


Hemp Agronomy: Current Advances, Questions, Challenges, and Opportunities
 ,
, 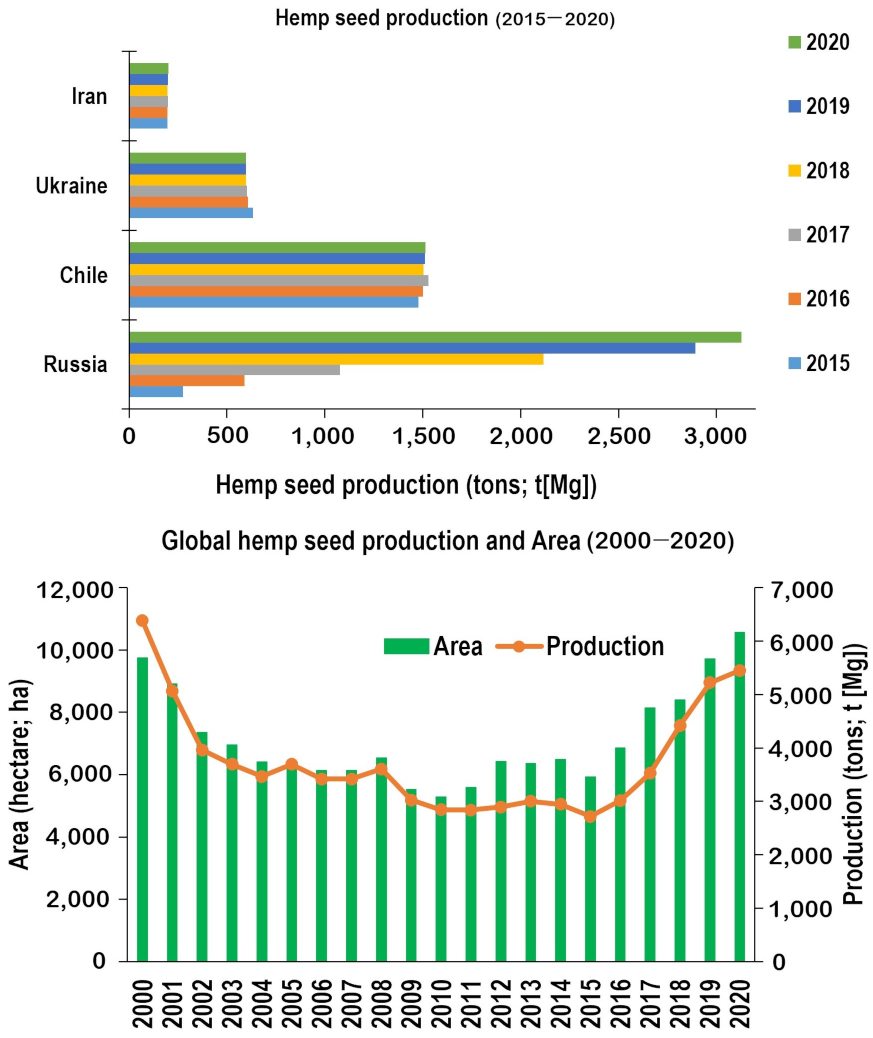{kind=link}
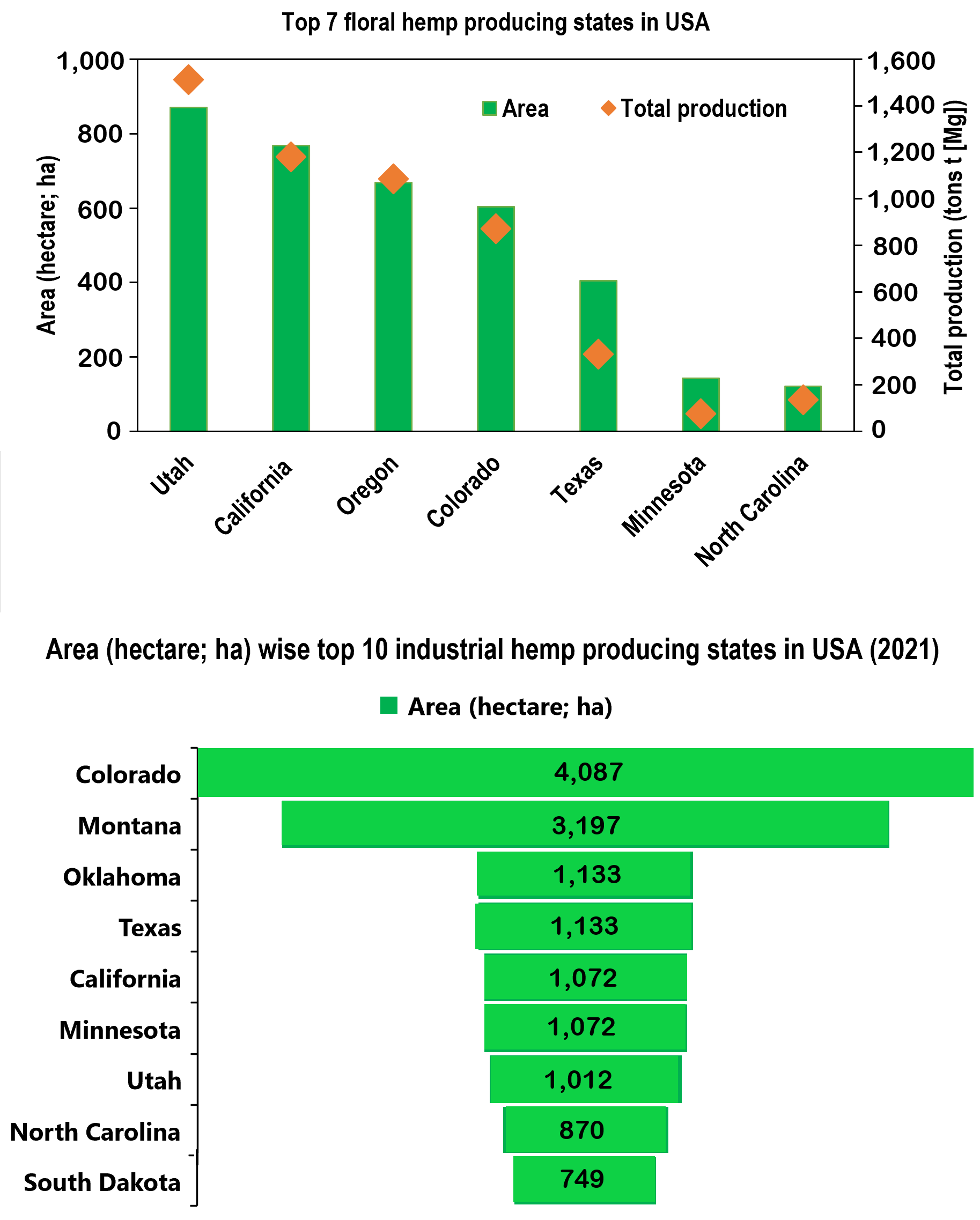{kind=link}
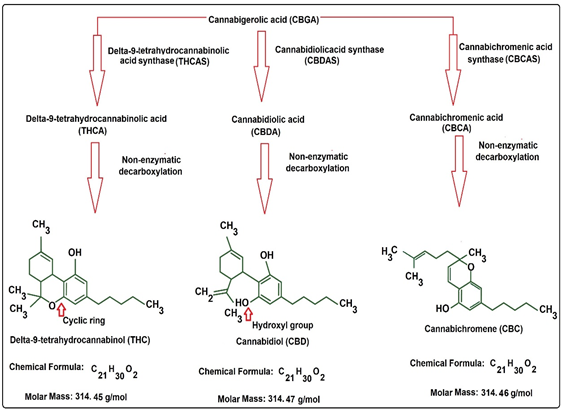{kind=link}
Abstract
1. Introduction
2. Factors Influencing Growth, Development, and Yield of Hemp
2.1. Water Deficit Stress
2.2. Heat Stress
2.3. Salinity Stress
- How feasible is it to manipulate and standardize the light spectrum for the cultivation of region-specific diverse hemp under a controlled environment with an emphasis on precise production of cannabinoids for medicinal purposes?
