Abstract
The coupling cycles of soil carbon (C), nitrogen (N), and phosphorus (P) have a significant impact on biogeochemical processes and ecosystem services. For centuries, large areas of floodplain wetlands in China have been extensively reclaimed for agricultural purposes due to population growth. However, little is known about the evolution of soil C:N:P stoichiometry along a reclamation chronosequence, particularly across different land uses. In this study, we investigated the variations in soil C:N:P ratios with land use and time gradients along a reclamation chronosequence comprising c. 0, 60, 100, 280, 2000, and 3000 years. Land reclamation induced nutrient decoupling, as it facilitated C and N accumulation from biological processes but restricted P supply controlled by geochemical processes. Soil C and N sequestration reached a stable state after 2000 years, while P declined steadily from 60 years. Soil C/P and N/P increased significantly and were controlled by organic carbon (OC) and total nitrogen (TN), respectively, indicating that an increase in C and N could also promote P uptake. Soil C/N declined in the first 60 years and stabilized at a threshold of 10:1. Different land use patterns following reclamation resulted in distinct soil nutrient structures. Paddies retained more OC and TN but exhibited lower adsorption of total phosphorus (TP) compared to adjacent dryland, leading to significant differences in C/P and N/P between land uses. Based on the redundancy analysis and random forest model, soil OC and TN were mainly affected by the abundance of bacteria metabolizing cellulose, while metal oxides, including Fe2O3 and CaO, could best predict TP. Soil C/P and N/P were mainly driven by soil texture and rose significantly with the increasing proportion of clay particles. Our study suggests that as reclamation proceeds, more anthropogenic management is required to regulate potential nutrient imbalances in order to prevent adverse effects on crop growth, soil quality, and ecosystem health. Additionally, any fertilization strategy should be developed based on dryland C and N deficiencies, and lack of P in paddies.
1. Introduction
Carbon, nitrogen, and phosphorus are three fundamental elements with maximal accumulations in plants and soil [1], and their biogeochemical cycle is crucial for preserving soil quality and avoiding excessive nutrient build-up [2,3]. Soil nutrient stoichiometry is a well-established theory for examining the coupling relationship between various nutrients, identifying soil-limiting nutrients, and formulating optimal nutrient management strategies [1,4]. Among these, soil C:N, C:P, and N:P ratios are considered effective indicators for assessing organic matter quality [5,6], phosphorus mineralization capacity [4,7], and nutrient constraint status [8,9]. Although soil C-N-P stoichiometric ratios are generally stable [10], many have hypothesized that they will vary dramatically under a changing environment, as factors such as soil acidity, mineral weathering extent, and biological activity can undergo substantial changes during this process [11,12,13].
Globally, an increasing amount of natural wetland has been converted for agricultural production due to the conflict between limited land resources and a growing population [14,15]. Knowledge of soil temporal dynamics and pedogenic evolution is essential for the sustainable use of these valuable soil resources [16]. Generally, as the reclamation time increases, soil nutrients tend to improve. For example, Zhang et al. [14] observed that soil OC and TN increased significantly over time in a coastal reclaimed zone. However, within the same transect, Yin et al. [17] demonstrated that long-term cultivation had a negative effect on TP reserves and increased the risk of leaching. Therefore, soil evolution exhibited a joint result of both biological and geochemical processes, during which bio-elements (C and N) tend to accumulate [13], while mineral elements (P) derived from geochemical processes may gradually run off [18]. Whether this opposite process will lead to a decoupling of soil C, N, and P cycles and if this decoupling will become more significant with increasing reclamation time remains largely unknown.
In addition to the impacts of reclamation time on soil nutrients, land use could also exert a substantial effect on soil redox conditions, plant organisms, and weathering intensity, which indirectly affect the retention of soil colloids to different nutrients and eventually alter the soil C-N-P ratios [19,20]. Therefore, numerous studies have been conducted to determine the effects of land use patterns on soil nutrient variations [11]. For instance, natural vegetation land (such as woodland and grassland) typically has higher levels of C and N but lower P than long-term-cultivated land [1,21]; nevertheless, the opposite has also been noted [22]. Additionally, the impact of land use may vary during different reclamation stages. For example, Roth et al. [23] observed that paddies accumulated more nutrients than non-paddy soil in the initial period of reclamation, whereas this disparity became negligible as the soil matured. Similarly, Zhang et al. [14] discovered that in the reclamation area of Cixi, as the reclamation proceeded, the SOC content in paddy fields increased continuously, but there was no significant change in dryland. Thus, reclamation time and land use patterns may interactively affect soil nutrient structure. However, information on this interaction remains limited, particularly at time scales spanning from centuries to millennia.
In a changing environment, alterations in edaphic properties (i.e., physical, chemical, and biological properties) can have a significant impact on the soil nutrient cycle [24,25,26]. However, prior studies on the impact of soil properties on OC, TN, TP, and their stoichiometries have primarily focused on regular drivers, such as soil pH, moisture, and texture, and have rarely considered the effect of major elements and biological indicators [27,28]. Soil major elements, such as Ca-, Fe-, and Al- oxides, are indicative of the extent of soil weathering [29], while biological indicators, such as soil bacterial functional genes, can reveal the biological activity of the soil [30,31]. Therefore, from the eco-stoichiometric perspective, it is essential to comprehensively and systematically compare the effects of both geochemical processes and biological processes on the content and ratio of OC, TN, and TP.
Eastern China is uniquely suited for wetland reclamation due to the presence of the Yangtze River, which continually transports sediment to its banks [14]. The Yangtze River, with a total length of approximately 6300 km, is the longest river in China. It flows from west to east, spanning 11 provinces including Qinghai, Hubei, and Anhui [32]. Since the 1950s, over 22,000 km2 of land has been reclaimed in this region [33]. Considering that more than 40% of the Chinese population live there but only 9% of the land is currently being cultivated [34], the trend in wetland utilization will continue into the foreseeable future. Our study is unique, as we constructed a soil chronosequence under fairly consistent climate, parent material, and terrain conditions and then collected samples from different land uses in each reclamation stage. This isolates time and land use variables from other soil impact factors, thereby identifying whether variations in soil nutrients result from reclamation time or land use pattern. Our purpose was to investigate the variation of soil C:N:P ratio over a reclamation chronosequence of c. 0, 60, 100, 280, 2000, and 3000 years under different land uses. Specially, we addressed the following questions: (i) Compared to soil C and N, does P show a different evolution along the reclamation time series? (ii) How do different land uses and human management practices influence the effects of reclamation years on soil nutrients? (iii) Which edaphic variables play a leading role on nutrients variation? We hypothesized that (i) prolonged reclamation time is expected to increase the content of OC and TN but reduce the TP reserve; (ii) at identical reclamation years, soil nutrients could be restructured across different land uses; and (iii) geochemical variables are correlated with the content of TP, while biological variables dominate the variations of OC and TN.
2. Materials and Methods
2.1. Study Sites
The current study was conducted on a scroll plain located in a meandering section of the Yangtze River, situated in eastern China’s Anhui Province (31.08°–31.27° N, 117.90°–118.01° E) (Figure 1). Over centuries, southward lateral accretion of point bars has constantly created fertile new land in the area. In order to render the new land suitable for agricultural purposes, a dozen levees were successively built during point bar expansion to contain Yangtze River floods. The levees built at different stages have divided the area into land tracts with distinct cultivation histories.

Figure 1.
Geographic location of the study area and sampling sites.
The newly formed point bar outside the latest-built levee, which is still receiving recent sediment deposition from the Yangtze River, represents the original soil parent material (time zero). The time of reclamation for various tracts of land was estimated using two different methods. For the particularly young land (i.e., 60, 160, and 280 years), the reclamation time was estimated based on the historical records of levee construction and the Yangtze River channel displacement [35,36]. For the two oldest sites (i.e., 2000 and 3000 years), optically stimulated luminescence (OSL) dating was used to determine the reclamation duration. This technique can accurately date fossils in geological sediments through ionized radiation [37]. Samples were collected from a depth of 60 cm and strictly shielded from light until analyzed. The OSL measurement procedure is detailed in Figure S1. All analyses were carried out at the Luminescence Dating Laboratory of Geomorphic Process team at the School of Geography and Ocean Science, Nanjing University.
