
Genetic Diversity of Colletotrichum spp. Causing Grape Anthracnose in Zhejiang, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Fungus Isolation
2.2. Molecular Characterization
2.3. Multi-Locus Phylogenetic Analysis
2.4. Morphological Characterization
2.5. Mycelial Growth, Sporulation and Conidial Germination of Colletotrichum spp.
2.6. Pathogenicity Testing
2.7. Statistical Analysis
3. Results
3.1. Colletotrichum Isolates Associated with Grape Anthracnose
3.2. Multi-Locus Phylogenetic Analyses
3.3. Species Distribution
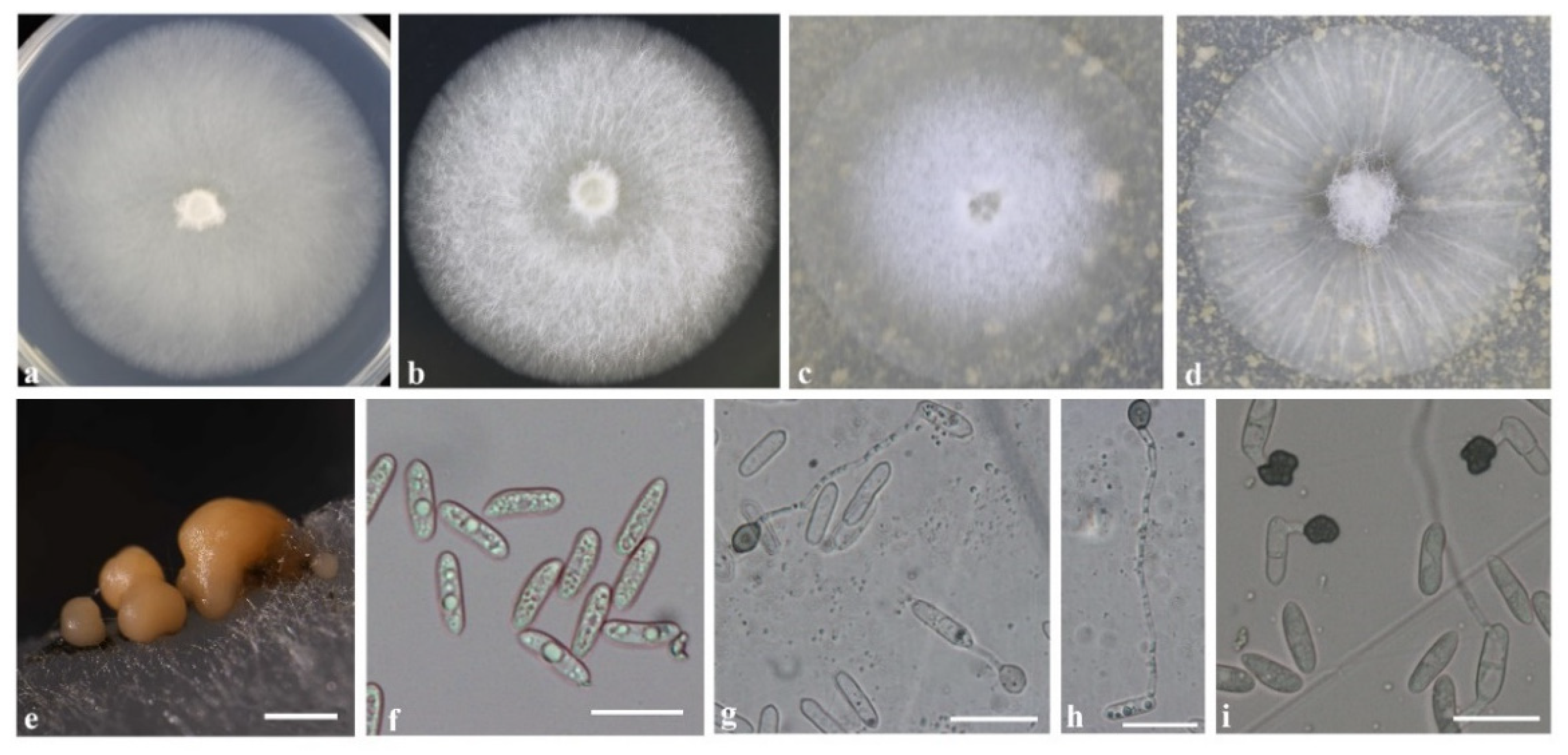
3.4. Morphology Description
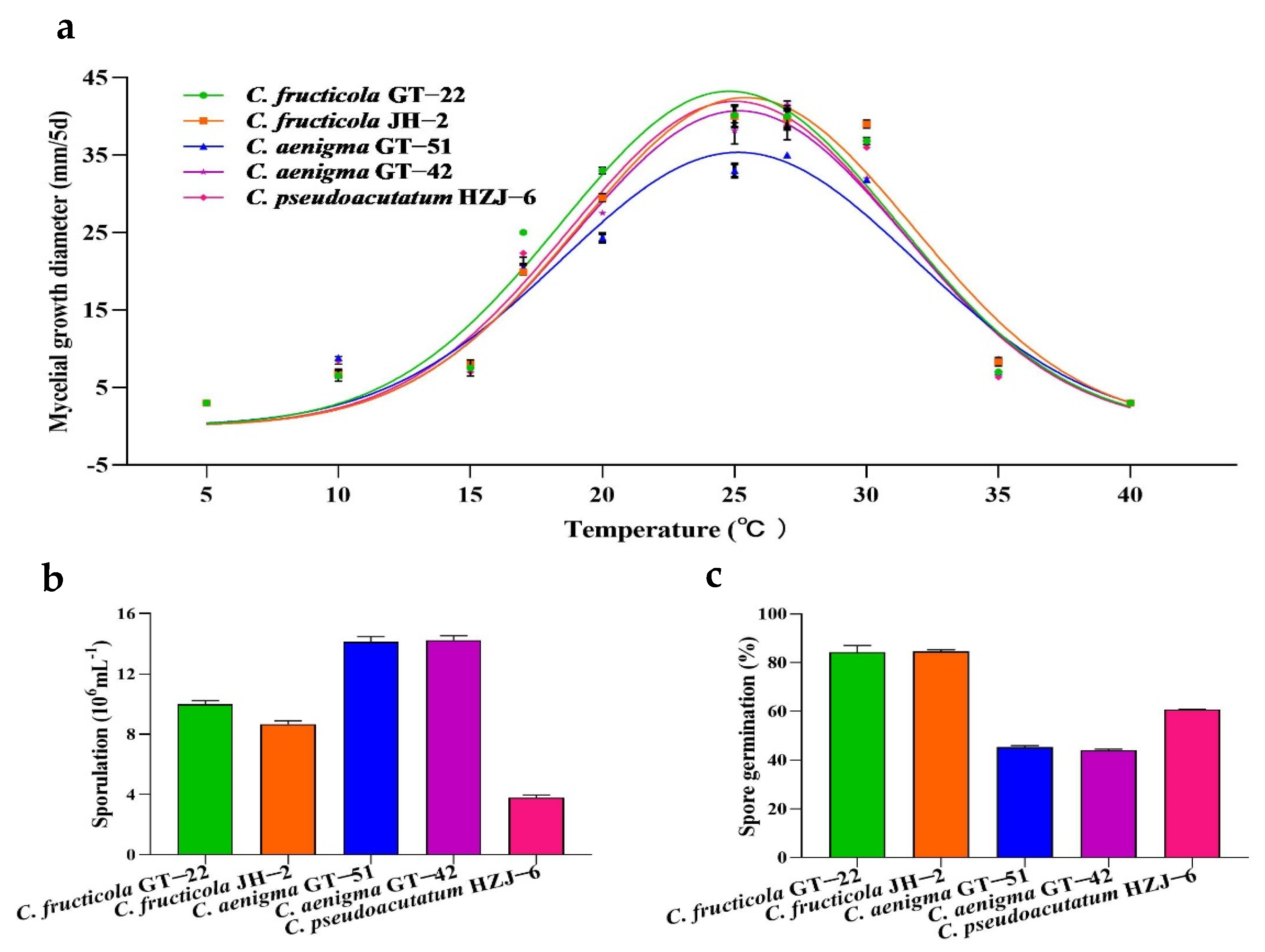
3.5. Mycelial Growth, Sporulation and Conidial Germination of Colletotrichum spp.
3.6. Pathogenicity Tests
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nassiri-Asl, M.; Hosseinzadeh, H. Review of the Pharmacological Effects of Vitis vinifera (Grape) and its Bioactive Constituents: An Update. Phytother. Res. 2016, 30, 1392–1403. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.-Q.; Deng, G.-F.; Guo, Y.-J.; Li, H.-B. Biological Activities of Polyphenols from Grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Li, Y.J.; Ma, X.Q.; Guo, J.; Liu, H.J.; Zheng, G.Q.; Tao, J.M. Development Situation and Prospects of Grape Industry in China. South China Fruits 2021, 50, 161–166. [Google Scholar]
- Whitelaw-Weckert, M.A.; Curtin, S.J.; Huang, R.; Steel, C.; Blanchard, C.; Roffey, P.E. Phylogenetic relationships and pathogenicity of Colletotrichum acutatum isolates from grape in subtropical Australia. Plant Pathol. 2007, 56, 448–463. [Google Scholar] [CrossRef]
- Shiraishi, M.; Koide, M.; Itamura, H.; Yamada, M.; Mitani, N.; Ueno, T.; Nakaune, R.; Nakano, M. Screening for Resistance to Ripe Rot Caused by Colletotrichum acutatum in Grape Germplasm. Vitis 2007, 46, 196–200. [Google Scholar]
- Ji, T.; Salotti, I.; Dong, C.; Li, M.; Rossi, V. Modeling the Effects of the Environment and the Host Plant on the Ripe Rot of Grapes, Caused by the Colletotrichum Species. Plants 2021, 10, 2288. [Google Scholar] [CrossRef]
- Meunier, M.; Steel, C.C. Effect of Colletotrichum acutatum Ripe Rot on the Composition and Sensory Attributes of Cabernet Sauvignon Grapes and Wine. Aus. J. Grape Wine Res. 2010, 15, 223–227. [Google Scholar] [CrossRef]