- Are there any possibilities to develop a photoperiod-insensitive hybrid for the production of cannabinoids, seed and seed oil, and fiber?
- Is it feasible to cultivate hemp chemotype III (CBD dominant with less than 0.3% THC; genotypes for medicinal cannabinoid production) in tropical and subtropical regions with weather conditions not well suited for genotypes grown for seed and fiber yield?
- What is the role of hemp roots in confronting diverse abiotic stress factors, particularly, water deficit and high day and night temperatures?
- Irrespective of the types of hemp, what is the magnitude of tolerance of hemp genotypes to agroclimatic regions with high humidity such as the tropics?
3. Improving Hemp Production by Modifying Agronomic Practices
3.1. Nitrogen Amount and Timing
3.2. P and K Amount and Timing
3.3. Seeding Density and Plant Density
3.4. Photoperiod Effects on Hemp Growth
3.5. Harvest Timing
3.6. Multiple Factors and Hemp Growth
- What is the feasibility of intercropping hemp with other crops such as cereal crops or legumes to increase the economic benefit of farmers and reduce nitrogen input?
- What are the relationships between plant density, harvesting time, soil nutrient and water availability, and cannabinoid and fatty acid profiles in low latitude regions?
4. Advances in Cannabinoid Detection and Quantification
- How can a universal standard cannabis extraction and assay protocol be developed based on different cannabis assay techniques?
5. Environmental Impact of Hemp
Phytoremediation, Carbon Sequestration and Bioenergy
- Is it feasible to utilize hemp for the phytoremediation of petroleum-contaminated sites (petroleum hydrocarbons; PAHs) and utilize biomass for the production of bioenergy?
- What is the feasibility to engage hemp genotypes with higher root growth and biomass to remediate heavy metal polluted locations?
- Several studies report uptake of heavy metals by hemp roots. Can roots be harvested economically or is higher root uptake also matched by higher concentration in leaves?
- How do hemp roots take part in phytoremediation, and can hemp genotypes with efficient hydraulic control increase foliar transpiration for the faster removal of contaminants?
- What is the precise carbon sequestration capacity of hemp in open field conditions?
- What are the possible ways to increase the carbon sequestration capacity of hemp by developing value-added products from hemp?
- How to increase bioenergy production from hemp with low input and processing costs?
6. Hemp Breeding
6.1. Public Hemp Breeding in the United States
6.2. Hemp Breeding in Other Countries
6.3. Breeding Methods and Breeding Goals
- Is it possible to develop hemp cultivars with THC levels low enough to allow them to be grown without exceeding legal THC limits under a wide array of agronomic and environmental conditions?
- Can gene editing efforts be useful in this regard?
- Although it has been reported that MAS (marker-assisted selection) was of low utility previously, with more advanced genomic tools, can this be changed?
- How many regions in hemp genomes can be possibly identified by genomic tools to elucidate the separation between drug and fiber types?
- Is it possible to use breeding to produce hemp cultivars with high levels of specific CBDs?
- Aside from CBD and THC contents, can new cultivars be developed that have suitable combinations of fiber content, disease resistance, and drought tolerance to make large-scale production for the fiber market feasible on land that would be otherwise unsuitable for agricultural production (for example, no irrigation in semi-arid climates)?
- Can alternative uses for hemp (e.g., hempcrete) be further developed, and cultivars with suitable agronomic traits be developed, that would allow for expanded markets for the crop?
- As the number of breeding programs in the U.S. is limited and the programs are fairly new, how well adapted are hemp cultivars across regions?
- There are numerous marijuana varieties available. Have these been developed by crossing or simply selection from existing materials? Has a screen of existing germplasm for useful traits been conducted in a systematic manner for industrial hemp?
7. Discussion and Future Perspectives
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fike, J. Industrial Hemp: Renewed Opportunities for an Ancient Crop. Crit. Rev. Plant Sci. 2016, 35, 406–424. [Google Scholar] [CrossRef]
- Williams, D.W. Industrial Hemp as a Modern Commodity Crop, 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 2020; Volume 182. [Google Scholar]
- Clarke, R.; Merlin, M. Cannabis Domestication, Breeding History, Present-day Genetic Diversity, and Future Prospects. Crit. Rev. Plant Sci. 2016, 35, 293–327. [Google Scholar] [CrossRef]
- Liu, F.H.; Hu, H.R.; Du, G.H.; Deng, G.; Yang, Y. Ethnobotanical Research on Origin, Cultivation, Distribution and Utilization of Hemp (Cannabis sativa L.) in China. Indian J. Tradit. Knowl. 2017, 16, 235–242. [Google Scholar]
- Rahn, B.; Pearson, B.J.; Trigiano, R.N.; Gray, D.J. The Derivation of Modern Cannabis Varieties. Crit. Rev. Plant Sci. 2017, 35, 328–348. [Google Scholar] [CrossRef]
- Ehrensing, D. Feasibility of Industrial Hemp Production in the United States Pacific Northwest; Oregon State University Extension Service: Corvallis, OR, USA, 1998; Report No. 681. [Google Scholar]
- USDA. Industrial Hemp in the United States: Status and Market Potential.; Economic Research Service. Report No. AGES-001E; USDA: Washington, DC, USA, 2000; p. 46.
- Zhang, M.; Anderson, S.L.; Brym, Z.T.; Pearson, B.J. Photoperiodic Flowering Response of Essential Oil, Grain, and Fiber Hemp (Cannabis sativa L.) Cultivars. Front. Plant Sci. 2021, 12, 1498. [Google Scholar] [CrossRef]
- Cherney, J.H.; Small, E. Industrial Hemp in North America: Production, Politics and Potential. Agronomy 2016, 6, 58. [Google Scholar] [CrossRef]
- Jiang, Y.; Sun, Y.; Zheng, D.; Han, C.; Cao, K.; Xu, L.; Liu, S.; Cao, Y.; Feng, N. Physiological and Transcriptome Analyses for Assessing the Effects of Exogenous Uniconazole on Drought Tolerance in Hemp (Cannabis sativa L.). Sci. Rep. 2021, 11, 14476. [Google Scholar] [CrossRef]
- Toth, J.A.; Smart, L.B.; Smart, C.D.; Stack, G.M.; Carlson, C.H.; Philippe, G.; Rose, J.K. Limited Effect of Environmental Stress on Cannabinoid Profiles in High-Cannabidiol Hemp (Cannabis sativa L.). GCB Bioenergy 2021, 13, 1666–1674. [Google Scholar] [CrossRef]
- Lazarjani, M.P.; Torres, S.; Hooker, T.; Fowlie, C.; Young, O.; Seyfoddin, A. Methods for Quantification of Cannabinoids: A Narrative Review. J. Cannabis Res. 2020, 2, 35. [Google Scholar] [CrossRef]
- Johnson, R. Hemp as an Agricultural Commodity; Congressional Research Service: Washington, DC, USA, 2018.