Covering an area of about 160 km2, the flat terrain of the study area is suitable for farming. The climate is characterized as subtropical monsoon, with an average annual temperature of 15.8 °C and annual precipitation of around 1170 mm. Normally, newly reclaimed land is used for wood plantations (poplar, Populus euramevicana) and grassland (Eleusine indica). Dryland farming (wheat, Triticum aestivum; rape, Brassica campestris) was initial introduced in the 60-year reclaimed area and subsequently cultivated in later stages. Over time, with the expansion of cultivation and substantial improvement in the water-holding capacity of the subsoil layers, paddy–dryland rotation (rice–wheat and rice–rape) has become increasingly popular. On land older than 280 years, samples were only collected from dryland and paddies, as the reclaimed field beyond this stage had already been fully utilized for agricultural production and lacked continuous natural vegetation.
The levels of fertilization remain nearly identical across different reclamation stages within the same land use, as verified by local farmers and our field observations. This enables us to study the impact of reclamation duration on soil properties after eliminating the interference caused by fertilization. Specifically, until the 1970s, chemical fertilizers were rarely applied in the study area, as for the whole country. Instead, various organic fertilizers such as compost, cooking ash, as well as animal and human waste, were recycled into farmland to address nutrient deficiencies. Since then, chemical fertilizers have been increasingly consumed. Presently, the majority of nutrient inputs come from mineral sources, typically, 156 kg N ha−1, 90 kg P2O5 ha−1, and 146 kg K2O ha−1, respectively, in the wheat/rapeseed season, and 156 kg N ha−1, 60 kg P2O5 ha−1, and 146 kg K2O ha−1, respectively, in the rice season.
2.2. Field Sampling and Laboratory Analyses
A total of 90 soil samples were collected in January 2021, and the location of each site was recorded using a global positioning system (GPS: Trex Venture, Garmin, Olathe, Kansas, USA). At each sampling location, five surface subsamples (0–20 cm) within a 10 m radius were collected and bulked into a representative sample. The composite samples were air-dried at room temperature (20–25 °C) for 30 days and subsequently sieved using 2 and 0.2 mm meshes, respectively, before undergoing physical and chemical analysis.
Soil OC content was measured by the Walkley–Black wet oxidation method [38], TN was determined with the Kjeldahl method after digestion with H2SO4 [39], and TP was digested by HClO4-H2SO4 mixture and measured by the molybdenum blue colorimetry method [40]. C, N, and P content units were recorded as g/kg, and their ratios were mass-based. To determine soil moisture, part of the fresh soil was dried for 24 h at 105 °C [41]. Soil pH was measured with a glass electrode pH meter at a ratio of 1:2.5 soil to water. Electrical conductivity (EC) was determined with a conductivity meter (ddsj-308, Shanghai, China) at a soil-to-water ratio of 1:5. Soil texture was divided into three fractions (sand, >0.02 mm; silt, 0.002–0.02 mm; clay, <0.002 mm) according to the international standard of soil particle-size fractionation. Specifically, 5% Na hexametaphosphate solution was used as dispersant and determined by the sedimentation method. The elemental contents of CaO, Fe2O3, and Al2O3 were measured using X-ray fluorescence spectrometry (XFS) after grinding 5 g of each air-dried sample to <200-mesh in an agate mortar and compressing it into a disk [42].
Bacteria were the most abundant soil microbiota, and therefore, we took the abundance of bacterial functional genes as the soil biological indicator. To obtain bacterial OTUs, we first extracted DNA, amplified it, and sequenced the 16S rRNA gene, following the detailed procedure described in Tian et al. [43] and Jiang et al. [44]. Next, we matched the bacterial OTUs with the FAPROTAX dataset (script version 1.1) to identify putative function groups. The FAPROTAX dataset is a database specifically developed to predict nutrient cycling (i.e., C, N, P, and S) and pathogenic genes in soil [44]. After obtaining all of the functional groups, we refined our selection to three groups that are relevant to nutrient cycling and also have the highest abundance, namely chemoheterotrophy, nitrogen fixation, and cellulolysis. For more information about the experimental design and sample properties, please refer to Tables S1 and S2.
2.3. Statistical Analysis
The effects of reclamation duration and land use on soil properties were analyzed using analysis of variance (ANOVA). A two-way ANOVA was conducted to assess the impact of reclamation duration, land use, and their interaction. To determine the differences among reclamation stages and land use patterns, one-way ANOVA was employed with a least significant difference (LSD) test at the p < 0.05 level of significance. The correlation between soil C, N, and P contents, as well as their ratios, was analyzed using ordinary least squares (OLS) regression analysis.
The multicollinearity between environmental factors was assessed using the variance inflation factor (VIF), and parameters with high levels of multicollinearity were eliminated for further analysis. The redundancy analysis (RDA) and random forest (RF) model were combined to investigate the impact of environmental factors on soil OC, TN, TP, and their ratios. RDA provides an intuitive representation of the directional relationship between soil properties and nutrients, while RF allows for the quantitative assessment of the impact of specific indicators. RDA was conducted using CANOCO 5.0 (Microcomputer Power, Ithaca, NY, USA) software [45] with forward selection of explanatory variables. RF is an ensemble machine-learning method based on decision trees [46]. Three hyperparameters are chosen during the forest-building process: the number of trees (ntree), variables tried at each split (mtry), and the minimal terminal node size (node size). Hyperparameter tuning is a crucial step in RF, where different combinations of hyperparameters are tested and evaluated using cross-validation. Cross-validation involves dividing the data into multiple folds, where one part is for training and another part for testing. The performance of the model is then evaluated, and the hyperparameters with the best performance are selected [46,47,48]. For this study, the hyperparameters were set to 500, 3, and 5, respectively. The forest-building process was conducted using the R package Random Forest (R Development Core Team 2019, Vienna, Austria).
3. Results
3.1. Dynamics of Soil Environmental Factors for Different Land Uses along the Chronosequence
The two-way ANOVA showed reclamation duration and land use patterns had a significant effect on the investigated environmental factors (p < 0.05) (Table S3), and their interactions could also exert significant effect (p < 0.05). Therefore, we further analyzed the variations of soil properties by controlling reclamation duration or land use.
The reclamation duration primarily affects the following soil properties: pH, EC, texture, biological indicators, and CaO. Without the influence of land use, soil pH, EC, and CaO generally exhibited a gradual decline with increasing reclamation time (excluding y60) (Figure 2). A significant increase of clay content in dryland and paddies began after an initial slight decrease over the first 60 years, which was in contrast to the trend of sand particles. All of the biological indicators in woodland showed a continuous increase, while those in dryland and paddies exhibited a slight decrease after 2000 years.

Figure 2.
Evolutionary trends in soil environmental factors along the chronosequence in different land uses. (a) pH; (b) electrical conductivity; (c) moisture; (d) clay; (e) silt; (f) sand; (g) chemoheterotrophy; (h) nitrogen fixation; (i) cellulolysis; (j) Fe2O3; (k) CaCO3; and (l) Al2O3. Different uppercase letters indicate that means are significantly different (p < 0.05) among land uses at the same reclamation stage. Different lowercase letters indicate means are significantly different (p < 0.05) among the same land use across reclamation stages.
Land use practices had a significant impact on all types of soil properties (p < 0.05). Grassland and woodland showed a higher level of finer particles (clay and silt), functional genes (chemoheterotrophy, nitrogen fixation, and cellulolysis), and Fe- and Al-oxides but lower pH, sand, and CaO when compared to the sediment. At each stage, dryland had a higher level of sand, nitrogen fixation (excluding y280), cellulolysis, and all the oxides. However, it displayed lower pH (excluding y160), EC, moisture, and clay particles when compared to paddies.
3.2. Soil Nutrient Contents and Ratio Dynamics for Different Land Uses along the Chronosequence
Soil OC, TN, TP, C/N, C/P, and N/P in reclaimed land showed obvious differences with sediment and changed significantly over time (Figure 3). Specifically, OC and TN increased from 11.0 g kg−1 and 0.85 g kg−1 (sediment) to 20.1 g kg−1 and 1.74 g kg−1 (y2000) and then decreased to 16.3 g kg−1 and 1.51 g kg−1 (y3000), respectively. After reclamation, the content of TP in sediment increased from 0.70 g kg−1 to 0.99 g kg−1 (y60) and then constantly decreased to 0.75 g kg−1 (y3000). Soil C/N decreased rapidly from 12.1 (sediment) to 10.2 (y60) and then fluctuated slightly over time. When the land was reclaimed for around 60 years, soil C/P decreased rapidly from 15.8 to 13.0 but gradually increased to 24.1 (y2000) and fluctuated slightly thereafter. Soil N/P showed a significant increase with reclamation time (p < 0.05), rising from 1.21 (sediment) to 2.10 (y2000), but subsequently, it showed no significant difference compared to 2.06 (y3000) (p > 0.05).