- Hong, S.-K.; Kim, W.-G.; Yun, H.-K.; Choi, K.-J. Morphological Variations, Genetic Diversity and Pathogenicity of Colletotrichum species Causing Grape Ripe Rot in Korea. Plant Pathol. J. 2008, 24, 269–278. [Google Scholar] [CrossRef]
- Yan, J.-Y.; Jayawardena, R.; Goonasekara, I.D.; Wang, Y.; Zhang, W.; Liu, M.; Huang, J.-B.; Wang, Z.-Y.; Shang, J.-J.; Peng, Y.-L.; et al. Diverse species of Colletotrichum associated with grapevine anthracnose in China. Fungal Divers. 2014, 71, 233–246. [Google Scholar] [CrossRef]
- Li, Z.; Santos, R.F.; Gao, L.; Chang, P.; Wang, X. Current status and future prospects of grapevine anthracnose caused by Elsinoe ampelina: An important disease in humid grape-growing regions. Mol. Plant Pathol. 2021, 22, 899–910. [Google Scholar] [CrossRef]
- Greer, L.; Harper, J.; Savocchia, S.; Samuelian, S.; Steel, C. Ripe rot of south-eastern Australian wine grapes is caused by two species of Colletotrichum: C. acutatum and C. gloeosporioides with differences in infection and fungicide sensitivity. Aust. J. Grape Wine Res. 2011, 17, 123–128. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Prusky, D. Pathogen quiescence in postharvest diseases. Annu. Rev. Phytopathol. 1996, 34, 413–434. [Google Scholar] [CrossRef]
- Oo, M.M.; Oh, S.-K. Identification and Characterization of New Record of Grape Ripe Rot Disease Caused by Colletotrichum viniferum in Korea. Mycobiology 2017, 45, 421–425. [Google Scholar] [CrossRef]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum Species Responsible for Anthracnose Diseases of Various Fruits. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Daykin, M.; Milholland, R.D.J.P. Ripe Rot of Muscadine Grape Caused by Colletotrichum gloeosporioides and Its Control. Phytopathology 1984, 74, 710–714. [Google Scholar]
- Salotti, I.; Ji, T.; Rossi, V. Temperature requirements of Colletotrichum spp. belonging to different clades. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Zulfiqar, M.; Brlansky, R.H.; Timmer, L.W. Infection of Flower and Vegetative Tissues of Citrus by Colletotrichum acutatum and C. gloeosporioides. Mycologia 1996, 88, 121. [Google Scholar] [CrossRef]
- Gunnell, P.S.; Gubler, W.D. Taxonomy and Morphology of Colletotrichum Species Pathogenic to Strawberry. Mycologia 1992, 84, 157–165. [Google Scholar] [CrossRef]