- Wimalasiri, E.M.; Jahanshiri, E.; Chimonyo, V.G.; Kuruppuarachchi, N.; Suhairi, T.; Azam-Ali, S.N.; Gregory, P.J. A Framework for the Development of Hemp (Cannabis sativa L.) as a Crop for the Future in Tropical Environments. Ind. Crops Prod. 2021, 172, 113999. [Google Scholar] [CrossRef]
- Malone, T.; Gomez, K. Hemp in the United States: A Case Study of Regulatory Path Dependence. Appl. Econ. Perspect. Policy 2019, 41, 199–214. [Google Scholar] [CrossRef]
- NCSL (National Conference of State Legislatures). Available online: https://www.ncsl.org/research/agriculture-and-rural-development/state-industrial-hemp-statutes (accessed on 22 December 2022).
- Hall, J.; Bhattarai, S.P.; Midmore, D.J. Effect of Industrial Hemp (Cannabis sativa L.) Planting Density on Weed Suppression, Crop Growth, Physiological Responses, and Fibre Yield in the Subtropics. Renew. Bioresour. 2014, 2, 1. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Santini, G.; Fiorini, D.; Sut, S.; Dall’Acqua, S.; Canale, A.; Maggi, F. The Essential Oil from Industrial Hemp (Cannabis sativa L.) By-Products as an Effective Tool for Insect Pest Management in Organic Crops. Ind. Crops Prod. 2018, 122, 308–315. [Google Scholar] [CrossRef]
- Amaducci, S.; Zatta, A.; Raffanini, M.; Venturi, G. Characterisation of Hemp (Cannabis sativa L.) Roots under Different Growing Conditions. Plant Soil 2008, 313, 227–235. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#rankings/countries_by_commodity (accessed on 22 December 2022).
- Globalnewswire. Global Cannabidiol (CBD) Market Report 2022. Available online: https://www.globenewswire.com/news-release/2022/08/03/2491082/28124/en/Global-Cannabidiol-CBD-Market-Report-2022-Featuring-Key-Players-Canopy-Growth-Corporation-High-Tide-Aurora-Pharmaceutical-others.html (accessed on 22 December 2022).
- NASS (National Agriculture Statistics Service). National Hemp Report. Agriculture Statistical Board; United States Department of Agriculture (USDA): Washington, DC, USA, 2022; pp. 1–24.
- Brand, E.J.; Zhao, Z. Cannabis in Chinese Medicine: Are Some Traditional Indications Referenced in Ancient Literature Related to Cannabinoids? Front. Pharmacol. 2017, 8, 108. [Google Scholar] [CrossRef]
- Touw, M. The Religious and Medicinal Uses of Cannabis in China, India and Tibet. J. Psychoact. Drugs 1981, 13, 23–34. [Google Scholar] [CrossRef]
- Butrica, J.L. The Medical Use of Cannabis Among the Greeks and Romans. J. Cannabis Ther. 2002, 2, 51–70. [Google Scholar] [CrossRef]
- Rheay, H.T.; Omondi, E.C.; Brewer, C.E. Potential of Hemp (Cannabis sativa L.) for Paired Phytoremediation and Bioenergy Production. GCB Bioenergy 2020, 13, 525–536. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Durán Zuazo, V.H.; Sánchez-Carnenero, C.; Hernández, A.; Ferreiro-Vera, C.; Casano, S. Seeking Suitable Agronomical Practices for Industrial Hemp (Cannabis sativa L.) Cultivation for Biomedical Applications. Ind. Crops Prod. 2019, 139, 111524. [Google Scholar] [CrossRef]
- Wang, H.; Xu, Z.; Kohandehghan, A.; Li, Z.; Cui, K.; Tan, X.; Stephenson, T.J.; King’Ondu, C.K.; Holt, C.M.; Olsen, B.C. Interconnected Carbon Nanosheets Derived from Hemp for Ultrafast Supercapacitors with High Energy. ACS Nano 2013, 7, 5131–5141. [Google Scholar] [CrossRef]
- Khattab, M.M.; Dahman, Y. Production and Recovery of Poly-3-Hydroxybutyrate Bioplastics using Agro-Industrial Residues of Hemp Hurd Biomass. Bioprocess Biosyst. Eng. 2019, 42, 1115–1127. [Google Scholar] [CrossRef]
- Asli, M.; Brachelet, F.; Sassine, E.; Antczak, E. Thermal and Hygroscopic Study of Hemp Concrete in Real Ambient Conditions. J. Build. Eng. 2021, 44, 102612. [Google Scholar] [CrossRef]
- Schluttenhofer, C.; Yuan, L. Challenges Towards Revitalizing Hemp: A Multifaceted Crop. Trends Plant Sci. 2017, 22, 917–929. [Google Scholar] [CrossRef]
- Salentijn, E.M.; Petit, J.; Trindade, L.M. The Complex Interactions Between Flowering Behavior and Fiber Quality in Hemp. Front. Plant Sci. 2019, 10, 614. [Google Scholar] [CrossRef]
- Fudge, D. Fifty years of J. R. Platt’s strong inference. J. Exp. Biol. 2014, 217, 1202–1204. [Google Scholar] [CrossRef]
- Farrell, M.J. The Measurement of Productive Efficiency. J. R. Stat. Society. Ser. A 1957, 120, 253–290. [Google Scholar] [CrossRef]
- Petit, J.; Salentijn, E.M.; Paulo, M.-J.; Thouminot, C.; van Dinter, B.J.; Magagnini, G.; Gusovius, H.-J.; Tang, K.; Amaducci, S.; Wang, S. Genetic Variability of Morphological, Flowering, and Biomass Quality Traits in Hemp (Cannabis sativa L.). Front. Plant Sci. 2020, 11, 102. [Google Scholar] [CrossRef]
- Schäfer, T.; Honermeier, B. Effect of sowing date and plant density on the cell morphology of hemp (Cannabis sativa L.). Ind. Crops Prod. 2006, 23, 88–98. [Google Scholar] [CrossRef]
- Amaducci, S.; Colauzzi, M.; Zatta, A.; Venturi, G. Flowering Dynamics in Monoecious and Dioecious Hemp Genotypes. J. Ind. Hemp 2008, 13, 5–19. [Google Scholar] [CrossRef]
- Hall, J.; Bhattarai, S.P.; Midmore, D.J. Review of Flowering Control in Industrial Hemp. J. Nat. Fibers 2012, 9, 23–36. [Google Scholar] [CrossRef]
- Hall, J.; Bhattarai, S.P.; Midmore, D.J. The Effects of Different Sowing Times on Maturity Rates, Biomass, and Plant Growth of Industrial Fiber Hemp. J. Nat. Fibers 2013, 10, 40–50. [Google Scholar] [CrossRef]
- Hall, J.; Bhattarai, S.P.; Midmore, D.J. The Effects of Photoperiod on Phenological Development and Yields of Industrial Hemp. J. Nat. Fibers 2014, 11, 87–106. [Google Scholar] [CrossRef]
- Sengloung, T.; Kaveeta, L.; Nanakorn, W. Effect of Sowing Date on Growth and Development of Thai Hemp (Cannabis sativa L.). Agric. Nat. Resour. 2009, 43, 423–431. [Google Scholar]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.; Rahman, M.H.; Cheong, E.J.; Lim, J.-D.; Lim, Y.-S. Cannabinoids Accumulation in Hemp (Cannabis sativa L.) Plants under LED Light Spectra and Their Discrete Role as a Stress Marker. Biology 2021, 10, 710. [Google Scholar] [CrossRef]
- Baldini, M.; Ferfuia, C.; Zuliani, F.; Danuso, F. Suitability Assessment of Different Hemp (Cannabis Sativa L.) Varieties to the Cultivation Environment. Ind. Crops Prod. 2020, 143, 111860. [Google Scholar] [CrossRef]