Figure 3.
OC, TN, and TP contents and ratios at different stages of land reclamation. (a) Organic carbon; (b) C/N ratio; (c) total nitrogen; (d) C/P ratio; (e) total phosphorus; (f) N/P ratio. Control: fresh sediment. Lines within the boxes give the median, boxes the 25th and 75th percentile, and whiskers the lowest and highest values excluding outliers. The dotted lines are the means (square symbols) connection of each reclamation stage. Different lowercase letters indicate means are different at p < 0.05 over different reclamation stages. The yellow-shaded arrows represent our interpretation on nutrient content changes and description of the primary dynamics of ratios.
Compared to sediment, woodland and grassland have significantly higher levels of OC and TN (p < 0.05), with increases ranging from 60.98 to 67.24% and 77.37 to 102.46%, respectively (Figure 4). Additionally, the OC and TN contents in paddies and woodland were significantly higher than that in dryland at each reclamation stage (p < 0.05) (except for a slightly lower OC in y3000). The TP content in woodland and grassland also increased by 18.03–22.18% when compared to sediment. In contrast to OC and TN, the TP content was found to be in the order of dryland > paddies > woodland.

Figure 4.
OC, TN, and TP contents and ratios along the chronosequence over different land uses. (a) Organic carbon; (b) C/N ratio; (c) total nitrogen; (d) C/P ratio; (e) total phosphorus; and (f) N/P ratio. Different uppercase letters indicate that means are different at p < 0.05 among land uses at the same reclamation stage. Different lowercase letters indicate means are different among the same land use across reclamation stages.
The C/N in woodland and grassland decreased slightly from 13.41 (sediment) to 10.69 and 11.7, respectively. However, there was no significant change in soil C/N with increasing reclamation time in different land uses (p > 0.05). The impact of land use on soil C/P and N/P remained consistent across different stages. When sediment was converted into woodland and grassland, soil C/P increased by 36.62–36.83%, and N/P increased by 51.07–66.39%. C/P and N/P in paddies and woodland were significantly higher than in dryland at the same reclamation stage (p < 0.05) (excluding y3000).
The correlation between soil nutrient content and ratios, as well as various nutrients, was presented in Figure 5. The results indicated a significant negative correlation between soil C/N and TN (p < 0.05), while no significant correlation was observed between soil C/N and OC (p > 0.05). In addition, soil C/P and N/P were significantly and positively correlated with OC and TN (p < 0.01), respectively, with stronger correlations observed than with TP. Furthermore, a significant positive correlation was found between OC and TN (p < 0.01), while no significant correlation was observed between either OC or TN with TP (p > 0.05).
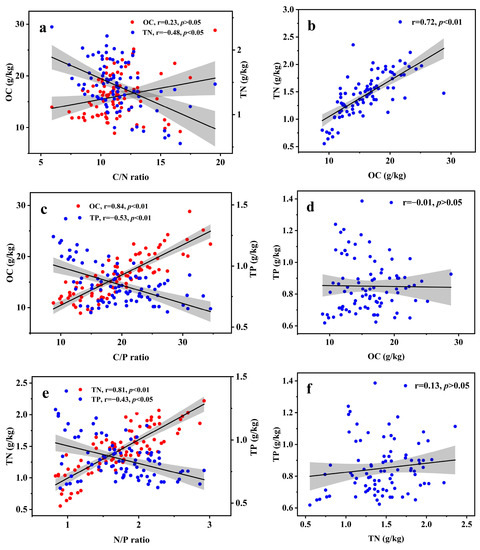
Figure 5.
Linear correlations between soil nutrients and eco-stoichiometric ratios (a–c) and interaction relationships of various nutrients (d–f). OC, organic carbon; TN, total nitrogen; TP, total phosphorus.
3.3. Identification of Primary Environmental Drivers Affecting Soil C:N:P Stoichiometry
Nine soil variables, namely reclamation duration (RD), pH, EC, clay, silt, Fe2O3, CaO, cellulolysis, and nitrogen fixation were selected based on VIF and Monte Carlo permutations (Table S4). Additionally, land use was also considered as a variable for subsequent calculation. The effects of soil properties on the structures of soil nutrients were evaluated using RDA ordination plots across different land uses, with the first two axes accounting for approximately 54% of the total variation (Figure 6). According to the vectors, the variation of OC and TN was found to be correlated with the abundance of microbial functional groups, including cellulolysis and nitrogen fixation, whereas TP was primarily determined by the presence of metal oxides (i.e., Fe2O3 and CaO) within the soil. The nutrient ratios, excluding C/N, were predominantly influenced by soil texture and showed a positive association with the percentage of fine particles, including clay and silt. Additionally, land use types strongly altered the composition of soil nutrients and formed distinct groups. The replicated samples collected from each reclaimed land were closely grouped and were distinctly separated from the sediment samples. The soil nutrients structure of paddies and dryland were clearly distinguished along the first axis, with paddies showing higher levels of OC, TN, C/P, and N/P, while dryland had higher TP. Notably, with increasing reclamation years, paddies potentially showed an increasing distinction from other groups.
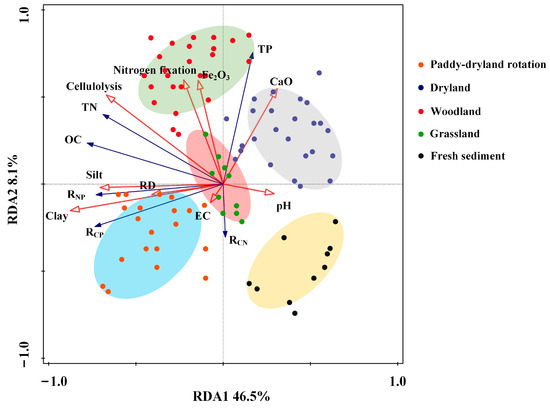
Figure 6.
Redundancy analysis (RDA) of soil nutrients stoichiometry with environmental variables. The significance of the RDA analysis results was tested by a Monte Carlo permutation test. OC, organic carbon; TN, total nitrogen; TP, total phosphorus; EC, electrical conductivity; RD, reclamation duration.
The RF model indicated that the abundance of the cellulolysis gene was the primary factor influencing both OC and TN (Figure 7). The contents of Fe2O3 and CaO were most significantly correlated with the soil TP. The proportion of clay particles was the primary factor explaining the variation in C/P and N/P. Although EC was identified as the most important factor influencing C/N, none of the investigated variables (including EC) had a significant impact on C/N.
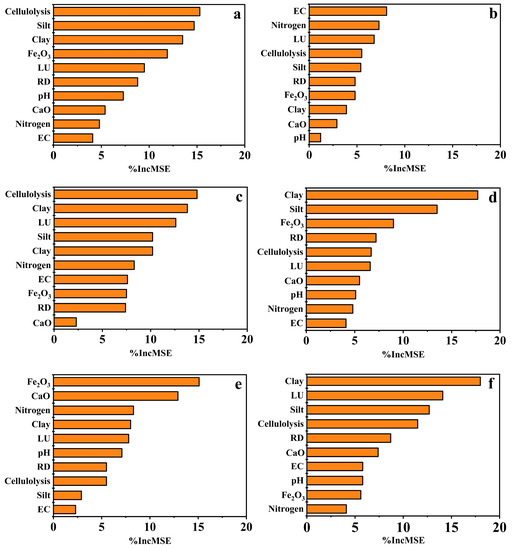
Figure 7.
Relative importance of each predictor calculated by the random forest model for each nutrient index: (a) organic carbon; (b) C/N ratio; (c) total nitrogen; (d) C/P ratio; (e) total phosphorus; and (f) N/P ratio. EC, Electrical conductivity; LU, land use; RD, reclamation duration; nitrogen, nitrogen fixation; % incMSE, increase in mean squared error.
