy13071785