- Anwar, F.; Latif, S.; Ashraf, M. Analytical Characterization of Hemp (Cannabis sativa) Seed Oil from Different Agro-Ecological Zones of Pakistan. J. Am. Oil Chem. Soc. 2006, 83, 323–329. [Google Scholar] [CrossRef]
- Cosentino, S.; Testa, G.; Scordia, D.; Venera, C. Sowing Time and Prediction of Flowering of Different Hemp (Cannabis sativa L.) Genotypes in Southern Europe. Ind. Crops Prod. 2012, 37, 20–33. [Google Scholar] [CrossRef]
- Gill, A.R.; Loveys, B.R.; Cowley, J.M.; Hall, T.; Cavagnaro, T.R.; Burton, R.A. Physiological and morphological responses of industrial hemp (Cannabis sativa L.) to water deficit. Ind. Crops Prod. 2022, 187, 115331. [Google Scholar] [CrossRef]
- Žydelis, R.; Herbst, M.; Weihermüller, L.; Ruzgas, R.; Volungevičius, J.; Barčauskaitė, K.; Tilvikienė, V. Yield potential and factor influencing yield gap in industrial hemp cultivation under nemoral climate conditions. Eur. J. Agron. 2022, 139, 126576. [Google Scholar] [CrossRef]
- Adesina, I.; Bhowmik, A.; Sharma, H.; Shahbazi, A. A Review on the Current State of Knowledge of Growing Conditions, Agronomic Soil Health Practices and Utilities of Hemp in the United States. Agriculture 2020, 10, 129. [Google Scholar] [CrossRef]
- Bocsa, I.; Karus, M.T. The Cultivation of Hemp: Botany, Varieties, Cultivation, and Harvesting; Hemptech: Sebastopo, CA, USA, 1998; ISBN 1886874034. [Google Scholar]
- Herppich, W.B.; Gusovius, H.-J.; Flemming, I.; Drastig, K. Effects of Drought and Heat on Photosynthetic Performance, Water Use and Yield of Two Selected Fiber Hemp Cultivars at a Poor-Soil Site in Brandenburg (Germany). Agronomy 2020, 10, 1361. [Google Scholar] [CrossRef]
- Cosentino, S.; Riggi, E.; Giorgio, T.; Scordia, D.; Venera, C. Evaluation of European Developed Fibre Hemp Genotypes (Cannabis sativa L.) in Semi-Arid Mediterranean Environment. Ind. Crops Prod. 2013, 50, 312–324. [Google Scholar] [CrossRef]
- Di Bari, V.; Campi, P.; Colucci, R.; Mastrorilli, M. Potential Productivity of Fibre Hemp in Southern Europe. Euphytica 2004, 140, 25–32. [Google Scholar] [CrossRef]
- Van der Werf, H.M.G. Agronomy and Crop Physiology of Fibre Hemp: A Literature Review. Center for Agrobiological Research (CABO-DLO) Report, 142. 1991. Available online: https://edepot.wur.nl/346939 (accessed on 18 January 2023).
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key Cultivation Techniques for Hemp in Europe and China. Ind. Crops Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Shiponi, S.; Bernstein, N. The Highs and Lows of P Supply in Medical Cannabis: Effects on Cannabinoids, the Ionome, and Morpho-Physiology. Front. Plant Sci. 2021, 12, 910. [Google Scholar] [CrossRef]
- Cockson, P.; Landis, H.; Smith, T.; Hicks, K.; Whipker, B.E. Characterization of Nutrient Disorders of Cannabis sativa. Appl. Sci. 2019, 9, 4432. [Google Scholar] [CrossRef]
- Maļceva, M.; Vikmane, M.; Stramkale, V. Changes of Photosynthesis-Related Parameters and Productivity of Cannabis sativa Under Different Nitrogen Supply. Environ. Exp. Biol. 2011, 9, 61–69. [Google Scholar]
- Deng, G.; Du, G.; Yang, Y.; Bao, Y.; Liu, F. Planting Density and Fertilization Evidently Influence the Fiber Yield of Hemp (Cannabis sativa L.). Agronomy 2019, 9, 368. [Google Scholar] [CrossRef]
- Landi, S.; Berni, R.; Capasso, G.; Hausman, J.-F.; Guerriero, G.; Esposito, S. Impact of Nitrogen Nutrition on Cannabis sativa: An Update on the Current Knowledge and Future Prospects. Int. J. Mol. Sci. 2019, 20, 5803. [Google Scholar] [CrossRef]
- Tang, K.; Struik, P.; Yin, X.; Calzolari, D.; Musio, S.; Thouminot, C.; Bjelková, M.; Stramkale, V.; Magagnini, G.; Amaducci, S. A Comprehensive Study of Planting Density and Nitrogen Fertilization Effect on Dual-Purpose Hemp (Cannabis sativa L.) Cultivation. Ind. Crops Prod. 2017, 107, 427–438. [Google Scholar] [CrossRef]
- Zhu, R.; Yuan, Q.; Du, G.; Deng, G.; Yang, Y.; Muzammal, R.; Liu, F. Morpho-Physiological Traits, Antioxidant Capacity and Nutrient Accumulation in Hemp (Cannabis sativa L.) Under Varying Levels of Nitrogen Nutrition. J. Plant Nutr. 2021, 45, 854–865. [Google Scholar] [CrossRef]
- Anderson II, S.L.; Pearson, B.; Kjelgren, R.; Brym, Z. Response of Essential Oil Hemp (Cannabis sativa L.) Growth, Biomass, and Cannabinoid Profiles to Varying Fertigation Rates. PLoS ONE 2021, 16, e0252985. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zha, W.; Tang, K.; Deng, G.; Du, G.; Liu, F. Effect of Nitrogen Supply on Growth and Nitrogen Utilization in Hemp (Cannabis sativa L.). Agronomy 2021, 11, 2310. [Google Scholar] [CrossRef]
- Aubin, M.-P.; Seguin, P.; Vanasse, A.; Tremblay, G.F.; Mustafa, A.F.; Charron, J.-B. Industrial Hemp Response to Nitrogen, Phosphorus, and Potassium Fertilization. Crop Forage Turfgrass Manag. 2015, 1, 1–10. [Google Scholar] [CrossRef]
- Finnan, J.; Burke, B. Potassium Fertilization of Hemp (Cannabis sativa). Ind. Crops Prod. 2013, 41, 419–422. [Google Scholar] [CrossRef]
- Yan, L.; Sunoj, V.S.J.; Short, A.W.; Lambers, H.; Elsheery, N.I.; Kajita, T.; Wee, A.K.S.; Cao, K.-F. Correlations Between Allocation to Foliar Phosphorus Fractions and Maintenance of Photosynthetic Integrity in Six Mangrove Populations as Affected by Chilling. New Phytol. 2021, 232, 2267–2282. [Google Scholar] [CrossRef]
- Campbell, B.J.; Berrada, A.F.; Hudalla, C.; Amaducci, S.; McKay, J.K. Genotype × Environment Interactions of Industrial Hemp Cultivars Highlight Diverse Responses to Environmental Factors. Agrosystems Geosci. Environ. 2019, 2, 180057. [Google Scholar] [CrossRef]
- Satriani, A.; Loperte, A.; Pascucci, S. The Cultivation of Industrial Hemp as Alternative Crop in a Less-Favoured Agricultural Area in Southern Italy: The Pignola Case Study. Pollutants 2021, 1, 169–180. [Google Scholar] [CrossRef]
- Sheldon, K.; Shekoofa, A.; Walker, E.; Kelly, H. Physiological Screening for Drought-Tolerance Traits Among Hemp (Cannabis sativa L.) Cultivars in Controlled Environments and in Field. J. Crop Improv. 2021, 35, 816–831. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y. Increasing Inflorescence Dry Weight and Cannabinoid Content in Medical Cannabis Using Controlled Drought Stress. HortScience 2019, 54, 964–969. [Google Scholar] [CrossRef]