4. Discussion
4.1. Effects of Reclamation Duration on Soil C, N, and P Contents and Stoichiometric Ratios
Based on the chronosequence, soil nutrient structure underwent significant changes over the course of millennia of reclamation. We hypothesized (i) that soil OC and TN contents would increase significantly over time, which was confirmed by their dynamics during the first 2000 years (Figure 3). The coupling variation between soil C, N, and the time factor can be ascribed to increased agricultural intensity during this period. After reclamation, the application of organic and chemical fertilizers could either directly introduce C and N into agroecosystems or indirectly promote biomass production to provide soil fertility [49,50]. Additionally, continuously improved soil properties (e.g., dealkalization and improved drainage) boosted soil biological activity, which plays a key role in regulating the soil nutrient cycle [14]. However, the OC and TN content in y3000 were lower than those in the y2000 site, providing evidence of mature soil degradation; this is a pervasive problem for reclaimed wetlands worldwide [12]. For example, Huang et al. [12] and Roth et al. [23] reported that the soil OC and TN contents in the Cixi reclamation area increased continuously during the first 300 years, but both subsequently showed significant decreases. The primary reason for fertility degradation in the oldest soil is due to its high acidity, resulting from the long-term soil weathering and the accumulation of acidic fertilizers [29]. Strong soil acidity could diminish soil fertility through the following mechanisms: First, the most important effect is its hindrance of microbial activity (Figure 2), which slows down critical processes related to the turnover of C and N. This includes the decomposition of external organisms into humus, atmospheric N fixation, and nitrification [51]. Moreover, microorganisms can secrete adhesives, promote organic matter aggregation, and increase soil colloids, thereby reducing the decomposition rate of organic matter. Second, soil acidification could decrease the concentration of Ca2+ in soil (Table S5), which is essential for maintaining soil fertility [52]. Ca2+ can bind with negative charges borne by soil particles and diminish the electrostatic repulsion between organic matter and negative charges. As a result, this makes it easier for organic matter to bind with soil particles. To alleviate soil degradation caused by long-term cultivation, various management practices can be implemented. One effective method is to apply an appropriate amount of lime to older farmland. This approach can help to reduce soil acidity and provide much-needed Ca2+ supplementation. Additionally, other practices, such as fallowing, crop rotation, and increasing the application of organic fertilizers, can also be highly beneficial in restoring and maintaining soil health [53]. Overall, our observations of C and N dynamics support the long-held notion that soil development results jointly from progressive (improvement) and regressive (degradation) processes [54]. The relative strength of these two opposing processes will determine whether evolution is progressing, stable, or regressing [55].
The dynamics of TP following reclamation were partially consistent with our original hypothesis (i) that prolonging reclamation time will reduce the TP reserve, as it exhibited two distinct stages during the 3000 yr time sequence (Figure 3). In the first 60 years, there was an increasing trend. This could be attributed to our original hypothesis based on previous chronosequence experiments conducted in natural ecosystems [10]. However, in agricultural land, the early increase in TP could be a transient response resulting from the external addition of P fertilizer [14]. After 60 years, a steady reduction in TP was observed, which was consistent with Walker and Syer’s model [56], indicating that as soil matured, the reserve and availability of P decreased constantly. Given that cultivation management in the older land was nearly identical to the younger sites, the decline in P during the later stage may be attributed to leaching losses over long-term reclamation [57]. In China, it is estimated that P leaching through runoff is 1.57 × 106 t in 2012, which is equivalent to 0.63 g per m2 [14]. Due to higher temperature and precipitation in our study area than other regions in China, P leaching is likely to be greater than the average.
Soil C-N-P stoichiometry could characterize C-, N-, and P-coupled cycles and connect different aspects of ecosystem functionality [58]. C/N serves as an effective indicator reflecting soil organic matter decomposition and C loss [59]. In our study, sediment C/N was significantly higher than that of reclaimed soil (p < 0.05), which is due to the fact that labile SOM in sediment was largely removed by river transportation, leaving behind recalcitrant substrates that typically have a higher C/N. After reclamation, fresh organics (e.g., plant stubble and manure) were incorporated into the soil, thereby causing rapid microbe growth to decompose organisms and mineralize organic nutrients (e.g., N and P). Soil microorganisms generally require a diet with the C/N of 24:1 to survive, with eight parts of C used for body maintenance and sixteen for energy [60]. Obviously, microbial decomposition consumed more C resources, leading to a reduction in the C/N of the remaining humus. Therefore, the decrease in soil C/N after reclamation indicated an improvement in soil biological activity and SOM quality, which was considered an important feature of initial soil development [59]. After 60 years, soil C/N remained well-constrained at each stage and ultimately stabilized at 10:1, which was similar to the average chemical composition of microbial cells, indicating that microorganisms may play a dominate role in regulating soil nutrient cycles [61].
C/P can be used to evaluate mineralization potential and P availability (4). In this study, soil C/P increased by 85% from 60 to 2000 years (Figure 3). During the earlier stages of reclamation, a rapid prevalence of biological control on nutrient cycling indicated that C and N were made available; meanwhile, P was depleted by parent material weathering, leading to a disparity in the C, N, and P cycles [54]. N/P can be used as an indicator to investigate soil-limiting nutrients [8]. In our study, N/P increased steadily over time, up to 2000 years. This indicated that land reclamation caused a decoupling between soil N and P, and crop growth and microbial decomposition were increasingly limited by P. Overall, the observed changes in soil C/P and N/P indicated reclamation disrupted the C, N, and P cycles in a nonlinear manner and would have a negative impact on the biogeochemical reactions controlling key ecosystem functions (i.e., primary production and material cycling) and services (i.e., food production and carbon sequestration) [13].
Notably, C/P and N/P were significantly controlled by OC and TN, respectively (p < 0.01) (Figure 5b,c). Therefore, although there was a weak correlation between TP and OC or TN (Figure 5e,f), the positive effect of elevated C and N addition on plant growth could stimulate soil P uptake [62,63]. With the increasing use of N fertilizers after reclamation, soil acidity was induced, leading to a decrease in the rate of SOM decomposition and favoring soil C accumulation [54]. Moreover, N additions could promote organisms to produce more extracellular phosphatase enzymes that cleave ester-P bonds in soil organic matter, thereby increasing P availability for crop uptake [64,65]. Although farmers tend to apply chemical P fertilizers, these compounds are less mobile and are poorly utilized during the crop season (less than 30% of total fertilization) [9]. Consequently, compared to accessible C and N, a slower rate of P increase can eventually lead to undesirable increases in C/P and N/P and limit crop growth due to soil P deficiency [13,66]. Similarly, Vitousek et al. [65] proposed that transactional P limitation (one type of P limitation) occurs generally when other resources (especially N) are adequate; even though P exists in primary minerals, its supply via weathering cannot keep up.
4.2. Effects of Land Use Pattern on Soil C, N, and P Contents and Stoichiometric Ratios
Land use patterns have been shown to affect soil nutrient variability by directly altering soil properties and influencing biological processes in the rhizosphere [67]. In unreclaimed land (y0), soil nutrient contents in grassland and woodland were significantly higher than in sediment (p < 0.05) (Figure 4). This can be attributed to the fact that natural vegetation growth provides more organic residues (i.e., dead branches and leaves) to the surface soil [26], which activates soil microbial function (e.g., cellulolysis process and nitrogen fixation) and ultimately accelerates the transformation of nutrients from organic to inorganic forms [68].
Our hypothesis (ii) was that land use patterns would significantly impact nutrient content, and this was confirmed by the finding that dryland had significantly lower OC and TN content compared to paddies at the same reclamation stage (p < 0.05) (Figure 4). Several factors may contribute to the inferiority of dryland in terms of sequestering OC and TN. Firstly, paddies receive a higher input of SOM (approximately 60–70%) compared to dryland [69], as most of the biomass produced in dryland is removed, and only a small proportion is returned to the surface soil. The amount of plant residues returned to the soil is generally positively correlated with OC accumulation [14]. Secondly, OC in the surface soil is sensitive to agricultural tillage practices, and seasonal tillage in dryland tends to destabilize soil macroaggregates, disrupt topsoil structure, and thus accelerate OC and N loss [70,71]. Lastly, the relatively lower levels of OC and TN in dryland soil may be attributed to faster decomposition rates of organic matter in well-aerated soils compared to waterlogged soils [23].