- Smith, B.J.; Black, L.L. Morphological, Cultural, and Pathogenic Variation among Colletotrichum Species Isolated from Strawberry. Plant Dis. 1990, 74, 69–76. [Google Scholar] [CrossRef]
- Dickman, M.B. Colletotrichum. In Fungal Pathology; Kronstad, J.W., Ed.; Springer: Dordrecht, The Netherlands, 2000; pp. 127–147. [Google Scholar]
- Chavhan, R.; Sable, S.; Narwade, A.; Hinge, V.; Kalbande, B.; Mukherjee, A.; Chakrabarty, P.; Kadam, U.S. Multiplex molecular marker-assisted analysis of significant pathogens of cotton (Gossypium sp.). Biocatal. Agric. Biotechnol. 2023, 47. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Hu, S.; Ye, B.; Hu, X.; Xiao, W.; Yu, H.; Zhang, C. Diversity and Resistance to Thiophanate-Methyl of Colletotrichum spp. in Strawberry Nursery and the Development of Rapid Detection Using LAMP Method. Agronomy 2022, 12, 2815. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum Species Associated with Anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef]
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74. [Google Scholar] [CrossRef]
- Chen, H.C.; Shen, B.W.; Wei, L.Z.; Fang, D.Y.; Wang, J.; Wu, J. Development Status and Countermeasures of Grape Industry in Pujiang. Sino-Overseas Grapevine Wine 2020, 4, 57–61. [Google Scholar]
- Arzanlou, M.; Bakhshi, M.; Karimi, K.; Torbati, M. Multigene phylogeny reveals three new records of Colletotrichum spp. and several new host records for the mycobiota of Iran. J. Plant Prot. Res. 2015, 55, 198–211. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ri-bosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Elsevier: Amsterdam, The Netherlands, 1990; Volume 31, pp. 315–322. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic rDNA ITS2 Types within a Monophyletic Lineage of the Fun-gusFusariumAre Nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.; Solon, S.L.; Crowhurst, R.N. Cloning and Molecular Characterization of the glyceralde-hyde-3-phosphate dehydrogenase-encoding gene and cDNA from the Plant Pathogenic Fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Chen, X.Y.; Dai, D.J.; Zhao, S.F.; Shen, Y.; Wang, H.D.; Zhang, C.Q. Genetic Diversity of Colletotrichum spp. Causing Strawberry Anthracnose in Zhejiang, China. Plant