- Park, S.-H.; Pauli, C.S.; Gostin, E.L.; Staples, S.K.; Seifried, D.; Kinney, C.; Vanden Heuvel, B.D. Effects of Short-Term Environmental Stresses on the Onset of Cannabinoid Production in Young Immature Flowers of Industrial Hemp (Cannabis sativa L.). J. Cannabis Res. 2022, 4, 1. [Google Scholar] [CrossRef]
- Babaei, M.; Ajdanian, L. Screening of Different Iranian Ecotypes of Cannabis under Water Deficit Stress. Sci. Hortic. 2020, 260, 108904. [Google Scholar] [CrossRef]
- Blandinieres, H.; Leoni, M.; Ferrarini, A.; Amaducci, S. Ranking 26 European Hemp (Cannabis sativa L.) Cultivars for Osmotic Stress Tolerance and Transpiration Efficiency. Ind. Crops Prod. 2021, 170, 113774. [Google Scholar] [CrossRef]
- Bahador, M.; Tadayon, M. Effect of Deficit Irrigation and Zeolite Levels on Phenology, Oil Yield and Water Use Efficiency of Hemp. Iran. J. Field Crop Sci. 2018, 49, 25–38. [Google Scholar] [CrossRef]
- Hu, H.; Liu, H.; Liu, F. Seed Germination of Hemp (Cannabis sativa L.) Cultivars Responds Differently to the Stress of Salt Type and Concentration. Ind. Crops Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Hu, H.; Liu, H.; Du, G.; Yang, F.; Deng, G.; Yang, Y.; Liu, F. Fiber and seed type of hemp (Cannabis sativa L.) responded differently to salt-alkali stress in seedling growth and physiological indices. Ind. Crops Prod. 2019, 129, 624–630. [Google Scholar] [CrossRef]
- Dixit, N. Salinity Induced Antioxidant Defense in Roots of Industrial Hemp (IH: Cannabis sativa L.) for Fiber during Seed Germination. Antioxidants 2022, 11, 244. [Google Scholar] [CrossRef]
- Bolt, M.A.; Beckerman, J.L.; Couture, J.J. Agronomic Management of Industrial Hemp Alters Foliar Traits and Herbivore Performance. Arthropod-Plant Interact. 2021, 15, 137–151. [Google Scholar] [CrossRef]
- Campiglia, E.; Gobbi, L.; Marucci, A.; Rapa, M.; Ruggieri, R.; Vinci, G. Hemp Seed Production: Environmental Impacts of Cannabis sativa L. Agronomic Practices by Life Cycle Assessment (LCA) and Carbon Footprint Methodologies. Sustainability 2020, 12, 6570. [Google Scholar] [CrossRef]
- Van der Werf, H.M.G.; van Geel, W.C.A.; van Gils, L.J.C.; Haverkort, A.J. Nitrogen Fertilization and Row Width Affect Self-Thinning and Productivity of Fibre Hemp (Cannabis sativa L.). Field Crops Res. 1995, 42, 27–37. [Google Scholar] [CrossRef]
- Vera, C.; Malhi, S.; Raney, J.; Wang, Z. The Effect of N and P Fertilization on Growth, Seed Yield and Quality of Industrial Hemp in the Parkland Region of Saskatchewan. Can. J. Plant Sci. 2004, 84, 939–947. [Google Scholar] [CrossRef]
- Vera, C.; Malhi, S.; Phelps, S.; May, W.; Johnson, E. N, P, and S Fertilization Effects on Industrial Hemp in Saskatchewan. Can. J. Plant Sci. 2010, 90, 179–184. [Google Scholar] [CrossRef]
- Barčauskaitė, K.; Barčauskaitė, A.; Szummy, A.; Tilvikienė, V. Variation of secondary metabolites in Cannabis sativa L. inflorescences under applied agrotechnological measures. Ind. Crops Prod. 2022, 188, 115570. [Google Scholar] [CrossRef]
- Amaducci, S.; Zatta, A.; Pelatti, F.; Venturi, G. Influence of Agronomic Factors on Yield and Quality of Hemp (Cannabis sativa L.) Fibre and Implication for an Innovative Production System. Field Crops Res. 2008, 107, 161–169. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Hernández, A.; Ferreiro-Vera, C.; Zuazo, V.H.D.; García, J.H.; Sánchez-Carnerero, C.; Casano, S. Yield of New Hemp Varieties for Medical Purposes under Semi-Arid Mediterranean Environment Conditions. Comun. Sci. 2020, 11, e3264. [Google Scholar] [CrossRef]
- Tsaliki, E.; Kalivas, A.; Jankauskiene, Z.; Irakli, M.; Cook, C.; Grigoriadis, I.; Panoras, I.; Vasilakoglou, I.; Dhima, K. Fibre and Seed Productivity of Industrial Hemp (Cannabis sativa L.) Varieties under Mediterranean Conditions. Agronomy 2021, 11, 171. [Google Scholar] [CrossRef]
- Cherrett, N.; Barrett, J.; Clemett, A.; Chadwick, M.; Chadwick, M.J. Ecological Footprint and Water Analysis of Cotton, Hemp and Polyester; Stockholm Environment Institute: Stockholm, Sweden, 2005. [Google Scholar]
- Calzolari, D.; Magagnini, G.; Lucini, L.; Grassi, G.; Appendino, G.B.; Amaducci, S. High Added-Value Compounds from Cannabis Threshing Residues. Ind. Crops Prod. 2017, 108, 558–563. [Google Scholar] [CrossRef]
- Hoppner, F.; Menge-Hartmann, U. Yield and Quality of Fibre and Oil of Fourteen Hemp Cultivars in Northern Germany at Two Harvest Dates. Landbauforsch. Völkenrode 2007, 3, 219–232. [Google Scholar]
- Vodolazska, D.; Lauridsen, C. Effects of Dietary Hemp Seed Oil to Sows on Fatty Acid Profiles, Nutritional and Immune Status of Piglets. J. Anim. Sci. Biotechnol. 2020, 11, 1–18. [Google Scholar]
- Abdollahi, M.; Sefidkon, F.; Calagari, M.; Mousavi, A.; Mahomoodally, M.F. A Comparative Study of Seed Yield and Oil Composition of Four Cultivars of Hemp (Cannabis sativa L.) Grown from Three Regions in Northern Iran. Ind. Crops Prod. 2020, 152, 112397. [Google Scholar] [CrossRef]
- Abdollahi, M.; Sefidkon, F.; Calagari, M.; Mousavi, A.; Mahomoodally, M.F. Impact of Four Hemp (Cannabis sativa L.) Varieties and Stage of Plant Growth on Yield and Composition of Essential Oils. Ind. Crops Prod. 2020, 155, 112793. [Google Scholar] [CrossRef]
- Sanchez, L.; Filter, C.; Baltensperger, D.; Kurouski, D. Confirmatory Non-Invasive and Non-Destructive Differentiation Between Hemp and Cannabis Using a Hand-Held Raman Spectrometer. RSC Adv. 2020, 10, 3212–3216. [Google Scholar] [CrossRef]
- Smith, B.C.; Lessard, P.; Pearson, R. Inter-Laboratory Variation in Cannabis Analysis: Pesticides and Potency in Distillates. Cannabis Sci. Technol. 2019, 2, 1–66. [Google Scholar]
- Reeves, R.D.; Baker, A.J.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A Global Database for Plants that Hyperaccumulate Metal and Metalloid Trace Elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef]
- Gong, Y.; Zhao, D.; Wang, Q. An Overview of Field-Scale Studies on Remediation of Soil Contaminated with Heavy Metals and Metalloids: Technical Progress over the Last Decade. Water Res. 2018, 147, 440–460. [Google Scholar] [CrossRef]
- EPA (Environmental Protection Agency). Phytoremediation Resource Guide. EPA-542-B-990-003.1999.