At each stage, it was observed that dryland had a higher TP and lower C/P and N/P than paddies, possibly due to the special practice of applying P fertilizer in southern China [72]. Local farmers tend to apply P fertilizer during the dryland farming season, when there was abundant soil ferric ion (Fe3+), which has a strong adsorption capacity for P ions. During the summer season, the fields are rotated to paddies, where the Fe3+ is reduced to ferrous iron (Fe2+), and the adsorbed P is massively released to facilitate rice growth [73]. While this fertilization approach has the potential to improve fertilizer utilization and minimize nutrient loss, it may also lead to an uneven distribution of P among different land uses. Moreover, the frequent flooding and drainage process in paddies can wash away various soil substances, including metal oxides, clays, and CaCO3, which are crucial soil P sorbents [50]. Previous studies have consistently demonstrated that land use patterns can significantly impact soil C, N, and P contents and stoichiometric ratios. For example, as reported by Zhang et al. [14], in the tidal flat reclamation area of Dafeng, paddy fields were superior to drylands in enriching C and N. Similarly, Zheng et al. [1] conducted a continental-scale study comparing the effects of different land use patterns on soil C, N, and P stoichiometry and discovered that compared to the woodland and paddy fields, uplands are relatively limited by C and N but enriched with P. These findings are consistent with the results of the current study.
4.3. Evolution of Soil Properties and Their Regulation of C:N:P Stoichiometry
The investigated properties were significantly impacted by the reclamation time and land use pattern (p < 0.05) (Figure 2). Soil pH gradually decreased over time due to the leaching of base cations such as calcium (Ca2+), magnesium (Mg2+), and potassium (K+), which are weathered from minerals [74]. Decreasing soil pH could restrict the turnover rate of soil nutrients and reduce their availability. Additionally, it can hinder plant root growth and increase the occurrence of pests and diseases. The dramatic decline in EC following reclamation was primarily due to natural desalination and artificial irrigation practices [28].
At the identical stages, pH in paddy soil was significantly higher than that in dryland. Due to the significant correlation between CaCO3 and pH (r = 0.65) (Figure S2), a higher value in paddies was related to the large amounts of OH- generated from the hydrolysis of CaCO3 under flooded conditions. Additionally, the paddy soil had a higher EC compared to dryland soil. This was due to the rising groundwater level that occurs during long-term flooding and the accumulation of soluble salts in the topsoil after evaporation [75]. Long-term irrigation and improved water-holding capacity from higher fine particle content leads to a higher soil moisture in paddies. Conversely, frequent plowing and lower vegetation coverage in dryland accelerated moisture loss [14]. However, although these environmental factors were obviously altered by reclamation time or land use pattern, the variability in soil nutrient stoichiometry explained by these variables was rather lower than biological and weathering indicators and soil texture.
The abundance of soil bacterial functional genes, especially those involved in cellulolysis, has been identified as the primary regulator of OC and TN (Figure 7). In addition to organic fertilizers such as human and animal manure, crop residues also play a crucial role in the formation of soil organic matter in agricultural fields [76]. In the research area, it is customary to incorporate straw into both dryland and paddy fields once a year during the crop harvest season. Dryland fields are typically integrated between May and June, with an average amount of 9000 kg/hm2. As for paddy fields, the usual practice is to incorporate straw in September, with an average amount of 12,000 kg/hm2 [77]. This practice has been shown to be superior to traditional agricultural practices, as the decomposition of crop straw can not only enhance soil nutrient content but also promote soil aeration and water retention and reduce environmental pollution caused by pesticides and fertilizers [76]. Crop straw is rich in cellulose, with an average content of 30–40% in primary crops (e.g., rice, wheat, and maize), making it an ideal candidate for the cellulolysis process [76]. However, the transformation of straw to humus largely depends on microbial functional genes capable of breaking down cellulose. Additionally, the soil environmental status improves significantly after reclamation, which promotes the proliferation of soil microorganisms [28]. Together, the recovery of microbial activity and the addition of abundant substrate significantly improve soil C. Since over 90% of the N in the surface layer of most soils is in organic form [14], its concentrations and availability are closely related to soil organic matter and also affected by the cellulolysis process.
In contrast, the content of soil metal oxides, including CaO and Fe2O3, was the most predominant factor in explaining the variability of soil P (Figure 7). As a mineral element, P originates from mineral weathering; however, its storage after release mainly depends on the soil’s adsorption capacity. As the pedogenetic age extends and soil acidity increases, the adsorption of P shifts from Ca2+- to oxide-dominated; nevertheless, both of them were recognized as important P adsorbents in soil [54]. Ca ions could bond with P and cause P precipitation as insoluble Ca-P species, which directly retain P electrostatically through the exchange of HCO3− for H2PO4 [78]. The adsorption of Fe/Al oxides on P was mainly due to their large specific surface areas, which provided a large number of adsorption sites [79].
Soil texture is apt to change at landscape and regional scales with soil weathering, resulting in a complex interaction between soil particles and the C, N, and P cycles [80]. In our study, OC, TN, C/P, and N/P were all positively correlated with clay and silt content (Figure 6), which was consistent with the findings of Wang et al. [10]. Soils with higher amounts of finer particles typically contain more C and N, as these particles could chemically stabilize and physically protect organic matter [81]. However, the effect of soil texture on TP was not evident, which was unexpected, as finer particles in the soil were also listed as important P adsorbents [82]. This phenomenon may be related to the redistribution of TP content caused by land use patterns in this study. Although there was still a positive correlation between TP and clay content in natural soils (sediment and grassland) (r = 0.62), this correlation was greatly disrupted by anthropogenic management in agricultural land, where large amounts of P fertilizer were applied to dryland with lower clay content. Conversely, paddy fields with a higher proportion of clay were nearly unfertilized with P, which dramatically altered the relationship between soil texture and TP.
4.4. Imbalance Risk of Soil C, N, and P Induced by Land Reclamation and Land Use Patterns
Knowledge of the directions, rates, and thresholds of soil development after reclamation, as well as its influencing factors and mechanisms, are required for sustainable utilization of soil resources [56]. In the present study, it was observed that land reclamation decoupled the nutrient cycles, as it facilitated C and N accumulation associated with biological processes, but had an adverse impact on the reserves of P which was controlled by geochemical processes. It is worth noting that the content of C, N, and P all declined after 2000 years, indicating fertility degradation in ancient soil and nonlinear evolution of soil properties. Moreover, the study revealed a succession threshold of soil C/N (10:1) over the chronosequence, and this similar ratio to microbial cells may indicate the crucial role of microorganisms in mediating C and N cycles.
Dryland and paddy–dryland rotation are the two primary agricultural land uses in eastern China. However, our study has revealed distinct variations in the content of C, N, and P, as well as their stoichiometric ratios between these two land uses. In paddies, the elevated levels of C and N were likely attributed to the massive return of organisms and lower decomposition under submergence [14]. Nonetheless, inadequate application of P fertilizer and losses of P adsorbents together reduced its soil P reserves. Conversely, in dryland, the removal of plant residue and well-aerated conditions hindered C and N sequestration, but constant manuring and effective adsorption capacity tended to sustain a high level of P. The stark contrast in anthropogenic management and soil conditions between drylands and paddies results in significantly lower levels of C/P and N/P in the former. Thus, we deduced that soil C/P and N/P were likely the crucial factors in distinguishing between dryland and paddies in eastern China. Differences in soil nutrient structure between paddies and drylands could lead to different constraints on crop growth. For example, lower OC and TN in dryland might limit wheat chlorophyll and protein synthesis, while insufficient P supply in paddy fields could result in poor root growth and slow metabolism [83,84]. As the reclamation proceeded, the distribution of nutrients within farmland would become increasingly uneven, which would not only lead to a reduction of fertilizer utilization rates but also cause environmental pollution and damage to soil health [1,11].
4.5. Discontinuous Evolution of Soil Properties along the Soil Chronosequence
In an ideal scenario, soil properties would undergo a continuous and consistent evolution over time. However, this process is challenging to observe through empirical research and may be nonexistent [54]. In our study, we found notable variations in the properties of the y60 soil compared to the adjacent stages (y0 and y160). This discontinuous evolution along soil chronosequence has also been documented in previous studies [85], possibly attributable to the river sedimentation during a specific stage. The succession of soil particle observed in our study provided some degree of confirmation. Soil texture, an important indicator of depositional environment [86], was compositionally different in the y60 soil compared to other reclamation stages (Figure 2). Nevertheless, we observed that except for a slightly higher TP and lower C/P, soil nutrient dynamics in this stage still aligned with our hypothesis.