Dis. 2020, 104, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D. TreeView: An Application to Display Phylogenetic Trees on Personal Computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Karimi, K.; Arzanlou, M.; Pertot, I. Weeds as Potential Inoculum reservoir for Colletotrichum nymphaeae Causing Strawberry Anthracnose in Iran and Rep-PCR Fingerprinting as Useful Marker to Differentiate C. acutatum Complex on Strawberry. Front. Microbiol. 2019, 10, 129. [Google Scholar] [CrossRef]
- Dillard, H.R. Influence of Temperature, pH, Osmotic Potential, and Fungicide Sensitivity on Germination of Conidia and Growth from Sclerotia of Colletotrichum coccodes in vitro. Phytopathology 1988, 78, 1357–1361. [Google Scholar] [CrossRef]
- Chung, P.C.; Wu, H.Y.; Wang, Y.W.; Ariyawansa, H.A.; Hu, H.P.; Hung, T.H.; Tzean, S.S.; Chung, C.L. Diversity and Patho-genicity of Colletotrichum Species Causing Strawberry Anthracnose in Taiwan and Description of a New Species, Colletotrichum miaoliense sp. nov. Sci. Rep. 2020, 10, 14664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, L.; Xu, X.; Zou, X.; Duan, K.; Gao, Q.H. Characterization and Fungicide Sensitivity of Colletotrichum Species Causing Strawberry Anthracnose in Eastern China. Plant Dis. 2020, 104, 1960–1968. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Tang, X.B.; Jayawardena, R.S.; Yan, J.Y.; Chen, Q.X. Identification and characterization of Colletotrichum species causing grape ripe rot in southern China. Mycosphere 2016, 7, 1177–1191. [Google Scholar] [CrossRef]
- Echeverrigaray, S.; Scariot, F.J.; Fontanella, G.; Favaron, F.; Sella, L.; Santos, M.C.; Schwambach, J.; Pedrotti, C.; Delamare, A.P.L. Colletotrichum species causing grape ripe rot disease in Vitis labrusca and V. vinifera varieties in the highlands of southern Brazil. Plant Pathol. 2020, 69, 1504–1512. [Google Scholar] [CrossRef]
- Kim, J.S.; Hassan, O.; Chang, T. First Report of Colletotrichum aenigma Causing Anthracnose of Grape in Korea. Plant Dis. 2021, 105, 2729. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Jayawardena, R.S.; Liu, Y.; Xing, Q.K.; Yan, J.Y.; Li, X.H. Molecular Identification of the Pathogen Causing Grape Anthracnose in Beijing. J. Plant Prot. 2018, 45, 393–394. [Google Scholar]