- Kozminska, A.; Wiszniewska, A.; Hanus-Fajerska, E.; Muszyńska, E. Recent Strategies of Increasing Metal Tolerance and Phytoremediation Potential Using Genetic Transformation of Plants. Plant Biotechnol. Rep. 2018, 12, 1–14. [Google Scholar] [CrossRef]
- Charkowski, E. Hemp ‘Eats’ Chernobyl Waste. Central Oregon Green Pages. 1998. Available online: www.empirenet.net/empnet/centrorg.htm (accessed on 18 January 2023).
- Citterio, S.; Santagostino, A.; Fumagalli, P.; Prato, N.; Ranalli, P.; Sgorbati, S. Heavy Metal Tolerance and Accumulation of Cd, Cr and Ni by Cannabis sativa L. Plant Soil 2003, 256, 243–252. [Google Scholar] [CrossRef]
- Husain, R.; Weeden, H.; Bogush, D.; Deguchi, M.; Soliman, M.; Potlakayala, S.; Katam, R.; Goldman, S.; Rudrabhatla, S. Enhanced Tolerance of Industrial Hemp (Cannabis sativa L.) Plants on Abandoned Mine Land Soil Leads to Overexpression of Cannabinoids. PLoS ONE 2019, 14, e0221570. [Google Scholar] [CrossRef]
- Deng, G.; Yang, M.; Saleem, M.H.; Rehman, M.; Fahad, S.; Yang, Y.; Elshikh, M.S.; Alkahtani, J.; Ali, S.; Khan, S.M. Nitrogen Fertilizer Ameliorate the Remedial Capacity of Industrial Hemp (Cannabis sativa L.) Grown in Lead Contaminated Soil. J. Plant Nutr. 2021, 44, 1770–1778. [Google Scholar] [CrossRef]
- Xu, Y.; Deng, G.; Guo, H.; Yang, M.; Yang, Q. Accumulation and sub cellular distribution of lead (Pb) in industrial hemp grown in Pb contaminated soil. Ind. Crops Prod. 2021, 161, 113220. [Google Scholar] [CrossRef]
- Linger, P.; Müssig, J.; Fischer, H.; Kobert, J. Industrial Hemp (Cannabis sativa L.) Growing on Heavy Metal Contaminated Soil: Fibre Quality and Phytoremediation Potential. Ind. Crops Prod. 2002, 16, 33–42. [Google Scholar] [CrossRef]
- Linger, P.; Ostwald, A.; Haensler, J. Cannabis sativa L. Growing on Heavy Metal Contaminated Soil: Growth, Cadmium Uptake and Photosynthesis. Biol. Plant. 2005, 49, 567–576. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q. Cadmium Tolerance and Accumulation in Eight Potential Energy Crops. Biotechnol. Adv. 2009, 27, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Cai, Q.; Liu, Q.; Wu, L. Salicylic Acid-Mediated Alleviation of Cadmium Toxicity in Hemp Plants in Relation to Cadmium Uptake, Photosynthesis, and Antioxidant Enzymes. Acta Physiol. Plant. 2009, 31, 969–977. [Google Scholar] [CrossRef]
- Campbell, S.; Paquin, D.; Awaya, J.D.; Li, Q.X. Remediation of Benzo[a]pyrene and Chrysene-Contaminated Soil with Industrial Hemp (Cannabis sativa). Int. J. Phytoremediation 2002, 4, 157–168. [Google Scholar] [CrossRef]
- Vandenhove, H.; Van Hees, M. Fibre Crops as Alternative Land Use for Radioactively Contaminated Arable Land. J. Environ. Radioact. 2005, 81, 131–141. [Google Scholar] [CrossRef]
- Grist.org. 2022. Available online: https://grist.org/science/pfas-is-contaminating-farms-can-hemp-help/ (accessed on 18 January 2023).
- Gunther, F. Carbon Sequestration for Everybody: Decrease Atmospheric Carbon Dioxide, Earn Money and Improve the Soil. 2007. Available online: https://terrapreta.bioenergylists.org/files/Terra%20pretav1_0.pdf (accessed on 18 January 2023).
- Jansson, C.; Wullschleger, S.D.; Kalluri, U.C.; Tuskan, G.A. Phytosequestration: Carbon Biosequestration by Plants and the Prospects of Genetic Engineering. BioScience 2010, 60, 685–696. [Google Scholar] [CrossRef]
- Kumar, V.G.; Ramadoss, R.; Rampradheep, G.S. A Study Report on Carbon Sequestration by Using Hempcrete. Mater. Today Proc. 2021, 45, 6369–6371. [Google Scholar] [CrossRef]
- Arehart, J.H.; Nelson, W.S.; Srubar III, W.V. On the Theoretical Carbon Storage and Carbon Sequestration Potential of Hempcrete. J. Clean. Prod. 2002, 266, 121846. [Google Scholar] [CrossRef]
- Jami, T.; Kumar, S. Assessment of Carbon Sequestration of Hemp Concrete. In International Conference on Advances in Construction Materials and Systems; RILEM: Chennai, India, 2017; pp. 665–673. [Google Scholar]
- Jami, T.; Karade, S.R.; Singh, L.P. A Review of the Properties of Hemp Concrete for Green Building Applications. J. Clean. Prod. 2019, 239, 117852. [Google Scholar] [CrossRef]
- Jami, T.; Rawtani, D.; Agriwal, Y.K. Hemp Concrete: Carbon-Negative Construction. Emerg. Mater. Res. 2016, 5, 240–247. [Google Scholar] [CrossRef]
- Kraszkiewicz, A.; Kachel, M.; Parafiniuk, S.; Zając, G.; Niedziółka, I.; Sprawka, M. Assessment of the Possibility of Using Hemp Biomass (Cannabis sativa L.) for Energy Purposes: A Case Study. Appl. Sci. 2019, 9, 4437. [Google Scholar] [CrossRef]
- Prade, T.; Svensson, S.-E.; Mattsson, J.E. Energy Balances for Biogas and Solid Biofuel Production from Industrial Hemp. Biomass Bioenergy 2012, 40, 36–52. [Google Scholar] [CrossRef]
- Kreuger, E.; Prade, T.; Escobar, F.; Svensson, S.E.; Englund, J.E.; Björnsson, L. Anaerobic Digestion of Industrial Hemp–Effect of Harvest Time on Methane Energy Yield Per Hectare. Biomass Bioenergy 2011, 35, 893–900. [Google Scholar] [CrossRef]
- Adamovics, A.; Dubrovskis, V.; Platace, R. Productivity of Industrial Hemp and its Utilization for Anaerobic Digestion. WIT Trans. Ecol. Environ. 2014, 190, 1045–1055. [Google Scholar] [CrossRef]
- Li, S.Y.; Stuart, J.D.; Li, Y.; Parnas, R.S. The Feasibility of Converting Cannabis sativa L. Oil into Biodiesel. Bioresour. Technol. 2010, 101, 8457–8460. [Google Scholar] [CrossRef]
- Rashid, U.; Bhatti, S.; Ansari, T.; Yunus, R.; Ibrahim, M. Biodiesel Production from Cannabis sativa Oil from Pakistan. Energy Sources Part A Recovery Util. Environ. Eff. 2016, 38, 865–875. [Google Scholar] [CrossRef]
- Salentijn, E.M.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New Developments in Fiber Hemp (Cannabis sativa L.) Breeding. Ind. Crops Prod. 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Cornell Hemp. 2022. Available online: https://hemp.cals.cornell.edu/ (accessed on 18 January 2023).