5. Conclusions
Our research has revealed an overall inverse succession between soil OC, TN, and TP during the process of land reclamation. Specifically, OC and TN increased from 11.0 g kg−1 and 0.85 g kg−1 to 20.1 g kg−1 and 1.74 g kg−1, respectively, while TP gradually decreased from 0.99 g kg−1 to 0.75 g kg−1. Soil C/P and N/P increased from 15.8 and 1.21 to 24.1 and 2.1, respectively. For the same stage, paddies stored more C and N but adsorbed less P than adjacent dryland, resulting in significant differences in soil C/P and N/P between land uses. Soil biological process mainly affected the content of C and N, while the concentration of Fe/Ca oxides dominated the reserve of P. The C/P and N/P exhibited a significant and positive correlation with the proportion of clay particles. Older soils have a significantly reduced adsorption capacity for P, which necessitates a moderate increase in the application of chemical P fertilizers to maintain a balance of soil C, N, and P, but it is crucial to avoid potential P loss and pollution risks. By analyzing the evolution of soil C:N:P stoichiometry, differences in nutrient structures between various reclamation stages and land use patterns could be revealed, providing a reference for dynamically adjusting farmland fertilization strategies and formulating land-use-oriented management practices. Although the present study found that reclamation duration and land use patterns could significantly drive the nutrient structure in soil, it remains unclear whether this will affect the overall nutrient characteristics within the ecosystem. Further analysis should comprehensively consider various sources of nutrients, such as plant tissues and microbial biomass. This study examines the evolutionary characteristics of soil nutrients along the reclamation gradients and the land use effect in the typical floodplain of the Yangtze River Basin. Its significant findings are expected to provide valuable guidance for the sustainable utilization of cultivated land resources and the preservation of agricultural productivity in comparable wetland reclamation areas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13041133/s1. Table S1: Species of plant/crop and sampling number at each land use over different reclamation duration. Table S2: Basic descriptive statistics for soil physical and chemical properties in the study area. Table S3: Two-way ANOVA for effects of reclamation duration and land use on soil properties. Table S4: Examination results by variance inflation factor (VIF). Table S5: CaCO3 contents (g/kg) along the reclamation chronosequence over different land uses. Figure S1: The optically stimulated luminescence (OSL) dating procedure. Figure S2: The linear correlation between soil CaCO3 and pH.
Author Contributions
Conceptualization, B.S. and C.G.; methodology, B.S.; software, B.S. and H.Z.; validation, C.G. and A.M.M.; formal analysis, B.S. and S.Y.; investigation, B.S., Y.Z., S.S., and H.J.; resources, C.G.; data curation, C.G.; writing—original draft preparation, B.S.; writing—review and editing, C.G.; visualization, B.S.; supervision, C.G.; project administration, C.G.; funding acquisition, C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Project No.41877002).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zheng, S.M.; Xia, Y.H.; Hu, Y.J.; Chen, X.B.; Rui, Y.C.; Gunina, A.; He, X.Y.; Ge, T.D.; Wu, J.S.; Su, Y.R.; et al. Stoichiometry of carbon, nitrogen, and phosphorus in soil: Effects of agricultural land use and climate at a continental scale. Soil Till. Res. 2021, 209, 104903. [Google Scholar] [CrossRef]
- Tian, H.Q.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Finzi, A.C.; Austin, A.T.; Cleland, E.E.; Frey, S.D.; Houlton, B.Z.; Wallenstein, M. Responses and feedbacks of coupled biogeochemical cycles to climate change: Examples from terrestrial ecosystems. Front. Ecol. Environ. 2011, 9, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.F.; Yang, X.T.; Deng, Q.; Liu, Q.; Wang, X.; Yang, H.; Ren, H. Soil C:N:P stoichiometry in tropical forests on Hainan Island of China: Spatial and vertical variations. Catena 2021, 201, 105228. [Google Scholar] [CrossRef]
- Ostrowska, A.; Porębska, G. Assessment of the C/N ratio as an indicator of the decomposability of organic matter in forest soils. Ecol. Indic. 2015, 49, 104–109. [Google Scholar] [CrossRef]
- Zhang, J.P.; Shen, C.D.; Ren, H.; Wang, J.; Han, W.D. Estimating change in sedimentary organic carbon content during mangrove restoration in southern China using carbon isotopic measurements. Pedosphere 2012, 22, 58–66. [Google Scholar] [CrossRef]
- Du, E.Z.; Doorn, M.; Vries, W. Spatially divergent trends of nitrogen versus phosphorus limitation across European forests. Sci. Total Environ. 2021, 771, 145391. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Rivas-ubach, A.; Janssens, I.A. The human-induced imbalance between C, N and P in Earth’s life system. Glob. Chang. Biol. 2012, 18, 3–6. [Google Scholar] [CrossRef]
- Wang, X.G.; Lü, X.T.; Zhang, H.Y.; Dijkstra, F.A.; Jiang, Y.G.; Wang, X.B.; Lu, J.Y.; Wuyunna; Wang, Z.W.; Han, X.G. Changes in soil C:N:P stoichiometry along an aridity gradient in drylands of northern China. Geoderma 2020, 361, 114087. [Google Scholar] [CrossRef]
- Guan, Y.J.; Zhou, W.; Bai, Z.K.; Cao, Y.G.; Huang, Y.H.; Huang, H.Y. Soil nutrient variations among different land use types after reclamation in the Pingshuo opencast coal mine on the Loess Plateau, China. Catena 2020, 188, 104427. [Google Scholar] [CrossRef]
- Huang, L.M.; Thompson, A.; Zhang, G.L. Long-term paddy cultivation significantly alters topsoil phosphorus transformation and degrades phosphorus sorption capacity. Soil Till. Res. 2014, 142, 32–41. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Gómez, M.; Soliveres, S.; García-Palacios, P.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, P.B.; Yin, A.J.; Yang, X.H.; Zhang, X.; Zhang, M.; Gao, C. Organic carbon and total nitrogen dynamics of reclaimed soils following intensive agricultural use in eastern China. Agric. Ecosyst. Environ. 2016, 235, 193–203. [Google Scholar] [CrossRef]
- Chen, L.M.; Zhang, G.L. Parent material uniformity and evolution of soil characteristics of a paddy soil chronosequence derived from marine sediments. Acta Pedol. Sin. 2009, 46, 753–763. (In Chinese) [Google Scholar]
- Chen, L.M.; Zhang, G.L.; Effland, W.R. Soil characteristic response times and pedogenic thresholds during the 1000-Year evolution of a paddy soil chronosequence. Soil Sci. Soc. Am. J. 2011, 75, 1807–1820. [Google Scholar] [CrossRef]