- Xu, Y.; Xiao, T.; Chu, S.P.; Liu, J.X.; Rui, D.M.; Yao, K.B.; Yang, J.H. The Diversity and Resistance to Benzimidazole Fungicides of the Colletotrichum spp. Casuing Grape Anthracnose in Jurong City of Jiangsu Province. J. Nanjing Agric. Univ. 2022, 45, 78–85. [Google Scholar]
- Peng, L.-J.; Sun, T.; Yang, Y.-L.; Cai, L.; Hyde, K.D.; Bahkali, A.; Liu, Z.-Y. Colletotrichum species on grape in Guizhou and Yunnan provinces, China. Mycoscience 2013, 54, 29–41. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Lou, J.; He, L.Q.; Wang, Q.D.; Chen, L.H.; Zhong, X.T.; Wu, C.F.; Zhang, L.Q.; Wang, Z.Q. First Report of Colletotrichum fructicola Causing Anthracnose on Cherry (Prunus avium) in China. Plant Dis. 2022, 106, 317. [Google Scholar] [CrossRef]
- Zakaria, L.; Sahak, S.; Zakaria, M.; Salleh, B. Characterisation of Colletotrichum Species Associated with Anthracnose of Banana. Trop. Life Sci. Res. 2009, 20, 119–125. [Google Scholar]
- Moral, J.; Agustí-Brisach, C.; Raya, M.C.; Jurado-Bello, J.; López-Moral, A.; Roca, L.F.; Chattaoui, M.; Rhouma, A.; Nigro, F.; Sergeeva, V.; et al. Diversity of Colletotrichum Species Associated with Olive Anthracnose Worldwide. J. Fungi 2021, 7, 741. [Google Scholar] [CrossRef] [PubMed]
- Southworth, E.A. Ripe Rot of Grapes and Apples. J. Mycol. 1891, 6, 164. [Google Scholar] [CrossRef]
- López-Moral, A.; Agustí-Brisach, C.; Lovera, M.; Arquero, O.; Trapero, A. Almond Anthracnose: Current Knowledge and Future Perspectives. Plants 2020, 9, 945. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kulshrestha, S. Colletotrichum gloeosporioides: An Anthracnose Causing Pathogen of Fruits and Vegetables. Biosci. Biotechnol. Res. Asia 2015, 12, 1233–1246. [Google Scholar] [CrossRef]
- Ngugi, H. Epidemiology of sorghum anthracnose (Colletotrichum sublineolum) and leaf blight (Exserohilum turcicum) in Kenya. Plant Pathol. 2000, 49, 129–140. [Google Scholar] [CrossRef]
- Lin, T.; Xu, X.F.; Dai, D.J.; Shi, H.J.; Wang, H.D.; Zhang, C.Q. Differentiation in development of benzimidazole resistance in Colletotrichum gloeosporioides complex populations from strawberry and grape hosts. Australas. Plant Pathol. 2016, 45, 241–249. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolates | Colony Morphology 1 | Conidia | Appressorium | ||
|---|---|---|---|---|---|---|