- AgriLife Today. Texas A&M AgriLife, Partners Achieve Stable Transformation in Industrial Hemp. 2022. Available online: https://agrilifetoday.tamu.edu/2022/01/03/texas-am-agrilife-partners-achieve-stable-transformation-in-industrial-hemp/?fbclid=IwAR0uXtj_yV9FEOm6Gc1u8P-y5WlRDtXTW6AWvY2Cq0vBUPtRbqR3pnfAt-s&fs=e&s=cl (accessed on 18 January 2023).
- Texas Row Crops Newsletter. Industrial Hemp Update. 2022. Available online: https://agrilife.org/texasrowcrops/2022/07/01/industrial-hemp-update/ (accessed on 18 January 2023).
- Meijer, W.J.M.; Van der Werf, H.M.G.; Mathijssen, E.W.J.M.; Van den Brink, P.W.M. Constraints to Dry Matter Production in Fiber Hemp (Cannabis sativa L.). Eur. J. Agron. 1995, 4, 109–117. [Google Scholar] [CrossRef]
- Hanks, A. Canadian Hemp Update. J. Ind. Hemp 2007, 13, 49–57. [Google Scholar] [CrossRef]
- Ranalli, P. Current Status and Future Scenarios of Hemp Breeding. Euphytica 2004, 140, 121–131. [Google Scholar] [CrossRef]
- Ellison, S. Hemp (Cannabis sativa L.) Research Priorities: Opinions from United States Hemp Stakeholders. GCB Bioenergy 2021, 13, 562–569. [Google Scholar] [CrossRef]
- Watanabe, K.; Itokawa, Y.; Yamaori, S.; Funahashi, T.; Kimura, T.; Kaji, T.; Usami, N.; Yamamoto, I. Conversion of Cannabidiol to Δ9-Tetrahydrocannabinol and Related Cannabinoids in Artificial Gastric Juice, and Their Pharmacological Effects in Mice. Forensic Toxicol. 2007, 25, 16–21. [Google Scholar] [CrossRef]
- Golombek, P.; Müller, M.; Barthlott, I.; Sproll, C.; Lachenmeier, D.W. Conversion of Cannabidiol (CBD) into Psychotropic Cannabinoids Including Tetrahydrocannabinol (THC): A Controversy in the Scientific Literature. Toxics 2020, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Roulac, J.W. Hemp Horizons: The Comeback of the World’s Most Promising Plant; Chelsea Green Publishing Company: Chelsea, VT, USA, 1997. [Google Scholar]
- Yan, W.; Wallace, D.H. Simulation and prediction of plant phenology for five crops based on photoperiod temperature interaction. Ann. Bot. 1998, 81, 705–716. [Google Scholar] [CrossRef]
- Van der Werf, H.; Mathussen, E.; Haverkort, A. The Potential of Hemp (Cannabis sativa L.) for Sustainable Fibre Production: A Crop Physiological Appraisal. Ann. Appl. Biol. 1996, 129, 109–123. [Google Scholar] [CrossRef]
- Tang, K.; Struik, P.C.; Amaducci, S.; Stomph, T.-J.; Yin, X. Hemp (Cannabis sativa L.) Leaf Photosynthesis in Relation to Nitrogen Content and Temperature: Implications for Hemp as a Bio-economically Sustainable Crop. GCB Bioenergy 2017, 9, 1573–1587. [Google Scholar] [CrossRef]
- Douglas, R. Invasion of the Drones: How Unmanned Aerial Vehicles Can Help Cannabis Growers Improve Crop Quality, Lower Costs, and Save the Planet. Cannabis Sci. Technol. 2020, 3, 50–54. [Google Scholar]
- Tang, K.; Wang, J.; Yang, Y.; Deng, G.; Yu, J.; Hu, W.; Guo, L.; Du, G.; Liu, F. Fiber Hemp (Cannabis sativa L.) Yield and Its Response to Fertilization and Planting Density in China. Ind. Crops Prod. 2022, 177, 114542. [Google Scholar] [CrossRef]
- Popper, K.R. The Logic of Scientific Discovery; Basic Books: New York, NY, USA, 1959; p. 479. [Google Scholar]
- Platt, J.R. Strong Inference. Science 1964, 146, 347–353. [Google Scholar] [CrossRef]
- Lyu, D.; Backer, R.; Robinson, W.G.; Smith, D.L. Plant Growth-Promoting Rhizobacteria for Cannabis Production: Yield, Cannabinoid Profile and Disease Resistance. Front. Microbiol. 2019, 10, 1761. [Google Scholar] [CrossRef] [PubMed]
- Citterio, S.; Prato, N.; Fumagalli, P.; Aina, R.; Massa, N.; Santagostino, A.; Sgorbati, S.; Berta, G. The Arbuscular Mycorrhizal Fungus Glomus mosseae Induces Growth and Metal Accumulation Changes in Cannabis sativa L. Chemosphere 2005, 59, 21–29. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M. Cannabis Pests. J. Int. Hemp Assoc. 1996, 3, 52–55. [Google Scholar]
- Young, E.M. Revival of Industrial Hemp: A Systematic Analysis of the Current Global Industry to Determine Limitations and Identify Future Potentials Within the Concept of Sustainability. Master’s Thesis, Lund University, Lund, Sweden, 2005. [Google Scholar]
- Ali, E.; Almagboul, A.; Khogali, S.; Gergeir, U. Antimicrobial Activity of Cannabis sativa L. Chin. Med. 2012, 3, 61–64. [Google Scholar] [CrossRef]
- Park, S.-H.; Staples, S.K.; Gostin, E.L.; Smith, J.P.; Vigil, J.J.; Seifried, D.; Kinney, C.; Pauli, C.S.; Heuvel, B.D.V. Contrasting Roles of Cannabidiol as an Insecticide and Rescuing Agent for Ethanol–Induced Death in the Tobacco Hornworm Manduca sexta. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- McPartland, J.M. Cannabis as Repellent and Pesticide. J. Int. Hemp Assoc. 1997, 4, 87–92. [Google Scholar]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd-Allah, E.F.; Bae, H. Genome Editing Tools in Plants. Genes 2017, 8, 399. [Google Scholar] [CrossRef]
- Dolgin, E. A Boosted Crop. Nature 2019, 572, S5–S7. Available online: https://media.nature.com/original/magazine-assets/d41586-019-02525-4/d41586-019-02525-4.pdf (accessed on 18 January 2023). [CrossRef]
- Cabezudo, B.; Recio, M.; Sánchez-Laulhé, J.; Trigo, M.D.M.; Toro, F.J.; Polvorinos, F. Atmospheric Transportation of Marihuana Pollen from North Africa to the Southwest of Europe. Atmos. Environ. 1997, 31, 3323–3328. [Google Scholar] [CrossRef]