- Yin, A.J.; Gao, C.; Zhang, M.; Wu, P.B.; Yang, X.H. Rapid changes in phosphorus species in soils developed on reclaimed tidal flat sediments. Geoderma 2017, 307, 46–53. [Google Scholar] [CrossRef]
- Edwards, P.J.; Jordan, C.F. Nutrient Cycling in Tropical Forest Ecosystems. J. Ecol. 1987, 75, 280. [Google Scholar] [CrossRef]
- Du, L.; Zhang, X.Z.; Zheng, Z.C.; Li, T.X.; Wang, Y.D.; Huang, H.G.; Yu, H.Y.; Ye, D.H.; Liu, T. Paddy soil nutrients and stoichiometric ratios as affected by anthropogenic activities during long-term tillage process in Chengdu Plain. J. Soil Sediments 2020, 20, 3835–3845. [Google Scholar] [CrossRef]
- Zhou, Y.; Boutton, T.W.; Wu, X.B. Soil C:N:P stoichiometry responds to vegetation change from grassland to woodland. Biogeochemistry 2018, 140, 341–357. [Google Scholar] [CrossRef]
- Liu, X.; Ma, J.; Ma, Z.W.; Li, L.H. Soil nutrient contents and stoichiometry as affected by land-use in an agro-pastoral region of northwest China. Catena 2016, 150, 146–153. [Google Scholar] [CrossRef]
- Kong, X.B.; Zhang, F.R.; Wei, Q.; Xu, Y.; Hui, J.G. Influence of land use change on soil nutrients in an intensive agricultural region of North China. Soil Till. Res. 2006, 88, 85–94. [Google Scholar] [CrossRef]
- Roth, P.J.; Lehndorff, E.; Cao, Z.H.; Zhuang, S.Y.; Bannert, A.; Wissing, L.; Schloter, M.; Kögel-Knabner, I.; Amelung, W. Accumulation of nitrogen and microbial residues during 2000 years of rice paddy and non-paddy soil development in the Yangtze River Delta, China. Glob. Chang. Biol. 2011, 17, 3405–3417. [Google Scholar] [CrossRef]
- Li, J.G.; Wan, X.; Liu, X.X.; Chen, Y.; Slaughter, L.C.; Weindorf, D.C.; Dong, Y.H. Changes in soil physical and chemical characteristics in intensively cultivated greenhouse vegetable fields in North China. Soil Till. Res. 2019, 195, 104366. [Google Scholar] [CrossRef]
- Cui, J.; Liu, C.; Li, Z.L.; Wang, L.; Chen, X.F.; Ye, Z.Z.; Fang, C.M. Long-term changes in topsoil chemical properties under centuries of cultivation after reclamation of coastal wetlands in the Yangtze Estuary, China. Soil Till. Res. 2012, 123, 50–60. [Google Scholar] [CrossRef]
- Tian, L.M.; Zhao, L.; Wu, X.D.; Fang, H.B.; Zhao, Y.H.; Hu, G.J.; Yue, G.Y.; Sheng, Y.; Wu, J.C.; Chen, J.; et al. Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grassland. Sci. Total Environ. 2018, 622–623, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Tie, L.H.; Zhang, S.B.; Peñuelas, J.; Sardans, J.; Zhou, S.X.; Hu, J.X.; Huang, C.D. Responses of soil C, N, and P stoichiometric ratios to N and S additions in a subtropical evergreen broad-leaved forest. Geoderma 2020, 379, 114633. [Google Scholar] [CrossRef]
- Wang, X.H.; Yang, J.H.; Xie, X.F.; Chen, X.J.; Pu, L.J.; Zhang, X.W. Soil microbial succession with soil development since costal reclamation. Catena 2020, 187, 104393. [Google Scholar] [CrossRef]
- Cheng, Y.Q.; Yang, L.Z.; Cao, Z.H.; Ci, E.; Yin, S.X. Chronosequential changes of selected pedogenic properties in paddy soils as compared with non-paddy soils. Geoderma 2009, 151, 31–41. [Google Scholar] [CrossRef]
- Sun, L.; Xun, W.B.; Huang, T.; Zhang, G.S.; Gao, J.S.; Ran, W.; Li, D.C.; Shen, Q.R.; Zhang, R.F. Alteration of the soil bacterial community during parent material maturation driven by different fertilization treatments. Soil Biol. Biochem. 2016, 96, 207–215. [Google Scholar] [CrossRef]
- Sun, R.B.; Zhang, X.X.; Guo, X.S.; Wang, D.Z.; Chu, H.Y. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Dai, S.B.; Lu, X.X. Sediment load change in the Yangtze River (Changjiang): A review. Geomorphology 2014, 215, 60–73. [Google Scholar] [CrossRef]
- Zhang, X.; Liao, X.L.; Huang, L.B.; Shan, Q.Y.; Hu, A.Y.; Yan, D.Z.; Zhang, J.; Long, X.E. Soil profile rather than reclamation time drives the mudflat soil microbial community in the wheat-maize rotation system of Nantong, China. J. Soil Sediments 2021, 21, 1672–1687. [Google Scholar] [CrossRef]
- Guo, L.Y.; Wang, D.L.; Wang, J.Y. Analysis of the arable land change and its causes in the eastern coastal China over the last decades. Chin. J. Agric. Resour. Reg. Plan. 2012, 33, 6–10. (In Chinese) [Google Scholar]
- Local Chronicles Compilation Committee of Wuwei County (LCCCW). Wuwei Prefectural Annals; Social Sciences Academic Press (China), SSAP: Beijing, China, 1993. (In Chinese) [Google Scholar]
- Records of Wuwei levee (RWL); Beijing Jiuzhou Publishing House: Beijing, China, 2005. (In Chinese)
- Madsen, A.T.; Murray, A.S. Optically stimulated luminescence dating of young sediments: A review. Geomorphology 2009, 109, 3–16. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; American Association of Agronomy: Madison, Wisconsin, 1982. [Google Scholar]
- Page, A.L. Methods of Soil Analysis, Part 2: Chemical and Microbial Properties, 2nd ed.; American Association of Agronomy: Madison, Wisconsin, 1982. [Google Scholar]
- Lu, R. Methods of Soil Agrochemistry Analysis; Agricultural Science and Technology Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Zhong, Z.K.; Wang, X.; Zhang, X.Y.; Zhang, W.; Xu, Y.D.; Ren, C.J.; Han, X.H.; Yang, G.H. Edaphic factors but not plant characteristics mainly alter soil microbial properties along a restoration chronosequence of Pinus tabulaeformis stands on Mt. Ziwuling, China. For. Ecol Manag. 2019, 453, 117625. [Google Scholar] [CrossRef]
- Zhang, H.; Yin, A.J.; Chen, Y.H.; Shao, S.S.; Wu, J.T.; Fan, M.M.; Chen, F.R.; Gao, C. Machine learning-based source identification and spatial prediction of heavy metals in soil in a rapid urbanization area, eastern China. J. Clean Prod. 2020, 273, 122858. [Google Scholar] [CrossRef]
- Tian, J.; Bu, L.Y.; Zhang, M.X.; Yuan, J.W.; Zhang, Y.L.; Wei, G.H.; Wang, H.L. Soil bacteria with distinct diversity and functions mediates the soil nutrients after introducing leguminous shrub in desert ecosystems. Glob. Ecol. Conserv. 2021, 31, e01841. [Google Scholar] [CrossRef]
- Jiang, S.; Xing, Y.J.; Liu, G.C.; Hu, C.Y.; Wang, X.C.; Yan, G.Y.; Wang, Q.G. Changes in soil bacterial and fungal community composition and functional groups during the succession of boreal forests. Soil Biol. Biochem. 2021, 161, 108393. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and User’s Guide: Software for Canonical Community Ordination (Version 5.0); Biometris: Wageningen, The Netherlands; České Budĕjovice, Czech Republic, 2002. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Breiman, L. Manual on Setting Up, Using, and Understanding Random Forests V3.1. 2002. Available online: https://www.stat.berkeley.edu/~breiman/Using_random_forests_V3.1.pdf (accessed on 10 October 2022).