| PDA 2 | MEA 3 | OA 4 | Size 5 (μm) | Size 5 (μm) | ||
| C. fructicola | GT-22 | R, C, G | R, C, W | R, C, W(c) to T(m) | 13.7–18.2 × 4.3–5.9 (16.9 ± 0.5) × (4.7 ± 0.5) | 6.4–8.5 × 4.8–7.3 (7.5 ± 0.7) × (5.7 ± 0.8) |
| JH-2 | R, C, G | R, C, W | R, Ra, W | 13.1–18.6 × 4.2–5.9 (16.8 ± 0.5) × (4.9 ± 0.3) | 7.2–9.9 × 6.1–8.8 (8.0 ± 0.4) × (6.9 ± 0.5) | |
| C. aenigma | GT-51 | R, C, GG | R, C, W | R, C, W(c) to T(m) | 13.2–17.1 × 4.1–5.6 (14.1 ± 0.4) × (5.1 ± 0.3) | 6.2–9.6 × 5.2–7.6 (6.9 ± 0.9) × (5.9 ± 0.6) |
| GT-42 | R, C, GG | R, C, W | R, C, W(c) to T(m) | 13.1–17.8 × 3.8–5.7 (13.9 ± 0.5) × (4.5 ± 0.5) | 6.0–8.3 × 4.9–6.2 (6.8 ± 0.6) × (5.7 ± 0.5) | |
| C. pseudoacutatum | HZJ-6 | R, C, A(c) to G(m) | R, C, O(c) to W(m) | R, C, O(c) to W(m) | 14.2–18.9 × 4.9–7.2 (16.6 ± 0.5) × (5.6 ± 0.4) | 7.5–10.1 × 5.2–7.2 (8.7 ± 0.9) × (5.4 ± 0.8) |
| Species | Isolates | Sporulation 1 (×106 mL−1) | Spore Germination 1 (%) | Disease Index 1 |
|---|---|---|---|---|
| C. fructicola | GT-22 | 10.0 ± 0.2 b | 84.2 ± 2.8 a | 44.8 ± 5.7 a |
| JH-2 | 8.7 ± 0.2 c | 84.5 ± 0.8 a | 45.8 ± 5.9 a | |
| C. aenigma | GT-51 | 14.1 ± 0.4 a | 45.3 ±0.6 c | 14.7 ± 2.1 b |
| GT-42 | 14.2 ± 0.3 a | 44.0 ± 0.5 c | 14.0 ± 3.9 b | |
| C. pseudoacutatum | HZJ-6 | 3.8 ± 0.2 d | 60.7 ± 0.3 b | 28.0 ± 1.7 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, B.; Zhang, J.; Chen, X.; Xiao, W.; Wu, J.; Yu, H.; Zhang, C. Genetic Diversity of Colletotrichum spp. Causing Grape Anthracnose in Zhejiang, China. Agronomy 2023, 13, 952. https://doi.org/10.3390/agronomy13040952
Ye B, Zhang J, Chen X, Xiao W, Wu J, Yu H, Zhang C. Genetic Diversity of Colletotrichum spp. Causing Grape Anthracnose in Zhejiang, China. Agronomy. 2023; 13(4):952. https://doi.org/10.3390/agronomy13040952
Chicago/Turabian StyleYe, Boyang, Jingqun Zhang, Xiangyang Chen, Wenfei Xiao, Jianyan Wu, Hong Yu, and Chuanqing Zhang. 2023. "Genetic Diversity of Colletotrichum spp. Causing Grape Anthracnose in Zhejiang, China" Agronomy 13, no. 4: 952. https://doi.org/10.3390/agronomy13040952
APA StyleYe, B., Zhang, J., Chen, X., Xiao, W., Wu, J., Yu, H., & Zhang, C. (2023). Genetic Diversity of Colletotrichum spp. Causing Grape Anthracnose in Zhejiang, China. Agronomy, 13(4), 952. https://doi.org/10.3390/agronomy13040952