- Carinanos, P.; Galán, C.; Alcázar, P.; Domínguez, E. Analysis of the Particles Transported with Dust-Clouds Reaching Cordoba, Southwestern Spain. Arch. Environ. Contam. Toxicol. 2004, 46, 141–146. [Google Scholar] [CrossRef]
- Welling, M.T.; Shapter, T.; Rose, T.J.; Liu, L.; Stanger, R.; King, G.J. A Belated Green Revolution for Cannabis: Virtual Genetic Resources to Fast-Track Cultivar Development. Front. Plant Sci. 2016, 7, 1113. [Google Scholar] [CrossRef]
- Pate, D.W. Possible Role of Ultraviolet Radiation in Evolution of Cannabis Chemotypes. Econ. Bot. 1983, 37, 396–405. [Google Scholar] [CrossRef]
- Pate, D.W. Chemical Ecology of Cannabis. J. Int. Hemp Assoc. 1994, 2, 32–37. [Google Scholar]
- Gülck, T.; Møller, B.L. Phytocannabinoids: Origins and Biosynthesis. Trends Plant Sci. 2020, 10, 985–1004. [Google Scholar] [CrossRef]
- Mansouri, H.; Asrar, Z.; Mehrabani, M. Effects of Gibberellic Acid on Primary Terpenoids and Delta-Tetrahydrocannabinol in Cannabis Sativa at Flowering Stage. J. Integr. Plant Biol. 2009, 51, 553–561. [Google Scholar] [CrossRef]
- McPartland, J.M.; Russo, E.B. Cannabis and Cannabis Extracts: Greater than the Sum of Their Parts? J. Cannabis Ther. 2001, 1, 103–132. [Google Scholar] [CrossRef]
- Izzo, A.A.; Borrelli, F.; Capasso, R.; Di Marzo, V.; Mechoulam, R. Non-Psychotropic Plant Cannabinoids: New Therapeutic Opportunities from an Ancient Herb. Trends Pharmacol. Sci. 2009, 30, 515–527. [Google Scholar] [CrossRef]
- Deguchi, M.; Bogush, D.; Weeden, H.; Spuhler, Z.; Potlakayala, S.; Kondo, T.; Zhang, Z.J.; Rudrabhatla, S. Establishment and Optimization of a Hemp (Cannabis sativa L.) Agroinfiltration System for Gene Expression and Silencing Studies. Sci. Rep. 2020, 10, 3504. [Google Scholar] [CrossRef]
- Sunoj, V.J.; Shroyer, K.J.; Jagadish, S.K.; Prasad, P.V. Diurnal Temperature Amplitude Alters Physiological and Growth Response of Maize (Zea mays L.) During the Vegetative Stage. Environ. Exp. Bot. 2016, 130, 113–121. [Google Scholar] [CrossRef]
- Sunoj, V.J.; Prasad, P.V.; Ciampitti, I.A.; Maswada, H.F. Narrowing Diurnal Temperature Amplitude Alters Carbon Tradeoff and Reduces Growth in C4 Crop Sorghum. Front. Plant Sci. 2020, 11, 1262. [Google Scholar] [CrossRef]
- Impa, S.M.; Sunoj, V.S.J.; Krassovskaya, I.; Bheemanahalli, R.; Obata, T.; Jagadish, S.V.K. Carbon Balance and Source-Sink Metabolic Changes in Winter Wheat Exposed to High Night-Time Temperature. Plant Cell Environ. 2019, 42, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Di Candilo, M.; Ranalli, P.; Dal Re, L. Heavy Metal Tolerance and Uptake of Cd, Pb and Tl by Hemp. Adv. Hortic. Sci. 2004, 18, 138–144. [Google Scholar]
- Gao, W.; Schlüter, S.; Blaser, S.R.G.A.; Shen, J.; Vetterlein, D. A Shape-Based Method for Automatic and Rapid Segmentation of Roots in Soil from X-Ray Computed Tomography Images: Rootine. Plant Soil 2019, 441, 643–655. [Google Scholar] [CrossRef]
- Zhang, X.; Derival, M.; Albrecht, U.; Ampatzidis, Y. Evaluation of a Ground Penetrating Radar to Map the Root Architecture of HLB-Infected Citrus Trees. Agronomy 2019, 9, 354. [Google Scholar] [CrossRef]
- Lisson, S.N.; Mendham, N.J.; Carberry, P.S. Development of a hemp (Cannabis sativa L.) Simulation Model: 4. Model Description and Validation. Aust. J. Exp. Agric. 2000, 40, 425–432. [Google Scholar] [CrossRef]
- Tang, K.; Fracasso, A.; Struik, P.C.; Yin, X.; Amaducci, S. Water-and Nitrogen-Use Efficiencies of Hemp (Cannabis sativa L.) Based on Whole-Canopy Measurements and Modeling. Front. Plant Sci. 2018, 9, 951. [Google Scholar] [CrossRef]
- Ramanathan, R. An Introduction to Data Envelopment Analysis: A Tool for Performance Measurement; Sage Publications: New Delhi, India, 2003. [Google Scholar]
- Maurer, J.L.; Griffin, T.W.; Sharda, A. Site-Specific Scale Efficiency Determined by Data Envelopment Analysis of Precision Agriculture Field Data. In Proceedings of the 13th International Conference on Precision Agriculture, St. Louis, MI, USA, 31 July–4 August 2016. [Google Scholar]
- Ceyhan, V.; Türkten, H.; Yıldırım, Ç.; Canan, S. Economic Viability of Industrial Hemp Production in Turkey. Ind. Crops Prod. 2022, 176, 114354. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sunoj Valiaparambil Sebastian, J.; Dong, X.; Trostle, C.; Pham, H.; Joshi, M.V.; Jessup, R.W.; Burow, M.D.; Provin, T.L. Hemp Agronomy: Current Advances, Questions, Challenges, and Opportunities. Agronomy 2023, 13, 475. https://doi.org/10.3390/agronomy13020475
Sunoj Valiaparambil Sebastian J, Dong X, Trostle C, Pham H, Joshi MV, Jessup RW, Burow MD, Provin TL. Hemp Agronomy: Current Advances, Questions, Challenges, and Opportunities. Agronomy. 2023; 13(2):475. https://doi.org/10.3390/agronomy13020475
Chicago/Turabian StyleSunoj Valiaparambil Sebastian, John, Xuejun Dong, Calvin Trostle, Hanh Pham, Madhumita V. Joshi, Russell W. Jessup, Mark D. Burow, and Tony L. Provin. 2023. "Hemp Agronomy: Current Advances, Questions, Challenges, and Opportunities" Agronomy 13, no. 2: 475. https://doi.org/10.3390/agronomy13020475
APA StyleSunoj Valiaparambil Sebastian, J., Dong, X., Trostle, C., Pham, H., Joshi, M. V., Jessup, R. W., Burow, M. D., & Provin, T. L. (2023). Hemp Agronomy: Current Advances, Questions, Challenges, and Opportunities. Agronomy, 13(2), 475. https://doi.org/10.3390/agronomy13020475