- Chen, J.; Zhang, H.; Fan, M.M.; Chen, F.R.; Gao, C. Machine-learning-based prediction and key factor identification of the organic carbon in riverine floodplain soils with intensive agricultural practices. J. Soil Sediments 2021, 21, 2896–2907. [Google Scholar] [CrossRef]
- Tan, J.L.; Kang, Y.H. Changes in soil properties under the influences of cropping and drip irrigation during the reclamation of severe salt-affected soils. Agric. Sci. China 2009, 8, 1228–1237. [Google Scholar] [CrossRef]
- Wissing, L.; Kölbl, A.; Schad, P.; Bräuer, T.; Cao, Z.-H.; Kögel-Knabner, I. Organic carbon accumulation on soil mineral surfaces in paddy soils derived from tidal wetlands. Geoderma 2014, 228-229, 90–103. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Adatte, T.; Verrecchia, E.P. A cascading influence of calcium carbonate on the biogeochemistry and pedogenic trajectories of subalpine soils, Switzerland. Geoderma 2019, 361, 114065. [Google Scholar] [CrossRef]
- Adams, M.L.; Hawke, D.J.; Nilsson, N.; Powell, K.J. The relationship between soil solution pH and Al3+concentrations in a range of South Island (New Zealand) soils. Soil Res. 2000, 38, 141–154. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Verrecchia, É.P. Calcium-mediated stabilisation of soil organic carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef]
- Huang, L.-M.; Thompson, A.; Zhang, G.-L.; Chen, L.-M.; Han, G.-Z.; Gong, Z.-T. The use of chronosequences in studies of paddy soil evolution: A review. Geoderma 2015, 237-238, 199–210. [Google Scholar] [CrossRef]
- Johnson, D.L.; Watson-Stegner, D. Evolution model of pedogenesis. Soil Sci. 1986, 143, 349–366. [Google Scholar] [CrossRef]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Yavitt, J.B.; Harms, K.E.; Garcia, M.N.; Mirabello, M.J.; Wright, S.J. Soil fertility and fine root dynamics in response to 4 years of nutrient (N, P, K) fertilization in a lowland tropical moist forest, Panama. Austral Ecology 2011, 36, 433–445. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, M.; Huang, Z.; Lin, T.C.; Vadeboncoeur, M.A.; Searle, E.B.; Chen, H.Y. Temporal changes in soil C-N-P stoichiometry over the past 60 years across subtropical China. Glob. Chang. Biol. 2018, 24, 1308–1320. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.H.; Huang, Z.Q.; He, Z.M.; Yu, Z.P.; Wang, M.H.; Murry, R.D.; Yang, Y.S. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Cong, R.H.; Wang, X.J.; Xu, M.G.; Zhang, W.J.; Xie, L.J.; Yang, X.Y.; Huang, S.M.; Wang, B.R. Dynamics of soil carbon to nitrogen ratio changes under long-term fertilizer addition in wheat-corn double cropping systems of China. Eur. J. Soil Sci. 2012, 63, 341–350. [Google Scholar] [CrossRef]
- Zhu, Z.K.; Zhou, J.; Shahbaz, M.; Tang, H.M.; Liu, S.L.; Zhang, W.J.; Yuan, H.Z.; Zhou, P.; Alharbi, H.; Wu, J.S.; et al. Microorganisms maintain C:N stoichiometric balance by regulating the priming effect in long-term fertilized soils. Appl. Soil Ecol. 2021, 167, 104033. [Google Scholar] [CrossRef]
- Deng, M.F.; Liu, L.L.; Sun, Z.Z.; Piao, S.L.; Ma, Y.C.; Chen, Y.W.; Wang, J.; Qiao, C.L.; Wang, X.; Li, P. Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-rupprechtii plantations. New Phytol. 2016, 212, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, S.L.; Yu, G.R. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Glob. Chang. Biol. 2016, 22, 934–943. [Google Scholar] [CrossRef]
- Seidel, F.; Lopez C., M.L.; Bonifacio, E.; Kurokawa, H.; Yamanaka, T.; Celi, L. Seasonal phosphorus and nitrogen cycling in four Japanese cool-temperate forest species. Plant Soil 2022, 472, 391–406. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef]
- Salem, M.A.; Al-Ethawi, L.A.; Eldrazi, Z.; Nouraldien, A.I. A case study of the total and available phosphorus concentration in libyan agricultural soils in different depths and seasons in long-term chemical and animal manure fertilization. Int. J. Res. Stud. Biosci. 2014, 2, 1–9. [Google Scholar]
- Gamboa, A.M.; Galicia, L. Differential influence of land use/cover change on topsoil carbon and microbial activity in low-latitude temperate forests. Agric. Ecosyst. Environ. 2011, 142, 280–290. [Google Scholar] [CrossRef]
- Liu, Y.L.; Wang, P.; Pan, G.X.; Crowley, D.; Li, L.Q.; Zheng, J.W.; Zhang, X.H.; Zheng, J.F. Functional and structural responses of bacterial and fungal communities from paddy fields following long-term rice cultivation. J. Soils Sediments 2015, 16, 1460–1471. [Google Scholar] [CrossRef]
- Lin, S.Y.; Wang, W.Q.; Vancov, Y.; Lai, D.Y.F.; Wang, C.; Wiesmeier, M.; Jin, Q.; Liu, X.Y.; Fang, Y.Y. Soil carbon, nutrients and their stoichiometry decrement in relation to paddy field degradation: Investigation in a subtropical region. Catena 2022, 217, 106484. [Google Scholar] [CrossRef]
- Wang, X.Y.; Li, Y.Q.; Gong, X.W.; Niu, Y.Y.; Chen, Y.P.; Shi, X.P.; Li, W. Storage, pattern and driving factors of soil organic carbon in an ecologically fragile zone of northern China. Geoderma 2019, 343, 155–165. [Google Scholar] [CrossRef]
- Wei, L.; Ge, T.D.; Zhu, Z.K.; Luo, Y.; Yang, Y.H.; Xiao, M.L.; Yan, Z.F.; Li, Y.H.; Wu, J.S.; Kuzyakov, Y. Comparing carbon and nitrogen stocks in paddy and upland soils: Accumulation, stabilization mechanisms, and environmental drivers. Geoderma 2021, 398, 115121. [Google Scholar]
- Zhao, Z.J.; Jin, R.; Fang, D.; Wang, H.; Dong, Y.; Xu, R.K.; Jiang, J. Paddy cultivation significantly alters the forms and contents of Fe oxides in an Oxisol and increases phosphate mobility. Soil Till. Res. 2018, 184, 176–180. [Google Scholar] [CrossRef]
- Zou, P.; Fu, J.R.; Cao, Z.H. Chronosequence of paddy soils and phosphorus sorption–desorption properties. J. Soil Sediments 2011, 11, 249–259. [Google Scholar] [CrossRef]
- Wang, J.; Zhuang, S.Y.; Zhu, Z.L. Soil organic nitrogen composition and mineralization of paddy soils in a cultivation chronosequence in China. J. Soil Sediments 2017, 17, 1588–1598. [Google Scholar] [CrossRef]
- Ezrin, M.H.; Amin, M.; Anuar, A.R.; Aimrun, W. Rice yield prediction using apparent electrical conductivity of paddy soils. Eur. J. Sci. Res. 2009, 37, 575–590. [Google Scholar]
- Li, H.; Zhang, Y.Y.; Yang, S.; Wang, Z.R.; Feng, X.; Liu, H.Y.; Jiang, Y. Variations in soil bacterial taxonomic profiles and putative functions in response to straw incorporation combined with N fertilization during the maize growing season. Agric. Ecosyst. Environ. 2019, 283, 106578. [Google Scholar] [CrossRef]
- Statistical Bereau of Anhui Province (SBAP). Statistical Yearbook of Anhui Province. 2019. Available online: http://tjj.ah.gov.cn/oldfiles/tjj/tjjweb/tjnj/2019/cn.html (accessed on 22 November 2022).
- Lei, Y.; Song, B.; Saakes, M.; van der Weijden, R.D.; Buisman, C.J. Interaction of calcium, phosphorus and natural organic matter in electrochemical recovery of phosphate. Water Res. 2018, 142, 10–17. [Google Scholar] [CrossRef]
- Peng, Y.; Sun, Y.; Fan, B.; Zhang, S.; Bolan, N.S.; Chen, Q.; Tsang, D.C. Fe/Al (hydr)oxides engineered biochar for reducing phosphorus leaching from a fertile calcareous soil. J. Clean Prod. 2021, 279, 123877. [Google Scholar] [CrossRef]
- Silver, W.L.; Neff, J.; Mcgroddy, M.; Veldkamp, E.; Cosme, K.R. Effects of soil texture on belowground carbon and nutrient storage in a lowland amazonian forest ecosystem. Ecosystems 2000, 3, 193–209. [Google Scholar] [CrossRef]
- Chen, J.S.; Chiu, C.Y. Characterization of soil organic matter in different particle-size fractions in humid subalpine soils by CP/MAS 13C NMR. Geoderma 2003, 117, 129–141. [Google Scholar] [CrossRef]
- Hou, E.Q.; Chen, C.R.; Luo, Y.Q.; Zhou, G.Y.; Kuang, Y.W.; Zhang, Y.G.; Heenan, M.; Lu, X.K.; Wen, D.Z. Effects of climate on soil phosphorus cycle and availability in natural terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 3344–3356. [Google Scholar] [CrossRef]
- Pan, W.L.; Kidwell, K.K.; McCracken, V.A.; Bolton, R.P.; Allen, M. Economically optimal wheat yield, protein and nitrogen use component responses to varying N supply and genotype. Front. Plant Sci. 2020, 10, 1790. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.L.; Wu, Q.X.; Zhu, J.Q. Nitrogen and phosphorus losses from paddy fields and the yield of rice with different water and nitrogen management practices. Sci. Rep. 2020, 10, 9734. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ye, Q.; Li, J.W.; Wang, X.D.; Li, X.; Ruan, L.; Tao, A. Determination for the uniformity of parent material of basalt-developed soil in the xinsheng basin. Chin. J. Soil Sci. 2021, 52, 253–260. (In Chinese) [Google Scholar]
- Huang, L.M.; Jia, X.X.; Shao, M.A.; Chen, L.M.; Han, G.Z.; Zhang, G.L. Phases and rates of iron and magnetism changes during paddy soil development on calcareous marine sediment and acid Quaternary red-clay. Sci. Rep. 2018, 8, 444. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).